
 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a Creative Commons Attribution 3.0 Unported Licence
Sodium ion interaction with psyllium husk (Plantago sp.)†
M. A.
Jimoh
a,
W.
MacNaughtan
a,
H. E. L.
Williams
b,
D.
Greetham
a,
R. L.
Linforth
a and
I. D.
Fisk
 *a
*a
aDivision of Food Sciences, School of Biosciences, University of Nottingham, Sutton Bonington Campus, Loughborough, Leicestershire LE12 5RD, UK. E-mail: Ian.fisk@nottingham.ac.uk; Fax: +44 (0)115 95 16142; Tel: +44 (0)115 95 16037
bCentre for Biomolecular Sciences, School of Chemistry, University of Nottingham, UK
First published on 31st August 2016
Abstract
The nature of and factors effecting sodium interactions with psyllium were investigated in vitro. In a batch extraction system, psyllium mucilage gel retained at least 50% of sodium across a range of concentrations (5–300 mg sodium per g psyllium) and pH (2–10) environments. FTIR and Na NMR analyses of psyllium gels indicated that binding was complex with non-specific multi-site interactions. The potential use of psyllium husk as a binding agent for the reduction of bioavailable sodium was therefore evaluated. The binding of sodium at physiologically relevant conditions (pH 1.2 (stomach) and 6.8 (intestine)) was studied in a gastrointestinal tract (GIT) pH simulated model. Results show consistently high sodium retention (∼50%) across the GIT model and less than 20% loss of bound sodium under the simulated intestinal pH conditions after repeated washings.
Introduction
Since the introduction of the UK salt reduction program in 2003,1 the consumption of salt in the population has been reduced by 15% according to data from the National Diet and Nutrition Survey.2 However with salt consumption at ∼8 g per day, salt consumption is yet to reach the recommended target of 5 g per day for adults. A high salt (sodium) diet increases the risk of high blood pressure which in turn increases the chance of having a heart attack or stroke. Reducing population salt intake by 1 g per day has been proposed to mitigate 4147 premature deaths and will save the National Health Service (NHS) £288 million annually.3 The World Health Organization (WHO) has pledged its support by developing tools for the identification of population salt consumption and major sources of sodium in the diet; support for the reformulation of a set number of products on the market; support in developing health communication material to increase consumer awareness on salt/sodium and to inform consumers on how to read and interpret food labels.4Sodium has multiple roles within food and as such salt reduction strategies are required to be multifaceted. One common approach is the replacement of sodium with similar salty tasting ions such as potassium and ammonium; unfortunately these salts carry additional flavours and aftertastes and therefore cannot be used as a direct complete replacement for sodium.
Sodium plays an anti-microbial and textural function in processed foods and the impact of sodium reduction on these functions has yet to be fully resolved. The use of flavour enhancers that amplify the perceived salty taste of sodium chloride, or replicate the flavour enhancing properties of sodium ions, include yeast extracts and specific nucleotide and glutamate preparations. Whilst these are partially successful, there is often an impact on production costs and require modifications to nutritional labels. One example being the use of seaweed as a potential salt replacer.5
A related approach to reduce sodium content in food is by gradual reduction of the salt content over an extended time period, typically months or years.6 This reduction by stealth requires industry-wide cooperation and long time periods to achieve significant reductions.
Most research approaches in our laboratories7–9 and indeed elsewhere,10–12 have focused on optimising the delivery or bioaccessibility of sodium (e.g. through studies on mitigating carriers to dissolution, diffusion and effecting mixing).
The work presented here aims to tackle the issue of salt reduction in the diet through modifying the (bio) availability of sodium (molecular binding). Our current hypothesis is that the binding of sodium to natural products such as polysaccharides (especially indigestible dietary fibres) would serve to reduce dietary sodium bioavailability and increase sodium excretion through the faecal route.
Natural polysaccharides like other biopolymers, have gained interest in the food and pharmaceutical industries due to their versatility and biocompatibility/biodegradability. They have found use as stabilizers, thickeners, binders, dispersants, lubricants, adhesives and drug-delivery agents, amongst other applications. There is ongoing interest in the use of plant based polysaccharides as a functional food ingredient in the prevention and treatment of obesity, diseases such as cardio-vascular disease and cancer e.g. (ref. 13) and references therein.
Psyllium (Plantago sp.) seeds and husk have gel forming and dietary bulking properties due to the seeds being coated in an external polysaccharide mucilage layer (husk). Psyllium husk has been used as a laxative for the treatment of irritable bowel disorder (IBD) symptoms and as a health supplement. Psyllium husk and its derivatives have also found application for targeted drug release e.g. as a vehicle for the targeted delivery to the colon.14 Environmental applications have included its use as natural flocculation agent in textile wastewater remediation15 and as a heavy metal ion scavenger. The mucilage from psyllium husk (Plantago ovata) is mainly composed of arabinose and xylose sugars in a highly branched polymer configuration.16–21
The aim of this study was to evaluate psyllium husk as a binding agent for the reduction of bioavailable sodium. The binding of sodium to psyllium was evaluated across a gastrointestinal tract (GIT) model to identify if psyllium husk has the potential to reduce total bioavailability of sodium.
Materials and methods
Psyllium samples (Elf Foods, Loughborough, UK), were stored in plastic air-tight sample containers, and used within their indicated shelf-life (psyllium seeds and husk Sidhpur SAT-ISABGOL Factory brand both packaged by East Foods Ltd).Sodium chloride, potassium hydroxide, ammonium chloride, ammonium hydroxide, hydrochloric and nitric acid were at least Analar® grade (>98% purity) and obtained from Fisher Scientific, Loughborough UK. TraceMetal Grade nitric acid (evaluated for 65 metals <1 ppb) was used for wet digestion prior to total trace element analysis. Ionic strength adjuster (ISA) was 21% ammonium chloride in 14% ammonium hydroxide solution.
Elemental analysis and material characterisation by FTIR
Total sodium, potassium, calcium and magnesium content were determined by inductively coupled plasma atomic emission spectrometry (ICP-AES) using a Perkin Elmer Optima 2000 DV after microwave digestion (Anton Paar microwave oven) of 200 mg psyllium seed and husk samples with concentrated nitric acid (6 mL). The digests were diluted with ultrapure water to 40 mL and analysed for sodium (589.592 nm line), potassium (766.49, 404.721 nm lines), calcium (315.887, 317.933 nm lines) and magnesium (279.077, 285.213 nm lines). Analysis was in triplicate with reagent blanks. Quantification was against standards run in the concentration range of 0–20 ppm.Operationally defined water extractible sodium fraction and acid extractible fraction were determined by carrying out parallel extractions of 500 mg of psyllium seed and husk samples with 5 mL of ultrapure water (18.2 MΩ) and 0.02 M hydrochloric acid respectively and analysis of extracts obtained for sodium.
CHN analysis of psyllium seed and husk samples was carried out on 2 mg samples by an external laboratory (Laboratory Services, Chemistry Department, University of Nottingham) using a Perkin Elmer CHN analyser.
Fourier Transform Infra-red Spectroscopy (FTIR) measurements were carried out using a Tensor 27 spectrometer (Bruker, Coventry UK) equipped with a deuterated triglycine sulfate (DTGS) detector in attenuated total internal reflectance (ATR) mode. Single spectra of ground seed (355 μm) and husk samples, as well as monosaccharide standards (arabinose, xylose and galactose) and xylan were recorded between 600 and 4000 cm−1, at 4 cm−1 resolution and resulted from the co-addition of 128 scans. The raw spectra were baseline corrected and vector normalised using the OPUS software.
Evaluation of sodium interaction with psyllium
For estimation of bound fraction, a general extraction procedure was developed in which 500 ± 1 mg of sample was shaken with 5 mL of test solution containing a known amount of sodium. 5 mL of ultrapure water or aqueous surfactant solution (Sherwood photometer diluent solution) was added and the mixture shaken for 30 min and solids separated by centrifugation at 412g rpm for 20 min (multi-function centrifuge, Thermo Electron Corp.). The supernatant was removed for instrumental analysis to estimate free sodium. Sodium analysis was by flame photometry (Flame photometer Model 410 Sherwood Scientific Ltd, Cambridge, UK) or potentiometry (Ion meter (Jenway pH meter 3023) with sodium ion selective electrode (Cole-Parmer, London UK)). For flame photometric analysis, 2 mL of sample (supernatant) was diluted as required using the diluent solution to be in the linear range 1–10 ppm of the instrument. A direct comparison between sample reading and those of calibration standards gave the concentration of sodium. For potentiometric analysis, 200 μL of ionic strength adjuster (ISA) was added to 2 mL of supernatant and the mixture analysed. Standards of same volume were also analysed after addition of ISA. Bound sodium was calculated as difference between total sodium added and the free sodium measured. % Retention or % bound sodium was calculated from eqn (1) below| % Bound = 100 × (total-free)sodium/total sodium | (1) |
Influence of sample preparation
To investigate the influence of particle size, type of grinding and properties of seed on the interaction of Plantago sp. with sodium, seed samples were ground using either a laboratory disc-mill (Perten AB disc mill LM 3310, Hägersten Sweden) for ‘milled’ sample or a coffee grinder (DeLonghi KG series, Hampshire, UK) for ‘ground’ sample. The ground material was sieved through a series of laboratory sieves (Endecotts Ltd, London, England) of mesh sizes 106–355 μm and size fractions collected. Psyllium husk samples were used as obtained. Hydrogels were developed by adding 100 mL of ultrapure water to 1–2 g of psyllium husk. The mixture was dried overnight at 40° C, ground and carried through sodium binding experiments for comparison.Evaluation of the role of pH on sodium interaction
To study the effect of pH on the retention of sodium by psyllium husk, the general extraction procedure described above was followed except that the pH of test Na standard solutions (0.1% w/w) was adjusted to 2, 4, 6, 8 and 10 using dilute hydrochloric acid and potassium hydroxide solutions. Final pH of the mixture was checked with a pH meter (InoLab Level 1 pH meter, VWR Germany).Analysis of binding capacity
To estimate binding capacity, 500 mg of psyllium husk was mixed with 25 mL of solutions with various starting sodium concentrations (5–300 mg g−1) and the general extraction procedure applied as before.Analysis of sodium retention at physiologically relevant pH
To test the theoretical suitability of psyllium husk (Plantago sp.) as a scavenger of sodium in food, the retention behaviour of psyllium at the physiologically relevant pH of 1.2 for the stomach and 6.8 for the small intestine was evaluated. In this experiment, synthetic gastric juice formulations and buffers were avoided as they contained high levels of sodium. Thus 500 mg of husk sample was mixed with 5 mL of 0.1% w/w test sodium solution and 20 mL of ultrapure water. The mixture was centrifuged (412g) and an aliquot of the supernatant taken for analysis. The pH of the remaining mixture was adjusted to pH 1.2 using hydrochloric acid, thus simulating stomach pH conditions. The mixture was again centrifuged (412g) and an aliquot of the supernatant taken for sodium analysis. Thereafter, a further 5 mL of 0.1% w/w sodium solution was added, (to adjust the final volume and simulate the reflux of sodium in the intestine), then the pH was adjusted to 6.8 with dilute potassium hydroxide solution, simulating pH conditions in the intestine. This mixture was separated by centrifugation as before and the concentration of sodium in the supernatant determined as before.To test the release of ‘psyllium bound’ sodium in the intestine, 5 mL of a 1% w/w sodium solution was added to 500 mg psyllium husk (100 mg sodium per g psyllium), shaken and the resulting ‘sodium loaded’ psyllium husk gel mixed with 20 mL of ultrapure water. After centrifugation, the supernatant was removed and sodium content determined. Further washings with 10 mL portions of ultrapure water followed. Sodium content in each washing step was also determined and compared.
Analysis of psyllium gels by FTIR
To investigate the nature of interaction and binding sites of sodium in psyllium, FTIR-ATR spectra of psyllium husk gels prepared under different treatment conditions were obtained using a Bruker spectrometer as previously described. Gels were prepared by adding 5 mL of test solution to 500 mg of husk and shaking (30 min). Acid and alkali treatment gels were prepared by adding 0.1 M HCL and 0.1 M KOH to husk samples respectively. For the sodium treatment gel, 1% (w/v) sodium solution was added. Sodium–calcium mixed gels contained 2.5 mL of 1% (w/v) sodium and 1% (w/v) calcium solutions. Calcium treatment gel contained 1% calcium solution. A water gel control was prepared by mixing 500 mg husk and 5 mL ultrapure water, a further sample control of sodium chloride solution was taken.Analysis of sodium binding by 23Na-NMR
Na-NMR spectra were recorded using a R4-1004 compact Na-NMR spectrometer (AMR Ltd, Oxfordshire). Scans (4k) were recorded with a 90° pulse width of 20 μs and a relaxation delay of 100 ms. For the experiments, psyllium gels prepared in 0.1–5% (w/w) sodium chloride solutions were weighed into NMR tubes and left to equilibrate at 20 °C prior to measurement. Standard sodium chloride solutions and blanks were also analysed.Na single quantum (SQ) and double quantum filtered (DQF) NMR experiments22 were recorded at 158 MHz on a 600 MHz Bruker Avance III spectrometer equipped with a 5 mm broadband probe. Samples contained D2O (1![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :10) to obtain a lock signal. DQ coherences were selected using a phase cycled pulse sequence. Optimum creation time for maximum amplitude of the DQ signal was found by locating the maximum intensity for spectra acquired at different creation times.
:10) to obtain a lock signal. DQ coherences were selected using a phase cycled pulse sequence. Optimum creation time for maximum amplitude of the DQ signal was found by locating the maximum intensity for spectra acquired at different creation times.
Data analysis
Statistical data analysis was carried out using the GraphPad Prism 6 statistical package (GraphPad Software, Inc.), Origin (OrignlabInc) and Excel (Microsoft Corp). Experiments were carried out in triplicate and values reported are means ± standard deviation.Results and discussion
Physico-chemical characterisation and binding capacity
Results of elemental analyses are given in Table 1. While the sodium and potassium content of husk was higher than that of seed samples, the nitrogen content of husk was negligible compared to that of seed, indicating no amide groups were likely to be involved in sodium binding in the husk.The binding of sodium to psyllium husk was evaluated in static batches; sodium was ‘lost’ from the psyllium gel after separation, indicating 69% sodium retention (Table 2). In order to identify physical fractions with optimum sodium binding, a range of samples were prepared (seed, gel, husk) with a range of grind sizes (whole seed to finely ground material, <212 μm), their ability to bind to sodium in a batch reactor was evaluated (Table 2).
| Sample | % Sodium retention |
|---|---|
| % Sodium retention is 100× (bound sodium/added sodium), mg g−1 husk ± standard deviation; added sodium was 10 mg g−1 psyllium. | |
| Fraction size | |
| Whole seed | 29 ± 2 |
| 212–250 μm | 51 ± 3 |
| 106–212 μm | 77 ± 0.3 |
| <106 μm | 71 ± 5 |
| Sample preparation | |
| Coarse grind | 55 ± 3 |
| Psyllium gel | 69 ± 1 |
| Husk | 69 ± 0.3 |
Particle sizes fractions of the seed of less than 212 μm had the best retention under the experimental conditions tested. This is to be expected as the smaller particle size fractions would have a greater number of broken cells with a larger proportion of exposed intra-cellular contents ultimately creating a larger surface area to facilitate sodium interaction. Van Craeyveld et al.23 reported the increased solubility and accessibility of arabinoxylan in ball-mill treated husk samples compared with untreated samples. As the retention of sodium in psyllium husk was about the same as the finely ground seed (<216 μm fraction), psyllium husk as received, was used for the rest of the studies (i.e. without further preparation). In the following discussions, psyllium, psyllium gel and husk are used interchangeably to mean samples prepared from psyllium husk.
The influence of pH on the binding of sodium by the psyllium husk
The influence of pH on sodium binding was studied over a range of pH's.2–10 The binding of sodium by psyllium husk varied slightly as a function of pH, as a slightly higher retention was observed under acidic conditions (pH 2) than at near neutral (pH 6 or pH 8) or alkaline pH conditions (pH 10) (Fig. 1). This slight increase in retention at low pH could be due to changes in charge state, modification of hydration shells or morphological changes of the biopolymer at acidic conditions. Indeed, the streaming potential of psyllium husk gel prepared in acid (pH 2) was at least 10 times less negative than those prepared in alkali (pH 11) or without pH adjustment (pH 6.7). Furthermore, charge density of psyllium gels increased as alkalinty increased from acidic gels (pH 2) to neutral (pH 6.7) to alkali (pH 11), indicating pH drives changes to the hydration shell and supports the hypothesis that sodium interaction may occur partially by a charge based mechanism. See ESI† for streaming potential method details.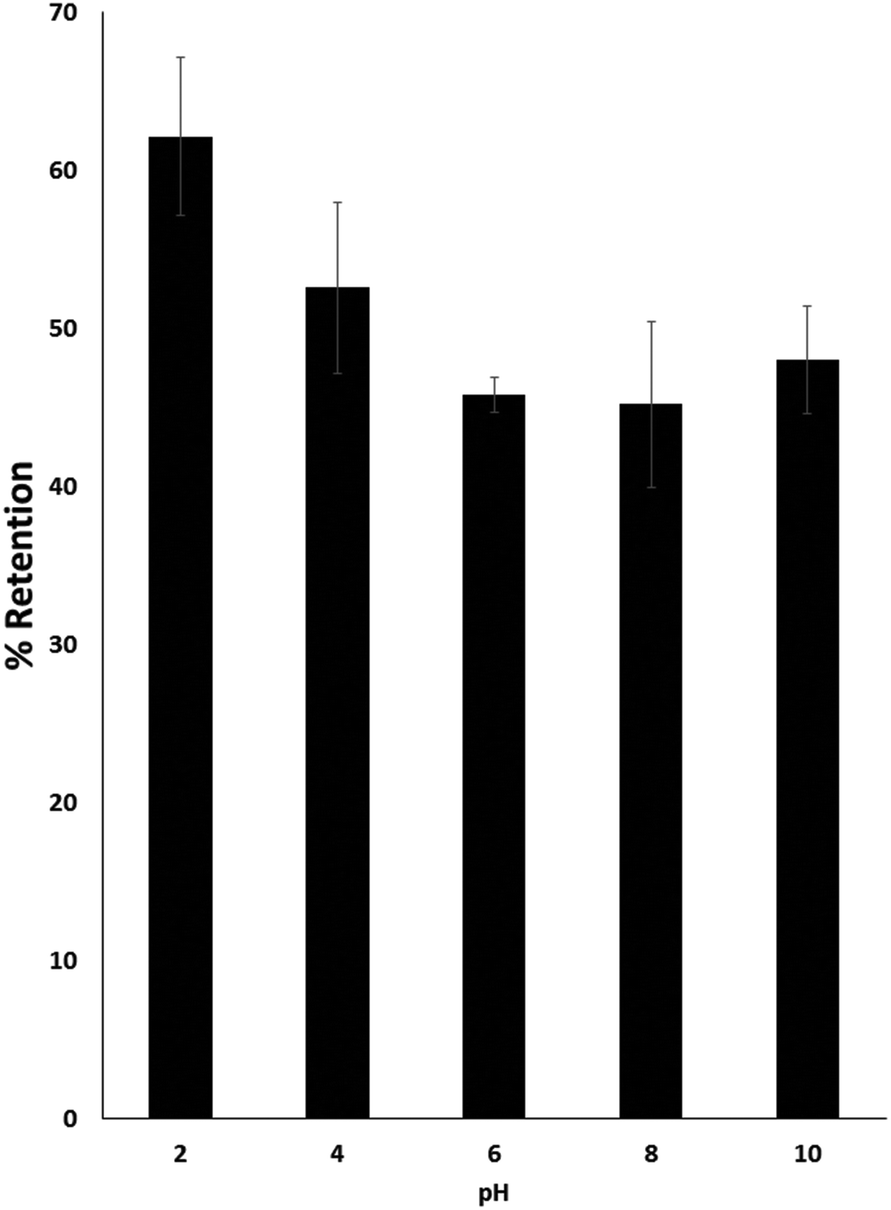 | ||
| Fig. 1 Influence of pH on sodium binding by psyllium husk. Gels were developed by mixing 5 g husk with 5 mL of 0.1% sodium solution, these were then adjusted to the required pH with HCl or KOH solution. | ||
Influence of pH as studied by FTIR spectroscopy
FTIR spectra of psyllium gels prepared in acid (0.1 M HCl) solutions showed distinct differences from those prepared in sodium (1%) solutions in the fingerprint region 1200–700 cm−1, O–H bands about 1630 and 3600–3300 cm−1. Subtraction of the acid spectra from those of alkali gels prepared in 0.1 M KOH solution showed major peak intensity differences at 693, 1049 and 1734 cm−1. The 2nd derivatives further revealed the appearance of a peak at 945 and 892 for the acid gel only in the β-pyranose 980–880 region indicating different interactions of sodium or reduced accessibility to the ring. Slight differences in the form of shifts to higher wavenumbers in 1740–1600 regions in the acid gel compared to the sodium and other gels were also observed. This indicates a strong interaction of acid with the psyllium husk; increased accessibility could explain the slight increase in sodium retention found at lower pH conditions within the batch isotherm experiments. The binding mechanism and possible binding sites were evaluated using FTIR and Na-NMR and are discussed below. Fig. 2 shows the binding behaviour of psyllium husk with different starting concentrations of sodium. Concentrations of sodium below 50 mg g−1 husk followed the Langmuir model (see ESI†), indicating a monolayer adsorption mechanism. At sodium concentrations higher than 100 mg g−1 husk, complex heterogeneous adsorption is assumed to take place through secondary layers. This would explain the high percentage of sodium (at least 50%) retained across the range of concentrations studied.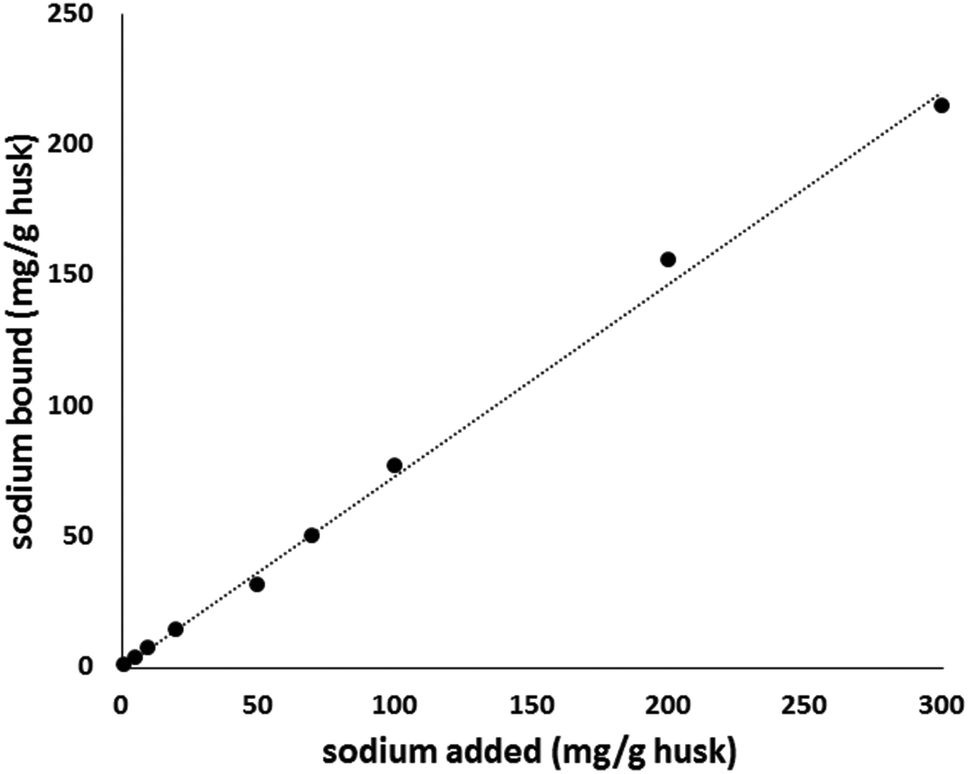 | ||
| Fig. 2 Effect of sodium concentration on sodium binding of psyllium. 500 mg of psyllium husk was mixed with solutions containing increasing concentrations of sodium. | ||
Sodium retention at physiologically relevant pH
Sodium retention at physiologically relevant pH 1.2 and 6.8 was tested in order to assess the retention performance of psyllium in the human gastrointestinal tract. Due to the high sodium content of synthetic gastric juice and buffers available, test solutions containing known amounts of sodium adjusted to relevant pH were used instead. Samples were held at pH values relating to the mouth, stomach and intestine to simulate digestion. It is assumed that most of the sample passes from the mouth to the stomach, at this stage, 47% of the sodium was shown to be retained within the gel (Table 3). If the model is continued further to the intestine, the sodium retention was 47% indicating a significant proportion of sodium is retained and not biologically available. To simulate the effect of sodium reflux into the intestine on the binding capability of psyllium, a further addition of 5 mL of 0.1% w/w sodium test solution was made into the GIT model system (Table 3). The extended GIT model showed that the introduction of a fresh flux of sodium in the intestine did not adversely affect the binding ability as seen in the last step of the simulated GIT model. These results indicate that psyllium could be a potential control mechanism for sodium bioaccessibility.| Model | pH | % Sodium retention |
|---|---|---|
| % Retention is calculated as % bound sodium/added sodium ± sd. | ||
| In-mouth | ∼6.5 | 47 ± 1 |
| Stomach | 1.2 | 45 ± 1 |
| Intestine | 6.8 | 47 ± 1 |
| Sodium addition | ||
| Intestine | 6.8 | 58 ± 6 |
Due to the production of digestive fluids and intake of liquid foods, release of bound sodium under intestinal pH conditions was studied with repeat washings. The results showed that over 80% of the initially bound sodium was retained after 12 repeated washings using a total volume of 120 mL (Fig. 3).
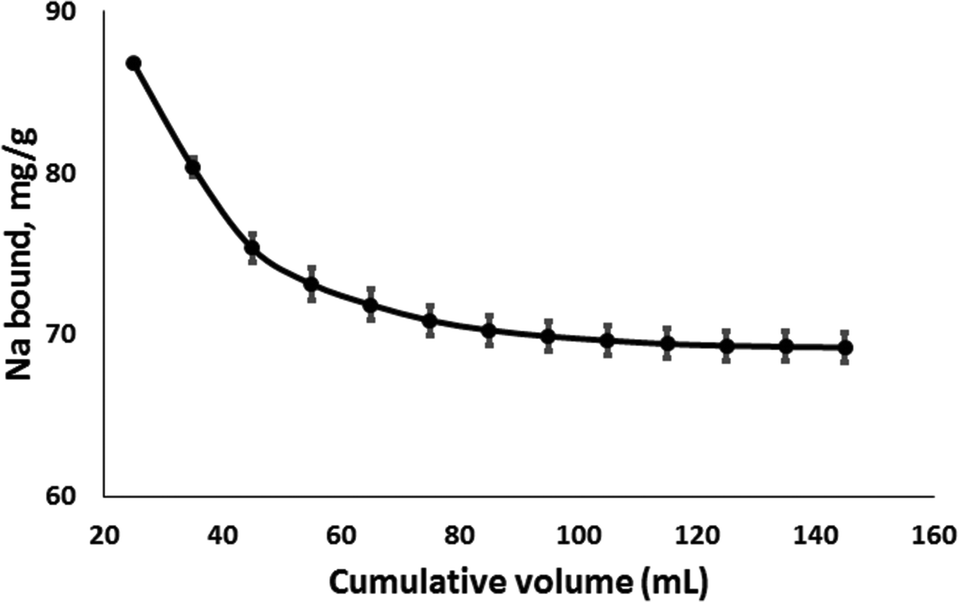 | ||
| Fig. 3 Stability of psyllium bound sodium under repeated washings. Starting concentration was 50 mg Na per 0.5 g husk (washing with 10 mL aliquots of ultrapure water). | ||
Further FTIR study on sodium interaction in psyllium gels
FTIR spectra were generated of psyllium gels prepared under different treatment conditions to propose possible sites of interaction. There were no gross differences between the native gel and the gel with sodium (P > 0.05, Fig. 4). As expected a peak at 1044 cm−1 (C![[double bond, length as m-dash]](https://www.rsc.org/images/entities/char_e001.gif) O) was not present in the deionised water and sodium chloride blank solutions. However, peak shifts in the 3600–3100 cm−1, 1650–1630 cm−1 and 690–600 cm−1 regions were further investigated using subtraction, random forest classification analysis and analysis of 2nd derivatives. Subtraction of different treatments from the sodium gel spectra showed interaction with sodium at 3500–3300, 1080–1040 and the arabinoxylan backbone at 690–600 cm−1 indicating multi-site, non-specific interactions probably with hydroxyl and carboxylic acid groups.
O) was not present in the deionised water and sodium chloride blank solutions. However, peak shifts in the 3600–3100 cm−1, 1650–1630 cm−1 and 690–600 cm−1 regions were further investigated using subtraction, random forest classification analysis and analysis of 2nd derivatives. Subtraction of different treatments from the sodium gel spectra showed interaction with sodium at 3500–3300, 1080–1040 and the arabinoxylan backbone at 690–600 cm−1 indicating multi-site, non-specific interactions probably with hydroxyl and carboxylic acid groups.
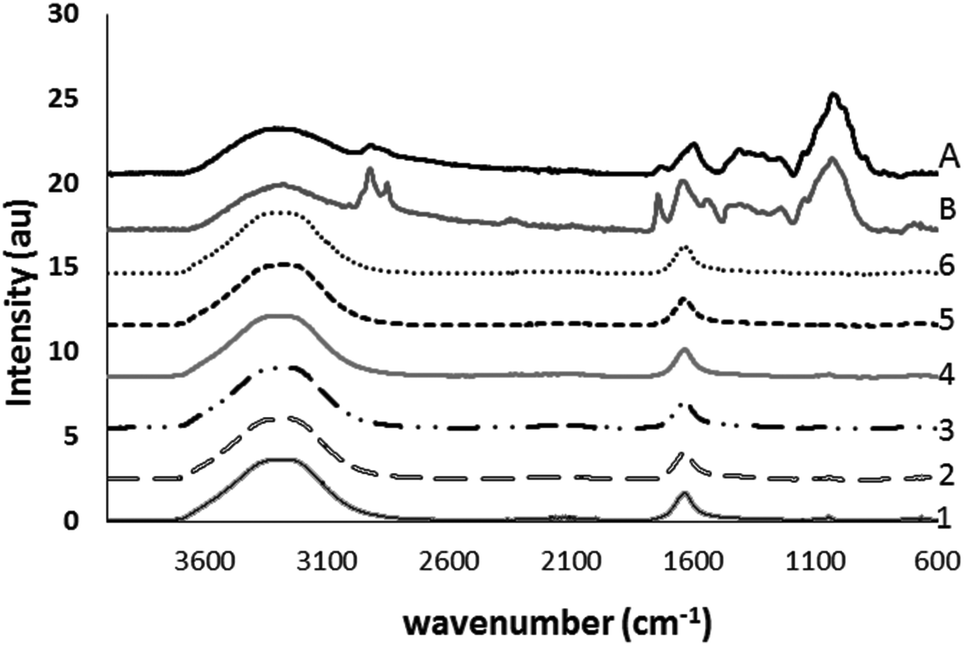 | ||
| Fig. 4 FTIR spectra of psyllium gels prepared under different treatments. 1. Psyllium gel in acid, 2. Psyllium gel in alkali, 3. Psyllium gel in water, 4. Psyllium gel containing sodium, 5. Deionised water and 6. 2% salt solution. Spectra of native husk (A) and seed (B) are added for comparison. | ||
Random forest classification confirmed that wavenumbers ranked in order of significance were the free H-bonding region 3997–3600, OH region of 1750–1600 and the 700–600 cm−1 polymer region.
Spectra presented as 2nd derivative also illustrated changes in the 680–660 cm−1 region which relates to the arabinoxylan backbone, this was similar for the water gel and the calcium containing gel. A shift to lower wavelengths indicated a weakening of bonds or greater interactions and was seen for sodium, acid and alkali treatments in the order sodium ≫ acid > alkali.
It must be borne in mind that the assignments of bands is not absolute and therefore Na-NMR experiments were carried out to further understand the mechanism of sodium binding within the psyllium gel.
23Na-NMR studies on sodium binding mechanism
Two approaches were followed in the 23Na-NMR study. In the 1st approach, free induction decay (FID) data obtained from runs of sodium standards and corresponding psyllium gels containing known but varying amounts of sodium were compared. The difference in measurable sodium can be used to indicate free sodium and bound sodium fractions. The results indicated that approx. 10% sodium is unmeasurable by NMR and is presumed to be strongly interacting (Fig. 5a). Since a higher binding affinity was observed in other experiments, the remaining bound fraction was assumed to be less tightly interacting with the biopolymer matrix, presumably through a hydration shell or steric interaction mechanism. However, we suspect that instrumental limitations (significant dead time of 50 μs for the probe), caused uncertainty in the measurement of the initial amplitude of the signal. Since the decay time for the signal decreases with increasing binding (see ESI Table 1†), the decay time values of the psyllium gel experiments may be biased. This begs the question of how bound is this bound fraction or ‘lost’ to the NMR fraction. The psyllium gel and the salt solution are essentially “liquid like” samples, but can exhibit a range of mobility and can generate different Na-NMR spectra due to the different physical phenomena involved. In the simple FID measurement as in the benchtop compact NMR described in the 1st approach, only the free tumbling ions can be measured.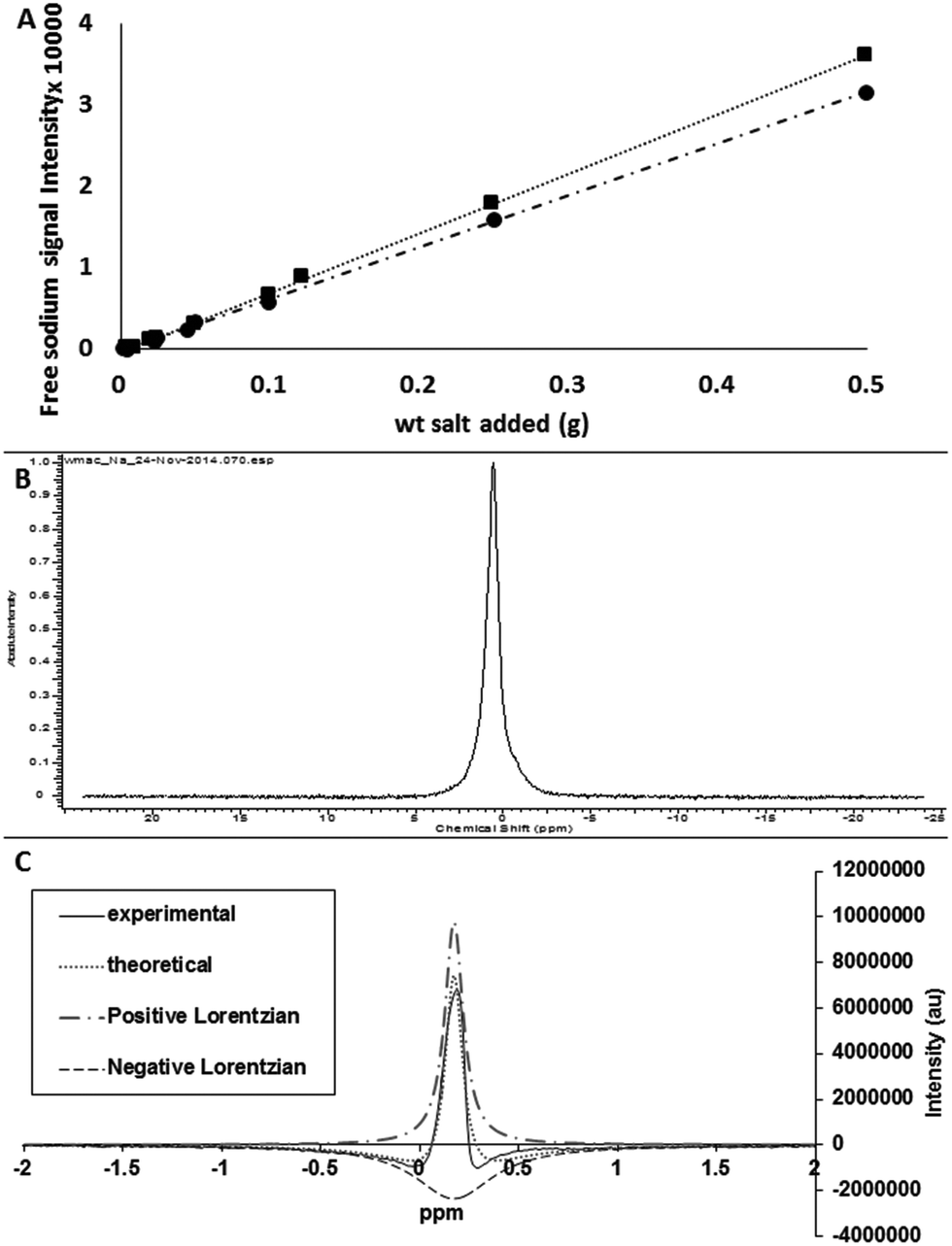 | ||
| Fig. 5 (A) Free sodium chloride detected by compact Na-NMR in standard sodium chloride solutions (square) and the corresponding concentrations of sodium chloride in psyllium gels (circle). Gels were prepared from 500 mg husk in 5 mL of the appropriate concentration of sodium chloride solution (top); (B) single quantum spectrum from psyllium gel prepared in 2% sodium chloride solution (middle); (C) example double quantum filtered signal generated from a carrageenan sample prepared in 2% sodium chloride solution using the same pulse sequence as for psyllium gel samples (no signal), and fit with double Lorentzian functions (bottom). | ||
In the 2nd approach, a high resolution 600 MHz NMR instrument afforded advanced quantum filtration experiments to measure those sodium nuclei which are experiencing a restriction in their motion and so distinguish the sodium ions in different environments(-isotropic/anisotropic). In a normal single quantum experiment, the satellite lines of the spectral triplet are broadened and this produces a double exponential decay in the single quantum FID or equivalently a broadened base in the spectral line (Fig. 5b). When passed through the double quantum filter the result is two antiphase Lorentzian lines as can be seen in the 2% sodium content carrageenan example in Fig. 5c. For psyllium gels with a 0%, 0.5% and 2% sodium content, no DQF signal was observed, indicating no strongly bound fraction of sodium within the gels. This points ultimately towards a non-specific occlusion mechanism.
Conclusions
Sodium was shown to bind to psyllium through a multi-site, non-specific mechanism. A tightly bound fraction is proposed to interact via mono layer adsorption, and a loosely bound fraction by a secondary occlusion process is postulated. The feasibility of psyllium as a scavenger of excess sodium in the diet has also been demonstrated. Psyllium is able to retain sodium across a range of pH, including pH conditions of physiological importance to digestion, and across a wide sodium concentration range. Furthermore binding capacity is not dependent on sodium loading. Psyllium therefore shows promise as a means of reducing the bioavailable fraction of ingested sodium in the body. The biological significance of the findings should be further investigated in-body, in the presence of additional, biologically relevant, counter-ions.Acknowledgements
ERDF, BBSRC (BB/N021126/1, BB/F017014/1) and the Food and Drink INET are acknowledged for funding this research. Dr J. Reps of the Advanced Data Analysis Centre (ADAC) University of Nottingham, provided FTIR data analysis support.References
- L. A. Wyness, J. L. Butriss and S. A. Stanner, Public Health Nutr., 2012, 15, 254–261 CrossRef PubMed.
- F. J. He, H. C. Brinsden and G. A. MacGregor, J. Hum. Hypertens., 2014, 28, 345–352 CAS.
- D. o. Health, New salt targets: F9 Salt Reduction 2017 pledge & F10 Out of Home Salt reduction pledge. https://responsibilitydeal.dh.gov.uk/responsibility-deal-food-network-new-salt-targets-f9-salt-reduction-2017-pledge-f10-out-of-home-salt-reduction-pledge/, accessed 16th May 2014.
- WHO, Global Strategy on Diet, Physical Activity and Health- Population sodium reduction strategies http://www.who.int/dietphysicalactivity/reducingsalt/en, accessed 16th May 2014.
- EUTasteProject, The Application of Edible Seaweed for Taste Enhancement and Salt Replacement http://tasteproject.net, accessed 12th September 2014.
- S. Girgis, B. Neal, J. Prescott, J. Prendergast, L. S. Dumbrel, C. Turner and M. Woodward, Eur. J. Clin. Nutr., 2003, 57, 616–620 CrossRef CAS PubMed.
- C. Morris, A.-L. Koliandris, B. Wolf, J. Hort and A. J. Taylor, Chemosens. Percept., 2009, 2, 1–8 CrossRef.
- R. Rama, N. Chiu, M. Carvalho Da Silva, L. Hewson, J. Hort and I. D. Fisk, J. Texture Stud., 2013, 44, 338–345 CrossRef.
- X. Tian and I. D. Fisk, Food Funct., 2012, 3, 376–380 CAS.
- J. L. Busch, C. Tournier, J. E. Knoop, G. Kooyman and G. Smit, Chem. Senses, 2009, 34, 341–348 CrossRef PubMed.
- M. W. J. Noort, J. H. F. Bult, M. Stieger and R. J. Hamer, J. Cereal Sci., 2010, 52, 378–386 CrossRef CAS.
- S. M. Goh, B. Leroux, C. A. Groeneschild and J. L. Busch, J. Food Sci., 2010, 75, S245–S249 CrossRef CAS PubMed.
- C. G. Biliaderis and M. S. Izydorczyk, Functional food carbohydrates, CRC Press, 1st edn, 2006 Search PubMed.
- B. Singh, Int. J. Pharm., 2007, 334, 1–14 CrossRef CAS PubMed.
- A. Mishra, R. Srinivasan and R. Dubey, Macromol. Mater. Eng., 2002, 287, 592–596 CrossRef CAS.
- J. Sandhu and G. Hudson, Carbohydr. Res., 1981, 247–259 CrossRef CAS.
- M. H. Fischer, N. Yu, G. R. Gray, J. Ralph, L. Anderson and J. A. Marlett, Carbohydr. Res., 2004, 339, 2009–2017 CrossRef CAS PubMed.
- R. Laidlaw and E. Percival, J. Chem. Soc., 1949, 1600–1607 RSC.
- J. Kennedy, J. Sandhu and D. Southgate, Carbohydr. Res., 1979, 75, 265–274 CrossRef CAS.
- Q. Guo, S. W. Cui, Q. Wang and J. Christopher Young, Carbohydr. Polym., 2008, 73, 35–43 CrossRef CAS.
- A. Farahnaky, H. Askari, M. Majzoobi and G. Mesbahi, J. Food Eng., 2010, 100, 294–301 CrossRef CAS.
- R. Kemp-Harper, S. Brown, C. Hughes, P. Styles and S. Wimperis, Prog. Nucl. Magn. Reson. Spectrosc., 1997, 30, 157–181 CrossRef CAS.
- V. Van Craeyveld and J. A. Delcour, J. Agric. Food Chem., 2008, 56, 11306–11311 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/c6fo00785f |
| This journal is © The Royal Society of Chemistry 2016 |