DOI:
10.1039/C5RA28171G
(Paper)
RSC Adv., 2016,
6, 27659-27668
Effect of sequential hydrolysis with endo- and exo-peptidase on bitterness properties of wheat gluten hydrolysates
Received
31st December 2015
, Accepted 1st March 2016
First published on 2nd March 2016
Abstract
After the bitterness of wheat gluten (WG) hydrolysates using single-enzyme treatment was assessed by taste dilution analysis (TDA), schemes of sequential hydrolysis with endo- and exo- peptidases were designed to prepare low-bitterness small-peptide powders. This study is the first to attempt the enrichment of the low-bitterness small peptide and search for explanations for the disparity of the bitterness between the schemes. The effects of the peptide nitrogen content, amino acid composition, molecular weight distribution and amino acid sequence on the sensory characterization and sensor response of the electronic tongue in WG hydrolysates are discussed. The results showed the disparity of the bitterness between all schemes was determined by the bitter taste thresholds of bitter peptides, but not the content of bitter peptides. Bitter taste thresholds were affected by the amino acid sequences and molecular weights of the bitter peptides. It was found that when WG was hydrolysed for 300 min by Proteax, the hydrolysates exhibited the lowest bitterness while they had the highest content of small peptides ranging from 180–500 Da.
Introduction
The water-insoluble characteristics of wheat gluten (WG), which has a high proportion of hydrophobic amino acids, strongly limit its application in the food industry. Enzymatic hydrolysis of WG to produce value-added peptides is very attractive.1,2 The resulting bioactive peptides comprise a great variety of peptides, with small peptides (2–6 amino acids) representing by far the largest category.3 It has been reported that small peptides were more rapidly and more evenly absorbed through the small intestine than a free amino acid mixture of the same amino acid composition,4 and had higher nutritive value.5,6 However, the content of small peptides in the protein hydrolysates was low and the enzyme-hydrolyzed protein had strong bitterness, which made them difficult to incorporate into foods. It is important to control the hydrolysis process to obtain an ideal debittering effect and high yield of small peptides, especially dipeptides and tripeptides which could be easily absorbed by the body.
Previous research showed that the main bitter substances of enzyme-catalyzed protein hydrolysates were bitter peptides of MWs below 6 kDa 7 but not free amino acids.8 The peptides that had a higher ratio of hydrophobic amino acids tended to have a smaller bitter taste threshold.9 For example, the bitterness of di-, tri- and tetra-leucines was 8, 15 and 30 times more intense than mono-leucine (Leu), respectively. The polyleucines were more bitter than expected from the number of Leu residues present in the Leu oligomers concerned.10 The same phenomenon extended to phenylalanine (Phe) and tyrosine (Tyr).11 Interestingly, the positions of hydrophobic amino acids in the peptide chain greatly impacted the bitter taste threshold of the peptide. Fitzgerald12 and Cho13 suggested that the peptide was more bitter when the hydrophobic amino acids were located in the endo position of a long-chain peptide than when located in the terminal position. However, for short peptides prepared at a high degree of hydrolysis (DH), the situation is the opposite. Therefore, it is necessary to investigate the effect of the bitter taste thresholds of bitter peptides on the bitterness of WG hydrolysates.
Proteax is a proteolytic enzyme preparation manufactured by a fermenting process with a selected strain of Aspergillus oryzae. Proteax yielded high endo- and exo-activities of proteins in contrast to other fungal proteinase preparations. For Proteax treatment of WG, peptide bonds were preferentially cleaved at the carboxyl-terminal of glutamine (Gln), serine (Ser), threonine (Thr) and methionine (Met) followed by cleavage on the carboxyl-terminal of Arg and finally on the carboxyl-terminal of Lys, alanine (Ala), Phe and Leu. It was concluded that Proteax was suitable for preparing low-bitterness small peptide powders.
Sequential hydrolysis with endo- and exo-peptidase should be more efficient to obtain small peptides, since the combination has a greater hydrolysis ability relative to the single endopeptidase or exopeptidase. The aim of the current research was to determine the effect of sequential hydrolysis with endo- and exo-peptidase on the bitterness properties of the resulting hydrolysates. The intention was to reduce the bitterness intensity of the peptide powder as much as possible. According to the enzymatic hydrolysis characteristics of a single enzyme, six types of experimental schemes were designed for a comparative study. The optimal scheme was proposed based on the bitterness of the WG hydrolysates and the yield of small peptides.
Experimental
Materials and reagents
WG (75% protein, wet base) was provided by Nan Tong Lian Hai Wei Jing Co., Ltd. (Jiangsu, China). Proteax (1400 U g−1, combined proteases) was donated by Amano Enzyme Inc (Nagoya, Japan). Alcalase (2.4L, 2.4 AU g−1, alkaline protease), Flavourzyme (500 LAPU g−1, exopeptidase), Neutrase 1.5MG (1.5 AU g−1; neutral protease) and Protamex (1.5 AU g−1; combined proteases) were kindly supplied by Novo Nordisk (Bagsvaerd, Denmark). Papain (6000 U mg−1, an alkaline protease from papaya latex) and Trypsin (50![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 000 U g−1, an alkaline protease from porcine pancreas) were purchased from Sinopharm Chemical Reagent Co., Ltd. (Shanghai, China). All other reagents and chemicals were of analytical grade.
000 U g−1, an alkaline protease from porcine pancreas) were purchased from Sinopharm Chemical Reagent Co., Ltd. (Shanghai, China). All other reagents and chemicals were of analytical grade.
Enzymatic hydrolysis of wheat gluten
WG was hydrolyzed with seven different enzymes, under the detailed conditions described in Table 1. 10 g of WG were dissolved in 200 mL of deionised water at room temperature. The suspension was adjusted to the appropriate pH, temperature and enzyme-to-substrate (E/S) ratio, depending on the respective enzyme used, and incubated for 1 h with continuous stirring. The reaction was continued for 12 h and the pH of the solution was kept constant by continuously adding 1.0 mol L−1 NaOH or HCl solution. The proteolysis reaction was stopped at intervals of 15, 30, 60, 120, 180, 300, 360, 480, and 720 min, with the enzyme immediately inactivated by heat treatment in boiling water for 15 min. Then the WG hydrolysates were centrifuged at 10000 × g for 15 min at 4 °C in a ZOPR-52D refrigerated centrifuge (Hitachi Koki Co. Ltd., Tokyo, Japan). The supernatants were lyophilized at −20 °C before further analysis.
Table 1 Conditions for the hydrolysis of wheat gluten by different proteases
| Hydrolysis condition |
Proteases |
| Proteax |
Alcalase |
Protamex |
Neutrase |
Flavourzyme |
Trypsin |
Papain |
| pH |
7 |
8 |
6.5 |
7 |
7 |
7 |
6.5 |
| T (°C) |
50 |
50 |
50 |
50 |
50 |
37 |
55 |
| E/S ratio (w/w) |
1:50 |
1:20 |
1:100 |
1:100 |
1:20 |
1:20 |
1:20 |
Determination of the degree of hydrolysis
The degree of hydrolysis (DH) during the enzymatic hydrolysis reaction was measured by the pH-stat method.14 DH was calculated based on the amount of NaOH consumed and defined as follows:| |
 | (1) |
where: B was the base (NaOH) consumed in mL; Nb was the normality of the base (NaOH); α was the average degree of dissociation of the α-NH2 groups; Mp was the mass of the substrate (protein, determined as N × 5.7) in the reaction; htot was total number of peptide bonds in the protein substrate (8.38 meq. per g protein for WG).
Taste dilution analysis
The bitterness of WG hydrolysates at different times (0–12 h) of enzymatic hydrolysis or at a certain degree of hydrolysis (5%, 9% or 15%) was evaluated by the TDA method,15 with some minor changes. Briefly, 1 g of freeze-dried enzymatic hydrolysates was dispersed in 100 mL of deionised water at room temperature. WG hydrolysates were diluted stepwise at a 1:1 ratio with deionized water, and were then presented to a trained sensory panel (5 males and 5 females, aged between 20 and 35 years) in the order of increasing solute concentrations, and judged in a triangle test. The taste dilution (TD) factor was defined as the dilution at which a taste difference between the diluted sample and two blanks (1% WG solution) could just be detected. The TD value was the average value of the evaluation results provided by the inspectors. The differences among the results should not be more than one of the dilution levels.
Preparation of powdered small peptides with low-bitterness
The preparation of extensively hydrolysed WG by five different enzymes (Proteax, Alcalase, Trypsin, Neutrase and Flavourzyme) according to the conditions described in Table 1 is illustrated in Fig. 1. On the basis of the DH and the corresponding TD factors of seven different enzymes, six types of experimental schemes were designed for preparing a low-bitterness small-peptide powder. WG (5% w/v) was hydrated for 1 h in a shaking water bath at room temperature. In the first three schemes, the WG suspensions were hydrolysed for 2, 5 or 8 h (abbreviated respectively as Pro-l, Pro-m and Pro-h) by Proteax alone. For the following three schemes, the WG suspensions were also hydrolysed for 2, 1 and 6 h (abbreviated respectively as Alc-2, Try-1, and Neu-6) by Alcalase, Trypsin and Neutrase respectively. Afterwards, Alc-2, Try-1, and Neu-6 were further hydrolyzed for 2 h (abbreviated respectively as Alc, Try, and Neu) under the enzymatic conditions for Flavourzyme described in Table 1. At the end of the hydrolysis, all the enzymes were inactivated by heat treatment in boiling water for 15 min. Then all the hydrolysates were centrifuged at 10000 × g for 15 min at 4 °C. The supernatant was freeze-dried and stored at −20 °C for further analysis.
 |
| | Fig. 1 Diagram of enzymatic hydrolysis of wheat gluten for preparing a small-peptide powder with low-bitterness. | |
Determination of soluble nitrogen, peptide nitrogen and amino nitrogen contents
The α-amino nitrogen content in the supernatants was evaluated by formal titration,16 while the soluble nitrogen (SN) content was measured by the micro-Kjeldahl procedure (N × 5.7). The SN, peptide nitrogen (PN) and amino nitrogen (AN) contents were calculated using the following formulae:| | |
SN (%) = (nitrogen in the supernatant of hydrolysates/total nitrogen) × 100
| (2) |
| | |
AN (%) = (α-amino nitrogen/total nitrogen) × 100
| (3) |
Analysis of amino acid composition
The amino acid composition of the WG hydrolysates was determined according to the method reported by Fekkes et al.17 The amino acids in the samples were analyzed using an Agilent liquid chromatograph 1100 with a UV detector operated at 338 nm. The ODS Hypersil (250 × 4.6 mm) column was applied at 40 °C with the mobile phase delivered at a flow rate of 1 mL min−1. For the determination of free amino acids, WG hydrolysates (1.0 g) were filled up to 50 mL in volumetric flasks with trichloroacetic acid (10%, v/v). After incubation for 1 h at room temperature, the solution was filtered through Whatman filter paper no 4. The filtrate was centrifuged at 7000 × g for 10 min, and the supernatant was stored at 4 °C before injection. The total amino acids were determined after the WG hydrolysates (0.12 g) were hydrolyzed at 110 °C for 24 h with 6 mol L−1 HCl in evacuated sealed tubes. Then the sample (25 mL) was transferred to volumetric flasks and filtered. The filtrate was then transferred to a 10 mL beaker and placed in a vacuum desiccator to remove hydrochloric acid. The dried sample was then dissolved in 1 mL 0.02 mol L−1 HCl and kept at 4 °C before injection. External standards were used for quantification. The contents of different amino acids including free amino acids or the total amino acids were expressed as mg g−1 WG hydrolysates.
Determination of molecular weight distribution
The MW distribution profiles of the WG hydrolysates were determined by high-performance gel-filtration chromatography (HPGFC) on a TSK gel G2000 SWXL 7.8 × 300 mm column (Tosoh Co., Tokyo, Japan) with a Waters 1525 liquid chromatography system (Waters Co., Milford, MA, USA) according to the method of Liu et al.,18 with some minor changes. HPGFC was carried out with the mobile phase (acetonitrile/water/trifluoroacetic acid, 10/90/0.1, v/v/v) at a flow rate of 0.5 mL min−1 and monitored at 220 nm and 30 °C. The standards used from Sigma were cytochrome C (12.5 kDa), aprotinin (6.5 kDa), bacitracin (1450 Da), tetrapeptide GGYR (451 Da), and tripeptide GGG (189 Da).
Electronic tongue measurements
Electronic tongue measurements were carried out with the taste sensing system TS-5000Z (Insent Inc., Japan). This electronic tongue was equipped with five lipid membrane sensors indicating different taste qualities and the corresponding reference electrodes. These sensors represent the gustatory stimuli umami (SB2AAE), bitterness (SB2C00), saltiness (SB2CT0) and sourness (SB2CA0), and the nociceptive sensation, astringency (SB2AE1). The sensor outputs were consequently obtained as values. Each sample was measured four times with the sensors of bitterness and astringency. One measurement cycle measured a reference solution (Vr), then the sample solution (Vs), followed by a short (2 × 3 s) cleaning procedure, measurement of the aftertaste (V′r) of the reference solution, and a cleaning procedure for 330 s. The aftertaste was measured by determining the change of the membrane potential caused by the substance adsorption to the lipid membrane after the short cleaning procedure. The calculation formulae of the sensor output for taste and aftertaste were:| | |
Sensor output for taste = Vs − Vr
| (5) |
| | |
Sensor output for aftertaste = V′r − Vr
| (6) |
Sensory analyses
Sensory evaluation panelists were trained three times a week with quinine standards for a period of 1 month. Ten trained panelists (5 male and 5 female, aged between 20 and 35 years) were chosen to evaluate the bitter taste of the WG hydrolysates. The standard solutions were presented in concentrations of 0, 8.0 × 10−6, 1.6 × 10−5, 2.4 × 10−5, 3.2 × 10−5 and 4.0 × 10−5 g mL−1 that were scored as 0, 1, 2, 3, 4 and 5, respectively. The standard conditions for sensory evaluation were a room temperature of 25 °C for WG hydrolysates at a concentration of 1% (g mL−1) at pH 6.5. The final values given by the panelists were averaged.
Statistical analysis
All of the tests of the WG hydrolysates were performed in triplicate and the values were expressed as the mean ± standard deviation (SD). The analysis of variance (ANOVA) of the data was carried out using SPSS software (version 16.0, SPSS Inc., Chicago, IL, USA). The differences between the means were evaluated by Duncan's multiple range test with a confidence interval of 95%.
Results and discussion
Effect of protease and hydrolysis time on TD factors of WG hydrolysates
The bitter taste thresholds and the corresponding contents of bitter peptides determined the bitterness of the peptide powder. Fig. 2 presents the TD factors of the hydrolysates hydrolyzed by endopeptidases (Neutrase, Papain, Alcalase and Trypsin), which increased as the DH increased. During the process of continuous hydrolysis, the average relative MW of the WG hydrolysates rapidly decreased, which facilitated the production of bitter peptides in large amounts and the exposure of hydrophobic groups, resulting in geometric growth of the content of bitter peptides and playing a critical role in bitterness sensing.
 |
| | Fig. 2 Effect of protease and hydrolysis time on the taste dilution factors of wheat gluten hydrolysates. | |
The enzymatic substrates of exopeptidase were the peptides, while for endopeptidase it was the protein. Hence the effect of exopeptidase was different from that of endopeptidase since it couldn't significantly increase the content of bitter peptides. However, the exopeptidase had a special role in the bitter taste threshold of peptides, resulting in the change of the WG hydrolysates' TD factor. For Flavourzyme, it didn't have a high endo-activity for proteolysis; instead it had high exo-activity to cut hydrophobic amino acids from the C-terminal of the bitter components and catalyze the degradation of proteins into large MW peptides and free amino acids. Most of the degradants of Flavourzyme hydrolysates weren't bitter substances, and the TD factor remained at a low level before 8 h. When the reaction time reached 12 h, spoilage of the free amino acids was caused mainly by the activity of the microorganism generated putrid odor and sour/bitter tastes, which increased the TD factor of the hydrolysates. For Protamex, the exo-activity was worse than that of Flavourzyme. The TD factor of hydrolysates prepared by Protamex never decreased.
Previous research19,20 predicated that the bitterness rose because of an increasing DH and the content of bitter peptides. A bell curve relationship was discovered between the bitterness and the relative average MW of the peptide fraction.13 When the DH increased to a certain level, the relative average MW decreased which caused the bitterness to reduce. In the case of Proteax, it had both high endo- and exo-activities, and thus the TD factor of the hydrolysates prepared by Proteax reached the highest TD factor at 2 h, which decreased at 5 h and remained unchanged at 8 h. WG was mainly hydrolyzed into peptides, which contained a large number of bitter peptides, by Proteax after 2 h. A portion of the bitter peptides was further hydrolyzed to smaller peptides and free amino acids after 5 h. However the structure of the smaller peptides made it difficult for them to generate bitterness, leading to the decrease of the bitterness intensity. In the current research, the TD factor of Proteax hydrolysates no longer decreased after 8 h, suggesting that part of the bitter substances weren't degraded. Moreover, when the hydrolysis time was longer than 12 h, the spoilage of free amino acids generated a putrid odor and sour/bitter tasting substances. Thus, extending the treatment time of exopeptidase isn't an ideal way to completely eliminate bitterness.
Effect of DH on TD factors of WG hydrolysates
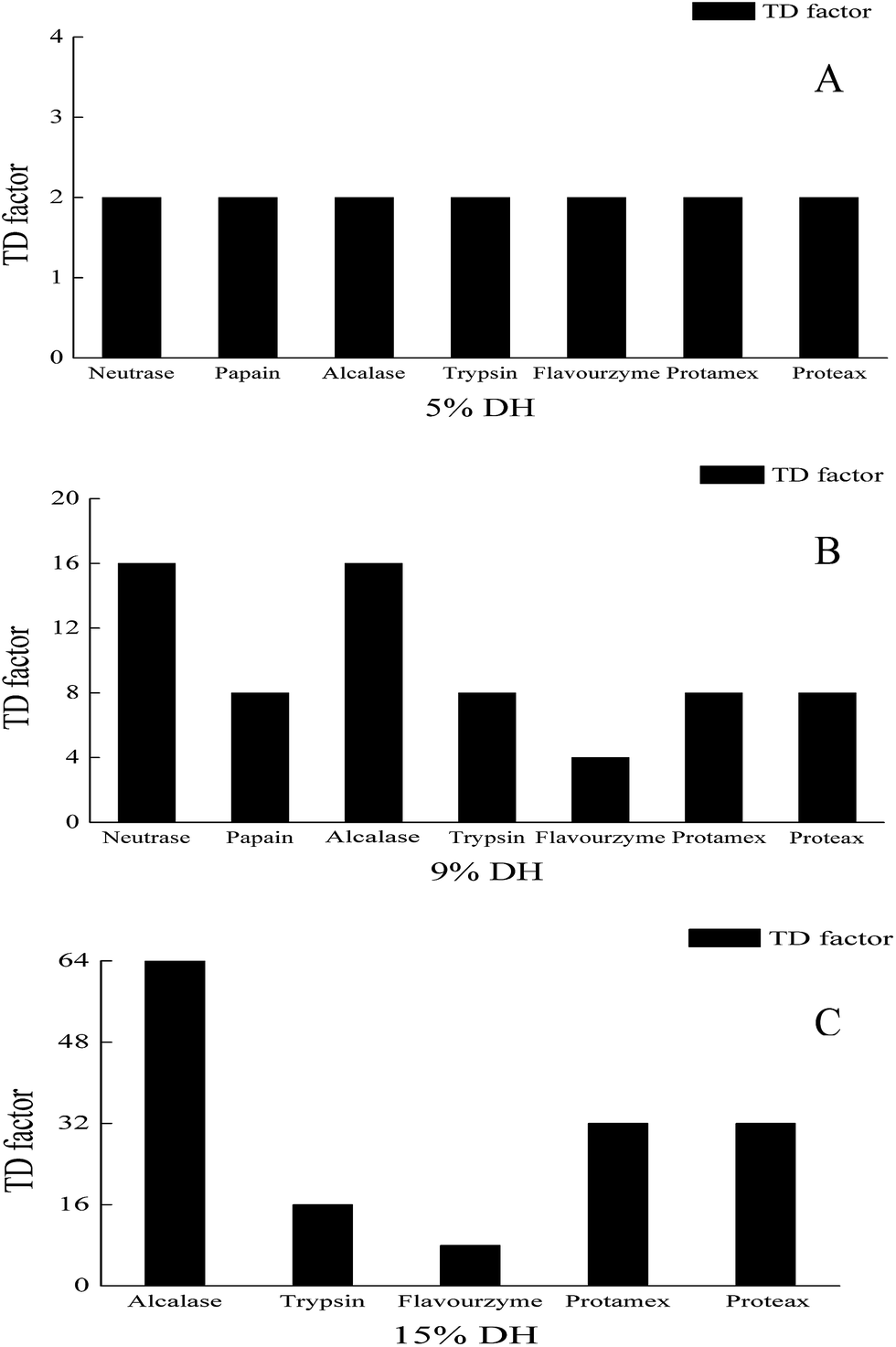
The content of the bitter peptides was relatively close for different WG hydrolysates with the same DH, the bitter taste thresholds of the bitter peptides determined the bitterness of the peptide powder. As shown in Fig. 3A, there was no perceived significant bitterness difference among the different WG hydrolysates at 5% DH. The hydrolysates contained a large number of high MW peptides with a low DH. Due to complex molecular configurations, the hydrophobic groups had difficulty in approaching taste receptors on the taste buds. The bitterness wasn't directly related to the enzyme cleavage sites and the bitter taste thresholds. However, Fig. 3B shows that the TD factors of the WG hydrolysates prepared by different proteases were different. The influence of the bitter taste threshold on the bitterness of WG hydrolysates was detected at 9% DH. The long-chain bitter peptides prepared by Papain and Trypsin had smaller bitter taste thresholds. Conversely, the short-chain bitter peptides prepared by Neutrase and Alcalase had smaller bitter taste thresholds.13 Neutrase and Alcalase had higher TD factors than Papain and Trypsin. The results indicate that the stereoscopic structure of long-chain peptides hindered the exposure of hydrophobic amino acids. Thus, the long-chain bitter peptides didn't play a major role in the bitterness intensity of the WG hydrolysates at 9% DH. In contrast, short-chain bitter peptides were the main contributor to the bitterness of the substances. Peptide bonds were preferentially cleaved by Proteax on the carboxyl-terminal of Gln, Ser, Thr and Met. Proteax had a lower TD factor than Neutrase and Alcalase.
 |
| | Fig. 3 Effect of the degree of hydrolysis on the taste dilution factors of wheat gluten hydrolysates. (A) 5% DH; (B) 9% DH; (C) 15% DH. | |
Fig. 3C shows that the effect of the bitter taste threshold on the bitterness intensity of the WG hydrolysates was more apparent, and the TD factors of the different WG hydrolysates had a greater disparity. Short peptides prepared by Alcalase had the lowest bitter taste threshold because of hydrophobic amino acids located at the C-termini of short peptides, so the TD factor was the highest after Alcalase treatment. The proline residue had the unique imino ring, which could change the steric distance between the binding unit (hydrophobic group composed of at least a three carbon skeleton) and the stimulating unit (a bulky basic group including an α-amino group) of bitter taste determinant sites21 by folding the peptide skeleton. According to the above principle, it was no wonder that the collaborative effect of the neighboring basic amino acids and proline residues generated the strong bitter taste. For Trypsin, it cleaved peptide bonds on the carboxyl-termini of Lys and Arg, which avoided the adjacent basic amino acid and proline residues; thus the TD factor was relatively low after treatment of Trypsin. Proteax finally cleaved the peptide bonds on the carboxyl-termini of Lys, Ala, Phe and Leu. The newly generated peptides had a low bitter taste threshold with their Phe and Leu groups located at the C-termini of the peptides. When the DH reached 15% within 2 h, the content of bitter peptides was the highest, and the debittering effect of the exopeptidases wasn't apparent and was still at the preliminary stage, so the TD factor was the same as that for Protamex.
Endopeptidase induced the production of a small-peptide powder, leading to an increased content of bitter peptides, which is the dominant factor deciding bitterness. Bitter peptides prepared by different proteases had different bitter taste thresholds which was the secondary factor affecting bitterness. The influence of the bitter taste threshold became gradually apparent as the content of bitter peptides and small peptides increased. For preparing a low-bitterness small-peptide powder with a high yield, a combined technology of endo- and exo-peptidase should be applied. Proteax had strong endo- and exo-activities, so Proteax hydrolysates obtained at 2 h, 5 h and 8 h were compared. Alcalase and Neutrase cleaved peptide bonds on the carboxyl-termini of hydrophobic amino acids and aromatic amino acids, respectively. Both Papain and Trypsin cleaved peptide bonds on the carboxyl-termini of basic amino acids, but Papain should be excluded because its low DH prohibited it from yielding enough small peptides. It was convenient to use Alcalase or Neutrase together with Flavourzyme to produce a low-bitterness small-peptide powder in a two-step enzymatic reaction. The appropriate time of hydrolysis should be screened.
Soluble nitrogen, peptide nitrogen and amino nitrogen contents of WG hydrolysates
As shown in Fig. 4, protein degradation had effectively improved the solubilization of insoluble macromolecular hydrophobic substances, resulting in a low loss rate of raw protein. Hence, the SN contents of WG hydrolysates were all more than 70%. With increasing hydrolysis time, Proteax's exo-activity became stronger which led to the AN content increasing gradually, causing the highest AN content of the Pro-h hydrolysate. Alcalase cleaved the peptide bonds on the carboxyl-termini of hydrophobic amino acids, which facilitated Flavourzyme further degrading the WG hydrolysates. Therefore Alc had the second highest AN content of all the enzyme solutions. Based on the nutritional information labels of the peptide series of products at home and abroad, the MWs of high-quality peptide powders are mostly less than 1000 Da. The PN content was directly related to the quality of the products and determined the value and the field of application of the products. The PN contents of all enzymatic hydrolysates were above 50%, and were 61.54% and 61.44%, for Pro-m and Alc, respectively. Sequential hydrolysis was a very good technical means for preparation of the peptide powder.
 |
| | Fig. 4 Soluble nitrogen, peptide nitrogen and amino nitrogen contents of wheat gluten hydrolysates. Different characters (a–c) represent significant difference of the soluble nitrogen content at p < 0.05 level among what gluten hydrolysates treated by different enzymatic treatment (Pro-l, Pro-m, Pro-h, Try, Alc and Neu). Different characters (d–g) represent significant difference of the peptide nitrogen content at p < 0.05 level among what gluten hydrolysates treated by different enzymatic treatment (Pro-l, Pro-m, Pro-h, Try, Alc and Neu). Different characters (h–m) represent significant difference of the amino nitrogen content at p < 0.05 level among what gluten hydrolysates treated by different enzymatic treatment (Pro-l, Pro-m, Pro-h, Try, Alc and Neu). | |
WG hydrolysates that had higher PN contents tended to have higher contents of bitter peptides. Neu treatment had the lowest PN content of all the schemes, which might give rise to the lowest content of bitter peptides. It is worth noting that the ratio between the PN content of Pro-m and Neu hydrolysates was much lower than that between hydrolysates with a high DH and hydrolysates with a low DH prepared by endopeptidase. Thus the differences between the contents of bitter peptides in all the schemes weren't enough to play a leading role in the bitterness intensity of the WG hydrolysates. The differences between the bitter taste thresholds of bitter peptides need to be analyzed.
Amino acid composition of peptides
The bitter amino acids22 all contained both the binding unit and the stimulating unit of bitter taste determinant sites. The bitter peptides tended to have a high proportion of hydrophobic amino acids. Therefore, the hydrophobic amino acid ratio and bitter taste amino acid ratio greatly influenced the bitter taste threshold of the bitter peptides. However, the effect of the total hydrophobic amino acid ratios and bitter taste amino acid ratios on the bitterness intensity of the WG hydrolysates wasn't certain. According to the result of the total hydrophobic amino acid ratios in Table 2, the bitterness intensity of Neu hydrolysates must have been kept at the lowest level. However Alc hydrolysates were just the opposite but this phenomenon was inconsistent with the sensory evaluation in Fig. 5. The total bitter taste amino acid ratio of peptides in the Pro-h hydrolysates was 37.60%, which was the lowest of all the schemes, so the bitter taste amino acid ratio of bitter peptides should also be the lowest. The bitterness intensity of Pro-h hydrolysates must have been kept at the lowest level but this phenomenon wasn't consistent with the sensory evaluation shown in Fig. 5. These results illustrate that the hydrophobic amino acids and bitter taste amino acids both contained many kinds of amino acids. The differences of the amino acid compositions of the bitter peptides between all the schemes weren't enough to affect the bitter taste thresholds of the bitter peptides. Perhaps the discrepancy of the bitter taste thresholds of the bitter peptides is induced by the amino acid sequence or the MW distribution.
Table 2 Amino acid composition of peptides (mg g−1 wheat gluten hydrolysates)a
| Peptide-bound amino acidsa |
Concentration (mg g−1) (mean ± SD) |
| Pro-l |
Pro-m |
Pro-h |
Alc |
Trp |
Neu |
| Different characters represent significant difference of peptide-bound amino acid at p < 0.05 level among what gluten hydrolysates treated by different enzymatic treatment (Pro-l, Pro-m, Pro-h, Try, Alc and Neu). aPeptide-bound amino acids were calculated by subtracting free amino acids from total amino acids. bAspartic acid + asparagine. cGlutamic acid + glutamine. dNot determined. eTotal hydrophobic amino acids contained Gly, Ala, Val, Leu, Pro, Met, Phe, Trp and Ile. The ratios of total hydrophobic amino acids were expressed as the percentage of the corresponding amino acids compared to the total amino acids. fTotal bitter baste amino acids contained Pro, Tyr, Lys, His, Arg, Val, Met, Phe, Ile, Leu, Try. The ratios of total bitter taste amino acids ratios were expressed as the percentage of the corresponding amino acids compared to the total amino acids. |
| Aspb |
27.21 ± 0.75c |
32.42 ± 1.60a |
30.23 ± 1.31a,b |
29.84 ± 0.98b |
27.25 ± 1.66c |
24.52 ± 1.25d |
| Gluc |
247.31 ± 11.65a,b |
254.65 ± 15.44a |
218.67 ± 14.15c |
251.26 ± 13.04a |
249.79 ± 13.40a |
224.62 ± 10.86b,c |
| Ser |
24.23 ± 1.38d |
28.07 ± 1.47a,b,c |
29.04 ± 1.50a,b |
30.47 ± 1.23a |
26.12 ± 1.14c,d |
26.77 ± 1.65b,c,d |
| Ala |
13.42 ± 1.06a |
14.04 ± 1.31a |
10.45 ± 1.01b |
15.19 ± 0.87a |
13.08 ± 1.11a |
10.47 ± 1.30b |
| Gly |
20.96 ± 1.44a,b |
22.21 ± 1.63a |
18.44 ± 1.23b |
22.45 ± 1.00a |
20.61 ± 1.42a,b |
18.66 ± 1.15b |
| Thr |
9.54 ± 1.07a |
9.32 ± 1.21a,b |
4.13 ± 0.76d |
6.82 ± 0.81c |
7.91 ± 0.45b,c |
6.76 ± 0.26c |
| Pro |
92.15 ± 9.19b |
103.70 ± 5.36a,b |
95.43 ± 8.66b |
101.04 ± 9.56a,b |
111.75 ± 8.55a |
88.27 ± 5.25b |
| Tyr |
14.94 ± 1.10a |
11.87 ± 1.20b |
8.67 ± 0.76d |
9.16 ± 1.01c,d |
10.74 ± 1.21b,c |
10.09 ± 0.64b,c,d |
| Lys |
6.29 ± 0.49a |
6.55 ± 0.50a |
4.78 ± 0.76c |
5.78 ± 0.59a,b,c |
6.19 ± 0.66a,b |
5.16 ± 0.30b,c |
| His |
10.09 ± 1.64a |
10.12 ± 1.10a |
6.72 ± 1.14b |
10.49 ± 0.75a |
10.10 ± 0.23a |
8.62 ± 0.38a |
| Arg |
15.61 ± 1.01a |
15.75 ± 1.71a |
8.99 ± 1.33c |
14.02 ± 0.81a,b |
14.58 ± 1.17a,b |
12.37 ± 0.98b |
| Val |
23.89 ± 0.98a |
24.22 ± 0.93a |
16.35 ± 1.66b,c |
22.73 ± 1.04a |
18.09 ± 2.07b |
13.94 ± 1.47c |
| Met |
7.18 ± 0.79a |
5.52 ± 0.85b |
1.75 ± 0.65d |
3.50 ± 0.61c |
4.24 ± 0.49c |
3.44 ± 0.38c |
| Phe |
26.66 ± 1.80b,c |
28.43 ± 1.90a,b |
20.83 ± 1.85d |
31.67 ± 1.90a |
25.41 ± 3.52b,c |
23.66 ± 2.43c,d |
| Ile |
22.64 ± 1.06a |
22.29 ± 1.60a |
15.05 ± 1.46b,c |
23.65 ± 1.37a |
16.84 ± 2.00b |
12.86 ± 1.57c |
| Leu |
25.25 ± 2.62a |
21.60 ± 1.47a |
11.00 ± 3.09b |
26.62 ± 1.48a |
21.19 ± 3.99a |
14.49 ± 3.46b |
| Try |
n.dd |
n.dd |
n.dd |
n.dd |
n.dd |
n.dd |
| Cys |
5.95 ± 0.58b |
6.57 ± 0.29a |
3.72 ± 0.26e |
4.25 ± 0.36d,e |
5.28 ± 0.21c |
4.68 ± 0.10d |
| Total |
593.30 ± 9.92b,c |
617.32 ± 11.62a |
504.23 ± 9.77d |
608.93 ± 8.19a,b |
589.16 ± 6.10c |
509.38 ± 8.55d |
| Total hydrophobic amino acid ratiose (%) |
39.13 ± 0.99a,b |
39.20 ± 1.20a,b |
37.54 ± 1.68b |
40.54 ± 2.04a |
39.25 ± 1.53a,b |
36.48 ± 1.44b |
| Total bitter taste amino acid ratiosf (%) |
41.24 ± 0.73a |
40.50 ± 1.51a,b |
37.60 ± 1.53c |
40.83 ± 2.41a |
40.59 ± 1.40a,b |
37.87 ± 1.09b,c |
 |
| | Fig. 5 Bitterness intensity scores of wheat gluten hydrolysates on the 5 point intensity scale. Different characters (a–h) represent significant difference of taste intensity at p < 0.05 level among what gluten hydrolysates treated by different enzymatic treatment (Try-1, Alc-2, Neu-6, Try, Alc, Neu, Pro-l, Pro-m and Pro-h). | |
Molecular weight distribution of WG hydrolysates
Table 3 shows the largest fractions of the peptides with a MW ranging from 180–500 Da. Compared to the conventional technology using the single endopeptidase or exopeptidase,23 sequential hydrolysis with endo- and exo-peptidase has greatly improved the content of small peptides, and strengthened the competitiveness of the peptide powder. Pro-l treatment gave the lowest content of the peptide fraction below 180 Da compared to the other treatments, so Pro-l hydrolysates had greater potential for biological activity. However, for the peptide fraction below 6 kDa, the peptides that had a longer peptide chain tended to have the smaller bitter taste thresholds on the premise of containing a high proportion of hydrophobic amino acids. Pro-l hydrolysates had a high proportion of the peptide fraction with a MW ranging from 500–3000 Da, which might contain a larger amount of strong bitter peptides than for the other schemes. Pro-m hydrolysates had the highest content of small peptides ranging from 180–500 Da, which favored the enrichment of small peptides. In addition to these advantages, Pro-m hydrolysates had a low proportion of the peptide fraction with a MW ranging from 500–3000 Da, so the bitter taste thresholds of bitter peptides prepared by Pro-m treatment were relatively low.
Table 3 Molecular weight distribution of wheat gluten hydrolysatesa
| Molecular weight (Da) |
Pro-l |
Pro-m |
Pro-h |
Trp |
Alc |
Neu |
| Different characters represent significant difference of molecular weight at p < 0.05 level among what gluten hydrolysates treated by different enzymatic treatment (Pro-l, Pro-m, Pro-h, Try, Alc and Neu). |
| ≥10000 |
1.24 ± 0.09a |
0.82 ± 0.03c |
0.56 ± 0.06e |
1.08 ± 0.03b |
0.67 ± 0.02d |
1.17 ± 0.03a |
| 6000–10000 |
4.88 ± 0.37a |
2.40 ± 0.15d |
1.41 ± 0.03f |
3.27 ± 0.06c |
1.93 ± 0.03e |
3.86 ± 0.05b |
| 3000–6000 |
1.73 ± 0.11d |
2.35 ± 0.16b |
1.17 ± 0.10e |
2.74 ± 0.09a |
2.15 ± 0.08c |
1.56 ± 0.11d |
| 1000–3000 |
11.42 ± 0.27a |
4.89 ± 0.40d |
3.15 ± 0.17e |
7.63 ± 0.44c |
4.53 ± 0.14d |
8.21 ± 0.17b |
| 500–1000 |
22.92 ± 1.63a |
11.75 ± 1.03d |
10.53 ± 0.73d |
15.25 ± 0.64c |
11.59 ± 0.64d |
18.07 ± 0.53b |
| 180–500 |
44.37 ± 1.36d |
53.26 ± 2.36a |
48.43 ± 1.12c |
49.28 ± 0.75b,c |
51.29 ± 0.93a,b |
44.96 ± 0.86d |
| ≤180 |
13.44 ± 1.14e |
24.53 ± 2.02c |
34.75 ± 2.19a |
20.75 ± 1.91d |
27.84 ± 1.76b |
22.17 ± 1.72c,d |
Sensory characterization of WG hydrolysates
To elaborate the debittering mechanism, the bitter intensity of Try-l, Alc-2 and Neu-6 hydrolysates prepared by single endopeptidase treatment were considered. As shown in Fig. 5, Alc-2 hydrolysates showed the highest bitter scores, followed by the hydrolysates of Pro-l, Try-1, Neu-6, Neu, Alc, Pro-h, Try and Pro-m. The DH values of Neu-6, Try-1, Pro-l and Alc-2 hydrolysates were 9.03%, 14.64%, 15.15% and 16.06%, respectively. Endopeptidase induced hydrolysis of WG and increased the content of bitter peptides, which was the dominant factor affecting the bitterness. This conclusion was further affirmed by the bitterness intensity scores of the four kinds of hydrolysates. The DH values of Alc-2 and Pro-l hydrolysates were relatively close, but their bitter scores were significantly different. The results indicated that the bitter taste thresholds of the bitter peptides of the Alc-2 hydrolysates were much lower than those of the Pro-l hydrolysates. With hydrolysates corresponding to a high degree of hydrolysis, the influence of the bitter taste thresholds on the bitterness gradually became apparent. The exopeptidase was effective to reduce the bitter taste threshold of bitter peptides. After effective chain-end degradation of Proteax and Flavourzyme, the bitterness intensity scores of Neu, Alc, Pro-h, Try and Pro-m hydrolysates were all below 3. The sequential hydrolysis was an ideal method to prepare low-bitterness small-peptide powders by endo- and exo-peptidase treatment.
During sequential hydrolysis, the WG hydrolysates reached a high degree of hydrolysis. The content of the peptide fraction with a large MW was low, suggesting the effect of the stereoscopic structure of long-chain peptides on hindering hydrophobic amino acid exposure was negligible. In addition, the differences of the amino acid composition of bitter peptides between all schemes weren't enough to affect the bitter taste threshold of the bitter peptides. The differences of the bitter taste thresholds of bitter peptides between all schemes were caused by the amino acid sequences and the MWs of the bitter peptides. When proline was among the basic amino acids, small peptides also elicited a strong bitter taste. In particular, the proline content was the second highest among the detected amino acids of the WG hydrolysates (Table 2). The particular amino acid sequence should be given more attention. Proteax and Trypsin cleaved peptide bonds on the carboxyl-termini of Lys and Arg, which destroyed the particular amino acid sequences. Therefore, the bitter scores of Pro-m, Pro-h and Try hydrolysates were relatively low. Through the comparison of the MW distribution of Pro-m, Pro-h and Try hydrolysates, the bitter score of Pro-h hydrolysates should be the lowest, which isn't consistant with the result. The most likely explanation is that spoilage of free amino acids generated a small amount of putrescine and cadaverine, which increased the bitterness.
Representation of sensor response of the electronic tongue
To compare the intensity of the sensor, the initial sense output of Try-1 treatment was defined as zero, being the frame of reference. The initial sensor outputs of other treatments were converted into new voltage values by a TS-5000Z software system. A greater number represented a stronger taste intensity. The intensity of the bitter sense is a measure of the size of the bitter taste produced by the bitter substance in a certain period of time. The sensor output of bitterness represented the instantaneous maximum intensity of the bitter taste. The sensor output of the bitter aftertaste (aftertaste-B) represented a sustained intensity of bitter taste, which was correlated to the hydrophobicity of bitter peptides. The sensor outputs of astringency and astringent aftertaste (aftertaste-A) were closely correlated to the hydrophobicity of bitter peptides, so these results were considered to analyze the sustained intensity of bitterness.
As shown in Fig. 6, the sensor output of bitterness gave the same results as those obtained with the sensory evaluation. The sensor outputs of aftertaste-B, astringency and aftertaste-A were closely correlated to each other, which wasn't consistent with the result obtained by the sensor output of bitterness. These results indicate that the instantaneous maximum intensity of bitter taste had a more important effect on the sensory evaluation than a sustained intensity of the bitter taste. According to the results of the sensor outputs of aftertaste-B, astringency and aftertaste-A in Table 4, the hydrophobicity of bitter peptides produced by Try treatment was stronger than that for Pro-m treatment. Try hydrolysates had a stronger sustained intensity of bitter taste than Pro-m hydrolysates, resulting in a higher bitterness intensity score for Try than Pro-m.
 |
| | Fig. 6 Radar distribution results of sensor responses of wheat gluten hydrolysates obtained by the electronic tongue. | |
Table 4 The sensor outputs of wheat gluten hydrolysates obtained by the electronic tonguea
| |
Bitterness (mV) |
Aftertaste-B (mV) |
Astringency (mV) |
Aftertaste-A (mV) |
| Different characters represent significant difference of the sensor outputs at p < 0.05 level among what gluten hydrolysates treated by different enzymatic treatment (Try-1, Alc-2, Neu-6, Try, Alc, Neu, Pro-l, Pro-m and Pro-h). |
| Try-1 |
0.000 ± 0.000c |
0.000 ± 0.000b |
0.000 ± 0.000b |
0.000 ± 0.000b |
| Alc-2 |
3.440 ± 0.145a |
0.640 ± 0.044a |
−0.557 ± 0.152d |
−0.300 ± 0.079c |
| Neu-6 |
−0.103 ± 0.086c |
0.563 ± 0.061a |
2.520 ± 0.070a |
0.890 ± 0.141a |
| Try |
−1.320 ± 0.070g |
−0.277 ± 0.006d |
−0.703 ± 0.150d |
−0.550 ± 0.061d |
| Alc |
−0.603 ± 0.064e |
−0.240 ± 0.089c,d |
−1.083 ± 0.163e |
−0.687 ± 0.106e,f |
| Neu |
−0.433 ± 0.067d |
−0.137 ± 0.110c |
−0.647 ± 0.134d |
−0.340 ± 0.026c |
| Pro-l |
1.467 ± 0.040b |
−0.127 ± 0.153b,c |
−0.223 ± 0.087c |
−0.273 ± 0.051c |
| Pro-m |
−1.917 ± 0.045h |
−0.327 ± 0.038d |
−0.717 ± 0.133d |
−0.563 ± 0.068d,e |
| Pro-h |
−0.900 ± 0.066f |
−0.363 ± 0.029d |
−1.193 ± 0.142e |
−0.730 ± 0.036f |
Conclusions
The bitter taste thresholds and the corresponding contents of bitter peptides determined their role in the bitterness of peptide powders prepared by endopeptidases. The content of bitter peptides increased at geometric rates and played a leading role in the bitterness. Bitter peptides prepared by different proteases had different bitter taste thresholds, which was the second important fact affecting the bitterness, the influence of which became gradually apparent as the DH increased. For preparing a low-bitterness small-peptide powder with a high yield, a combined technology of endo- and exo-peptidase treatment was attempted in this study. During the sequential hydrolysis, the differences of bitterness between all the schemes were determined by the bitter taste thresholds of the bitter peptides, but not the content of bitter peptides. The differences of the bitter taste thresholds of bitter peptides between all the schemes was caused by the amino acid sequences and MWs of the bitter peptides.
Overall, in all the schemes, Pro-m hydrolysates exhibited the best characteristics, having the highest content of small peptides ranging from 180–500 Da, with SN and PN contents of 82.82% and 61.54%, respectively, and a lower bitterness intensity score than for other schemes. In the future, more attention should be focused on the bitter taste threshold when preparing low-bitterness small-peptide powders by sequential hydrolysis with endo- and exo-peptidase.
Acknowledgements
The study was supported by National High Technology Research and Development Program of China (863 Program, No. 2013AA102201), the Key R & D Programs of Jiangsu Province (BE2015327) and the Jiangsu province “Collaborative Innovation Center for Modern Grain Circulation and Safety” industry development program.
Notes and references
- I. J. Joye and D. J. McClements, J. Agric. Food Chem., 2014, 62, 2623–2630 CrossRef CAS PubMed.
- R. E. Cian, J. Vioque and S. R. Drago, Food Res. Int., 2015, 69, 216–223 CrossRef CAS.
- S. L. Lahrichi, M. Affolter, I. S. Zolezzi and A. Panchaud, J. Proteomics, 2013, 88, 83–91 CrossRef CAS PubMed.
- K. Horibe, Nihon Rinsho Geka Gakkai Zasshi, 1987, 88, 808–815 CAS.
- H. Hara, R. Funabiki, M. Iwata and K. Yamazaki, J. Nutr., 1984, 114, 1122–1129 CAS.
- E. R. Gilbert, E. A. Wong and K. E. Webb Jr, J. Anim. Sci., 2008, 86, 2135–2155 CrossRef CAS PubMed.
- K. H. Ney, Zeitschrift für Lebensmittel-Untersuchung und -Forschung, 1971, 2, 64 CrossRef.
- T. K. Murray and B. E. Baker, J. Sci. Food Agric., 1952, 3, 470–475 CrossRef CAS.
- N. Ishibashi, I. Ono, K. Kato, T. Shigenaga, I. Shinoda, H. Okai and S. Fukui, Agric. Biol. Chem., 1988, 52, 91–94 CrossRef CAS.
- N. Ishibashi, Y. Arita, H. Kanehisa, K. Kouge, H. Okai and S. Fukui, Agric. Biol. Chem., 1987, 51, 2389–2394 CrossRef CAS.
- N. Ishibashi, K. Sadamori, O. Yamamoto, H. Kanehisa, K. Kouge, E. Kikuchi, H. Okai and S. Fukui, Agric. Biol. Chem., 1987, 51, 3309–3313 CrossRef CAS.
- R. J. FitzGerald and G. O'Cuinn, Biotechnol. Adv., 2006, 24, 234–237 CrossRef CAS PubMed.
- M. J. Cho, Doctor, Characterization of bitter peptides from soy protein hydrolysates, University of Missouri-Columbia, 2000 Search PubMed.
- J. Adler-Nissen, Enzymic hydrolysis of food proteins, Elsevier Applied Science Publishers, New York, 1986 Search PubMed.
- W. H. Seo, H. G. Lee and H. H. Baek, J. Food Sci., 2008, 73, S41–S46 CrossRef CAS PubMed.
- S. Nilsang, S. Lertsiri, M. Suphantharika and A. Assavanig, J. Food Eng., 2005, 70, 571–578 CrossRef.
- D. Fekkes, A. Vandalen, M. Edelman and A. Voskuilen, J. Chromatogr. B: Biomed. Sci. Appl., 1995, 669, 177–186 CrossRef CAS PubMed.
- P. Liu, M. Huang, S. Song, K. Hayat, X. Zhang, S. Xia and C. Jia, Food Bioprocess Technol., 2010, 5, 1775–1789 CrossRef.
- G. B. Quaglia and E. Orban, J. Sci. Food Agric., 1987, 38, 271–276 CrossRef CAS.
- H. Geisenhoff, Master, Bitterness of soy protein hydrolysates according to molecular weight of peptides, Iowa State University, 2009 Search PubMed.
- N. Ishibashi, T. Kubo, M. Chino, H. Fukui, I. Shinoda, E. Kikuchi and H. Okai, Agric. Biol. Chem., 1988, 52, 95–98 CrossRef CAS.
- L. Liao, C. Y. Qiu, T. X. Liu, M. M. Zhao, J. Y. Ren and H. F. Zhao, J. Cereal Sci., 2010, 52, 395–403 CrossRef CAS.
- J. S. Wang, M. M. Zhao, Q. Z. Zhao, Y. Bao and Y. M. Jiang, J. Food Sci., 2007, 72, C103–C107 CrossRef CAS PubMed.
|
| This journal is © The Royal Society of Chemistry 2016 |
Click here to see how this site uses Cookies. View our privacy policy here.