
Transcriptome-guided discovery and functional characterization of two UDP-sugar 4-epimerase families involved in the biosynthesis of anti-tumor polysaccharides in Ornithogalum caudatum†
Abstract
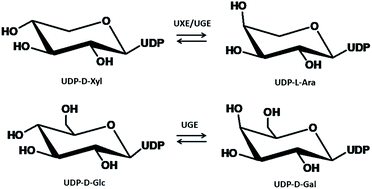
UDP-glucose 4-epimerase (UGE) and UDP-xylose 4-epimerase (UXE), two important UDP-sugar 4-epimerases, are well known to be essential for de novo biosynthesis of UDP-D-galactose and UDP-L-arabinose, two universal sugar donors for the formation of four galactose- and arabinose-containing anticancer polysaccharides in Ornithogalum caudatum. However, very little is known about their cDNA sequences. Furthermore, the functional significance of the two epimerases in the biosynthesis of these anticancer polysaccharides in O. caudatum had not been documented. Here, we presented a full characterization of UGE and UXE, which were deemed to be responsible for anticancer polysaccharides biosynthesis in O. caudatum. Specifically, a transcriptome-guided search for the two epimerase genes in O. caudatum was first performed in the present study. A total of 4 unigenes sharing high sequence identity with UDP-sugar 4-epimerases were retrieved from transcriptome assembly. Four full-length cDNAs encoding UDP-sugar 4-epimerases, including two UGE-like and two UXE-like genes, were then isolated by reverse transcription polymerase chain reaction (RT-PCR) from O. caudatum. Bio-informatic analysis indicated the two UDP-sugar 4-epimerase families shared two common conserved domains, namely an N-terminal GxxGxxG motif and a catalytic Ser/Thr-Tyr-Lys triad. A phylogenetic analysis revealed the two members in the same UGE family could be classified into two subgroups, revealing their divergently functional significance. These candidate isoenzymes were screened by functional expression in E. coli individually as standalone enzymes. Two UGE-like cDNAs were identified to be bona fide genes, exhibiting both UGE and UXE activities. To further explore the possible role of these epimerase proteins in polysaccharides biosynthesis, transcript profiles of the four genes were subsequently examined by real-time quantitative PCR in various O. caudatum tissues. OcUGE1, OcUGE2 and OcUXE1 were therefore assumed to be responsible for the biosynthesis of the four galactose- and arabinose-containing polysaccharides due to their expression profiles in O. caudatum. Taken together, these data provide further comprehensive knowledge for polysaccharides biosynthesis in O. caudatum and broaden the potential application of UGE in metabolic engineering or synthetic biology as a potential gene part.
 Please wait while we load your content...
Please wait while we load your content...

