
Cellulose as an efficient matrix for lipase and transaminase immobilization†
Abstract
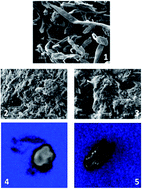
Immobilization of enzymes is important to improve their stability and to facilitate their recyclability, aiming to make biocatalytic processes more efficient. One of the important aspects is the utilization of cheap, abundant, and environmentally friendly carriers for enzyme immobilization. Here we report the use of functionalized cellulose for lipase and transaminase immobilization. High immobilization efficiencies (up to 90%) could be achieved for the transaminase from Vibrio fluvialis. For immobilized lipase CAL-B as well as the transaminase, good conversions and recyclability could be demonstrated in kinetic resolutions to afford chiral alcohols or amines. Moreover, such application of the immobilized transaminase enabled very high conversions in a continuous-flow process in the asymmetric synthesis of (S)-phenylethylamine (80% conversion, >99% ee).
 Please wait while we load your content...
Please wait while we load your content...

