
Curcumin induces cell death of human papillary thyroid carcinoma BCPAP cells through endoplasmic reticulum stress
Abstract
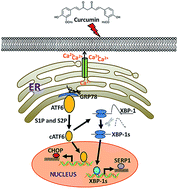
Curcumin, a major active component of Curcuma longa, is a natural polyphenolic antioxidant compound which has a strong potential for cancer prevention and treatment. However, the effect of curcumin on papillary thyroid carcinoma cells has not been investigated. In the present study, we report that curcumin induces cell death of papillary thyroid carcinoma (PTC) cell line BCPAP cells through endoplasmic reticulum (ER) stress. Curcumin suppressed cell viability of BCPAP cells in a dose-dependent manner. A large body of evidence demonstrates that ER stress contributes to cell death in certain contexts. ER stress was induced by curcumin treatment in BCPAP cells, as evidenced by the up-regulation of intracellular calcium level, and up-regulated expression of CCAAT/enhancer binding protein homologous protein (CHOP) at both mRNA and protein levels compared to the control group. Moreover, the activation of activating transcription factor 6 (ATF6)/X-box binding protein-1 (XBP-1) pathways could be involved in the ER stress signaling of curcumin treatment. Thus, our results provide new insights into the molecular mechanisms of curcumin-mediated cell death in PTC cells and suggest curcumin might be a potential chemotherapeutic agent which is able to fight against papillary thyroid cancer.
 Please wait while we load your content...
Please wait while we load your content...

