
New structural hypostases of the A·T and G·C Watson–Crick DNA base pairs caused by their mutagenic tautomerisation in a wobble manner: a QM/QTAIM prediction†
Ol’ha O. Brovarets’ab and
Dmytro M. Hovorun*ab
aDepartment of Molecular and Quantum Biophysics, Institute of Molecular Biology and Genetics, National Academy of Sciences of Ukraine, 150 Akademika Zabolotnoho Str., 03680 Kyiv, Ukraine. E-mail: dhovorun@imbg.org.ua
bDepartment of Molecular Biotechnology and Bioinformatics, Institute of High Technologies, Taras Shevchenko National University of Kyiv, 2-h Akademika Hlushkova Ave., 03022 Kyiv, Ukraine
First published on 15th October 2015
Abstract
It is widely known that canonical DNA base pairs preserve the Watson–Crick (WC) scheme of pairing that enables accurate transfer of genetic information across generations. Herein, we firstly present a discovered biologically important ability of the classical A·T(WC) and G·C(WC) nucleobase pairs to tautomerise into the wobble (w) H-bonded mismatches – A*·T↑(w), A·T*O2↑(w), A·T*↓(w) and G·C*↑(w), G*·C↓(w), G·C*↓(w), G*·C↑(w), respectively, containing mutagenic tautomers of the bases (denoted by asterisks), and vice versa. QM and QATIM calculations show that these non-dissociative tautomerisation processes, which are quite fast in comparison with the rate of the DNA replication in the cell, are controlled by the highly stable and highly polar transition states – tight A+·T−, A−·T+, G+·C− and G−·C+ ion pairs, which are formed by the protonated and deprotonated bases. The estimation of the populations of the tautomerised states of the A·T(WC) (6.1 × 10−9 to 1.5 × 10−7) and G·C(WC) (4.2 × 10−11 to 1.4 × 10−9) base pairs in the continuum with ε = 4, corresponding to the interface of the protein–nucleic acid interactions, points to their involvement in the process of the nucleation of the spontaneous point replication errors in DNA, arising with frequencies from around 10−11 to 10−9 errors per nucleotide replicated. The presented concept of the mutagenic quantum wobbling of the canonical A·T(WC) and G·C(WC) nucleobase pairs as an intrinsic property resulting in their tautomerisation allows us to explain the increase in the frequency of point mutations induced by 2-aminopurine, and also to interpret satisfactorily the NMR experimental literature data that show a slow exchange between the G·P(WC) Watson–Crick-like base pair with a corresponding wobble pair, which includes the 6H,8H-3,4-dihydropyrimido[4,5-c][1,2]oxazin-7-one (P) mutagen. These findings significantly expand the horizons of the classical Watson–Crick tautomeric hypothesis of spontaneous point mutagenesis. Collectively, our investigation revealed the hitherto unknown ability of the canonical Watson–Crick DNA base pairs to switch into the wobble mismatches with mutagenic tautomers, clarifying the nature of genome instability. Moreover, this paper reveals new facets of the Watson–Crick tautomeric hypothesis of the spontaneous point mutagenesis arising at DNA replication, and significantly broadens the possibilities for the rational design of chemical mutagens with a targeted action, which could be interesting for medicine, synthetic biology and biotechnology.
Although DNA is replicated in the cell with high precision, errors still occur.1,2 In recent years the Watson–Crick tautomeric hypothesis3 that associates the spontaneous point mutations arising in DNA with the transition of nucleotide bases from the canonical into the rare, mutagenic tautomeric forms4,5 resulting in the formation of the base pairs by their participation, has received reliable experimental evidence,6–10 in particular X-ray analysis,7,8 NMR relaxation dispersion measurements,9,10 and theoretical11–13 verifications. Nevertheless, these data do not allow us to answer on the sore point of whether the physico-chemical mechanisms enable the DNA bases in the canonical tautomeric form to acquire a rare or mutagenic tautomeric form, before the dissociation of the Watson–Crick (WC) nucleobase pairs into the monomers caused by the replication machinery, in order to produce mispairs, resulting in further misincorporations and as a result, spontaneous point mutations during DNA replication. At the same time, establishment of the physico-chemical mechanisms11–16 that ensure the transition of the nucleotide bases from the canonical into the rare tautomeric form during DNA replication, i.e. at the forced dissociation of the Watson–Crick nucleobase pairs into the monomers by the replication machinery, remains a bottleneck.
As of today, it has been definitely established that double proton transfer (DPT) along neighboring intermolecular H-bonds17 both in the Watson–Crick18–21 and wobble22,23 DNA base pairs, as well as in the complexes of amino acid residues of the DNA-binding proteins with nucleotide bases24,25 can’t cope with this biologically important task.
In this paper we have pursued the goal of revealing unknown properties of the well-studied canonical A·T(WC) and G·C(WC) DNA base pairs, which are inherent to these H-bonded complexes and are able to explain the intrinsic DNA mutability without the involvement of external agents.
All calculations of the geometries and harmonic vibrational frequencies of the considered base pairs and transition states of their conversion have been performed using the Gaussian’09 package26 at the DFT (B3LYP)/6-311++G(d,p) level of theory27–29 that has been applied for analogous systems and verified to give accurate geometrical structures, normal mode frequencies, barrier heights and characteristics of intermolecular H-bonds.20,21,30 A scaling factor that is equal to 0.9668 has been applied in the present work for the correction of the harmonic frequencies of all studied base pairs.31–35 We have confirmed the minima and TSs, located by means of the Synchronous Transit-Guided Quasi-Newton method,36 on the potential energy landscape by the absence or presence of the imaginary frequency in the vibrational spectra of the complexes. We applied the standard TS theory37 to estimate the activation barriers of the tautomerisation reactions.
Reaction pathways have been monitored by following the intrinsic reaction coordinate (IRC) in the forward and reverse directions from each TS using the Hessian-based predictor–corrector integration algorithm38 with tight convergence criteria. These calculations eventually ensure that the proper reaction pathway, connecting the expected reactants and products on each side of the TS, has been found. We have investigated the evolution of the energetic and geometric characteristics of the H-bonds and base pairs along the reaction pathway establishing them at each point of the IRC.39–41
In order to consider the electronic correlation effects as accurately as possible, we followed geometry optimizations with single point energy calculations using the MP2 level of theory42 and 6-311++G(2df,pd) Pople’s basis set of valence triple-ζ quality43,44 and aug-cc-pVDZ Dunning’s cc-type basis set,45 augmented with polarization and/or diffuse functions.
Physico-chemical parameters have been estimated by the known formulas of the physico-chemical kinetics37 (for more details see the Methods section in the ESI†). Bader’s quantum theory of atoms in molecules (QTAIM)46,47 was applied to analyse the electron density distribution.48,49
In the present paper we have considered the simplest physico-chemical model of the base mispairs in the base-pair recognition pocket of the high-fidelity DNA-polymerase, namely the H-bonded pairs of nucleotide bases in the continuum with ε = 1/ε = 4.53,70 In this case, we have relied on the results obtained in previous work,50 in which the adequacy of this model was convincingly proved, at least in the study of the tautomerisation of the H-bonded pairs of nucleotide bases, where the insignificance of the influence of the stacking and the sugar-phosphate backbone on the tautomerisation process has been demonstrated. Thereby, their impact can be neglected in the first approximation. In addition, the applied model can help to distinguish the lowest structural level, at which the tautomerisation effects can be observed, and to estimate the changes at the sequential complications of the model.
In this study we have chosen the simplest level of the base pairs that adequately reflects the processes occurring in real systems51 without the deprivation of the structurally functional properties of the bases in the composition of DNA. In this case, the value of the effective dielectric constant ε (1 < ε < 4), that is characteristic for the anhydrous molecular crystals, satisfactorily models the substantially hydrophobic recognition pocket of the DNA-polymerase machinery as a part of the replisome.19,21,25
Mutagenic tautomeric wobbling of the canonical A·T Watson–Crick DNA base pair
In this paper we discovered for the first time a new physico-chemical property of the classical A·T(WC) DNA base pair to be in the fast (in comparison with the rate of the DNA replication in the cell2) tautomeric equilibrium with the short-lived wobble (w) A*·T↑(w), A·T*O2↑(w) and A·T*↓(w) H-bonded mismatches, comprising the mutagenic tautomers of nucleotide bases (here and below marked by an asterisk) (Fig. 1 and 2, Table 1 and Fig. S1, Tables S1–S3†). It turned out that the A·T(WC) DNA base pair exists simultaneously in three other biologically important hypostases – A*·T↑(w), A·T*O2↑(w) and A·T*↓(w). Their forced dissociation into monomers by the DNA-polymerase machinery with the need generates mutagenic tautomers of the DNA bases, which are themselves long-lived structures51 and are able to cause spontaneous point mutations.11–13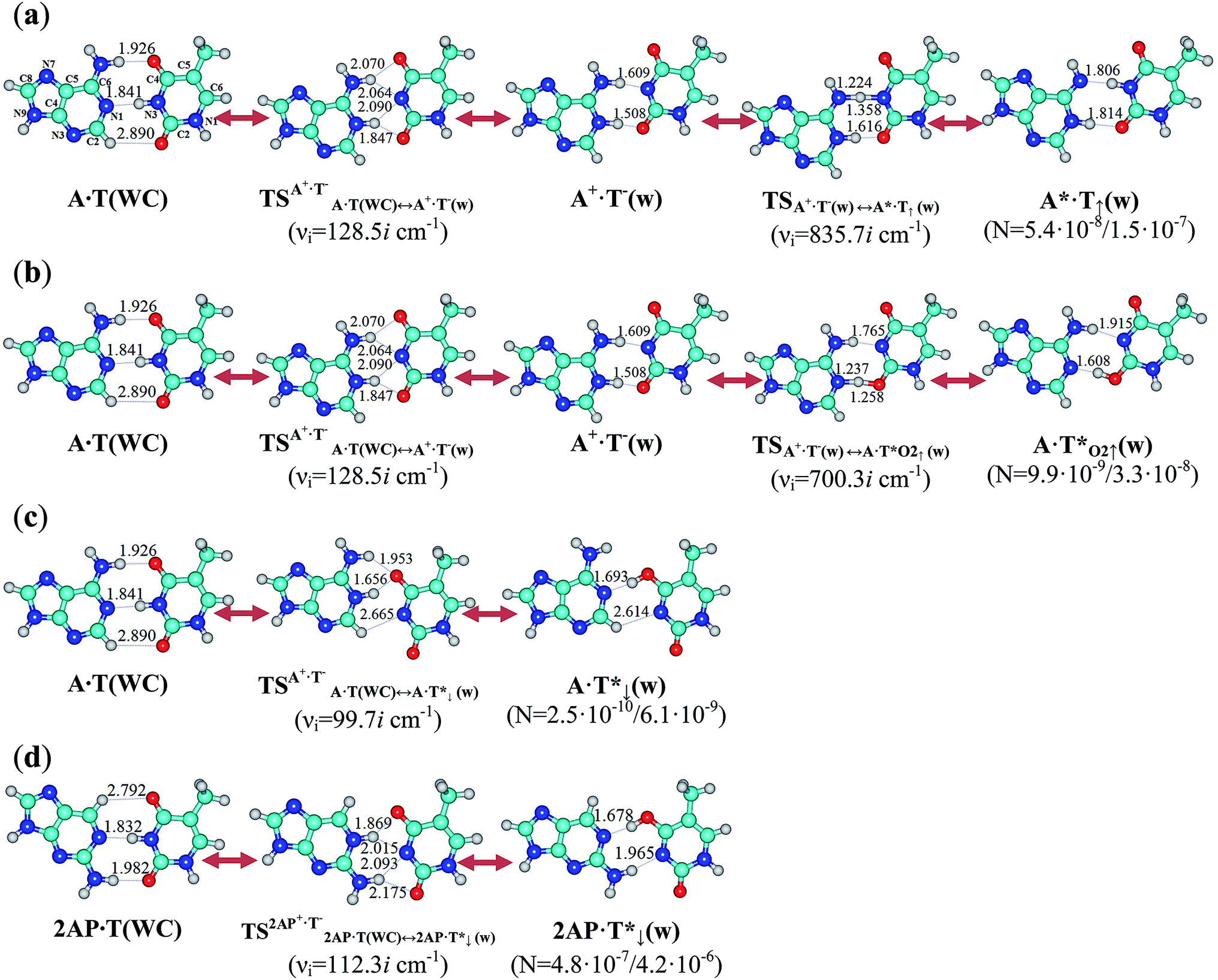 | ||
| Fig. 1 Stationary structures optimized at the B3LYP/6-311++G(d,p) level of theory (ε = 1) on the reaction pathways of the (a) A·T(WC) ↔ A+·T−(w) ↔ A*·T↑(w), (b) A·T(WC) ↔ A+·T−(w) ↔ A·T*O2↑(w), (c) A·T(WC) ↔ A·T*↓(w), and (d) 2AP·T(WC) ↔ 2AP·T*↓(w) tautomerisations. All transition states of these processes represent tight A+·T−/2AP+·T− ion pairs, the geometry of which is no longer Watson–Crick, but is not yet wobble. Populations N of the wobble mispairs containing rare tautomers are presented below them in brackets (MP2/aug-cc-pVDZ//B3LYP/6-311++G(d,p) level of theory in the continuum with ε = 1/ε = 4 at T = 298.15 K). The dotted lines indicate AH⋯B H-bonds and continuous lines indicate loosened A–H–B covalent bridges (their lengths in the continuum with ε = 1 are presented in angstroms; for more detailed physico-chemical characteristics of the H-bonds see Table S1†); carbon atoms are in light-blue, nitrogen atoms in dark-blue, hydrogen atoms in grey and oxygen atoms in red; νi is the imaginary frequency; the numeration of atoms depicted on the A·T(WC) base pair is standard. | ||
 | ||
| Fig. 2 Energetic profiles of the mutagenic tautomeric wobbling of the A·T(WC) and 2AP·T(WC) DNA base pairs due to their tautomeric transformations via the (a) A·T(WC) ↔ A+·T−(w) ↔ A*·T↑(w), A·T(WC) ↔ A+·T−(w) ↔ A·T*O2↑(w), A·T(WC) ↔ A·T*↓(w) and (b) 2AP·T(WC) ↔ 2AP·T*↓(w) tautomerisation pathways, depicted with key stationary structures on the potential energy hypersurface (see also Fig. 1). These tautomeric conversions occur through the initial nipping off of the migrating proton from the T base and joining it to the A/2AP complementary base, then the shifting and significant rebuilding of the bases relative to each other within the base pair into the major or minor groove sides of the DNA helix, and the further addition of the mobile or other acidic proton to the neighboring N3 nitrogen or O2/O4 oxygen atoms of the T base. The A·T(WC) ↔ A*·T↑(w) and A·T(WC) ↔ A·T*O2↑(w) tautomerisation reactions proceed via the dynamically unstable intermediate tight A+·T−(w) ion pair with wobble architecture, exactly at which bifurcation occurs into two different pathways. All transition states of these processes represent itself tight A+·T−/2AP+·T− ion pairs, the geometry of which is no longer Watson–Crick, but is not yet wobble. These data are obtained by following the IRC at the B3LYP/6-311++G(d,p) level of theory in the continuum with ε = 1. For the detailed designations of the intermolecular bonds, atoms, their numeration and complex populations see Fig. 1 caption. | ||
| Tautomerisation | εa | ΔGb | ΔEc | ΔΔGTSd | ΔΔETSe | ΔΔGf | ΔΔEg | τ99.9%h | τi | Nj |
|---|---|---|---|---|---|---|---|---|---|---|
| a The dielectric constant.b The Gibbs free energy of the product relative to the reactant of the tautomerisation reaction (T = 298.15 K), kcal mol−1.c The electronic energy of the product relative to the reactant of the tautomerisation reaction, kcal mol−1.d The Gibbs free energy barrier for the forward reaction of tautomerisation kcal mol−1.e The electronic energy barrier for the forward reaction of tautomerisation kcal mol−1.f The Gibbs free energy barrier for the reverse reaction of tautomerisation kcal mol−1.g The electronic energy barrier for the reverse reaction of tautomerisation kcal mol−1.h The time necessary to reach 99.9% of the equilibrium concentration between the reactant and the product of the tautomerisation reaction, s.i The lifetime of the product of the tautomerisation reaction, s.j Population of the wobble mispairs containing mutagenic tautomers. See also summary Table S2 for the Gibbs and electronic energies of the mispairs and TSs relatively the global minimum – the A·T(WC) DNA base pair. | ||||||||||
| A·T(WC) ↔ A*·T↑(w) | 1 | 9.90 | 9.59 | 16.72 | 16.02 | 6.82 | 6.43 | 1.10 × 10−7 | 1.59 × 10−8 | 5.4 × 10−8 |
| 4 | 9.32 | 8.78 | 14.41 | 13.56 | 5.09 | 4.78 | 9.59 × 10−9 | 8.62 × 10−10 | 1.5 × 10−7 | |
| A·T(WC) ↔ A·T*O2↑(w) | 1 | 10.91 | 11.25 | 16.72 | 16.02 | 5.81 | 4.77 | 2.02 × 10−8 | 2.92 × 10−9 | 9.9 × 10−9 |
| 4 | 10.19 | 10.48 | 14.41 | 13.56 | 4.22 | 3.07 | 1.36 × 10−9 | 1.97 × 10−10 | 3.3 × 10−8 | |
| A·T(WC) ↔ A·T*↓(w) | 1 | 13.08 | 14.84 | 20.28 | 20.41 | 7.20 | 5.57 | 2.09 × 10−7 | 3.03 × 10−8 | 2.5 × 10−10 |
| 4 | 10.69 | 12.47 | 16.03 | 16.42 | 5.34 | 3.94 | 9.20 × 10−9 | 1.33 × 10−9 | 6.1 × 10−9 | |
| 2AP·T(WC) ↔ 2AP·T*↓(w) | 1 | 8.62 | 8.33 | 18.66 | 17.53 | 10.04 | 9.21 | 2.53 × 10−5 | 3.66 × 10−6 | 4.8 × 10−7 |
| 4 | 7.33 | 7.79 | 15.48 | 14.72 | 8.15 | 6.93 | 1.04 × 10−6 | 1.50 × 10−7 | 4.2 × 10−6 | |
A common feature of all three A·T(WC) ↔ A*·T↑(w), A·T(WC) ↔ A·T*O2↑(w) and A·T(WC) ↔ A·T*↓(w) routes of the quantum mutagenic tautomerisation via the sequential DPT accompanied with the structural rearrangement of the bases lies in the fact that they are controlled by the plane symmetrical, highly stable and highly polar transition states – tight ion pairs, formed by the protonated A+ base and deprotonated T− base – and occur by the non-dissociative mechanism (Fig. 1, 2 and S1†). This non-dissociative quantum mechanism of the mutual transformation of the base pairs is accompanied by the significant alteration of their geometry (from Watson–Crick to wobble and vice versa) and succession of the patterns (from 9 to 10) of specific intermolecular interactions including both AH⋯B H-bonds and loosened A–H–B covalent bridges along the IRC of tautomerisation (Fig. 1–3 and Table S5†). A quite high stabilization energy (>100 kcal mol−1) of the transition states of the tautomerisation processes (Table S3†) negates the direct participation of water molecules in them as a chemical agent, and eliminates the influence of the stacking interactions52 on the course of the tautomerisation. This point of view11–13 is confirmed experimentally for the similar G·T(w) ↔ G*·T(WC) tautomerisation.9 The Cs symmetry of the structures involved in all three tautomerisation processes remains unchanged; at this, the orientation of the methyl group of the T base also remains undisturbed. Notably, tautomerised A*·T↑(w), A·T*O2↑(w) and A·T*↓(w) mismatches and transition states of their tautomerisation – A·T(WC) ↔ A*·T↑(w), A·T(WC) ↔ A·T*O2↑(w) and A·T(WC) ↔ A·T*↓(w) – have considerably more polar structure than the A·T(WC) DNA base pair (Table S1†). These structures, that are involved in the above described processes of tautomerisation, are thermodynamically stable structures with a negative value of Gibbs free energy of interaction (ΔGint < 0) (Table S3†).
 | ||
| Fig. 3 Exchange patterns of the intermolecular AH⋯B H-bonds along the IRC of the biologically important tautomerisations of the A·T(WC) DNA base pair via the sequential DPT, accompanied by the structural rearrangement of the bases: (a) A·T(WC) ↔ A*·T↑(w), (b) A·T(WC) ↔ A·T*O2↑(w) and (c) A·T(WC) ↔ A·T*↓(w) (B3LYP/6-311++G(d,p) level of theory (ε = 1)). Here and below, the perpendicular lines denote the partition of the range into different regions. Detailed numerical characteristics of the unique patterns of the specific intermolecular interactions, the sequential change of which accompanies the non-dissociative processes of tautomerisation, are presented in Table S5.† | ||
The A·T(WC) ↔ A·T*↓(w) tautomerisation process, where a two-stage transition of the same proton occurs, is much easier than the two others, A·T(WC) ↔ A*·T↑(w) and A·T(WC) ↔ A·T*O2↑(w) (Fig. 1 and 2). These two last processes take place through a common 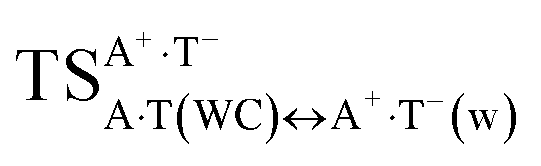 transition state and a dynamically unstable intermediate – the H-bonded tight A+·T−(w) ion pair with wobble architecture, and exactly at this point, a bifurcation of the tautomerisation into two different (in topological and energetic aspects) pathways occurs, the final products of which are A*·T↑(w) and A·T*O2↑(w) mispairs.
transition state and a dynamically unstable intermediate – the H-bonded tight A+·T−(w) ion pair with wobble architecture, and exactly at this point, a bifurcation of the tautomerisation into two different (in topological and energetic aspects) pathways occurs, the final products of which are A*·T↑(w) and A·T*O2↑(w) mispairs. 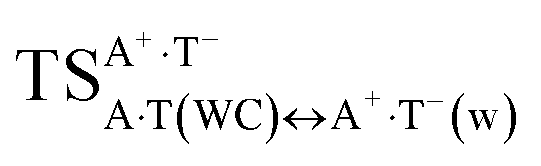 is in fact the transition state for both A·T(WC) ↔ A*·T↑(w) and A·T(WC) ↔ A·T*O2↑(w) tautomerisation reactions (Fig. 1 and 2). It was established at the MP2/aug-cc-pVDZ//B3LYP/6-311++G(d,p) level of theory that the A+·T−(w) intermediate is absent on the potential energy surface, since the A+·T−(w) ↔ A·T*O2↑(w) and A+·T−(w) ↔ A*·T↑(w) tautomeric conversions are barrier-less in the continuum with ε = 1/ε = 4 (see Table S4†).
is in fact the transition state for both A·T(WC) ↔ A*·T↑(w) and A·T(WC) ↔ A·T*O2↑(w) tautomerisation reactions (Fig. 1 and 2). It was established at the MP2/aug-cc-pVDZ//B3LYP/6-311++G(d,p) level of theory that the A+·T−(w) intermediate is absent on the potential energy surface, since the A+·T−(w) ↔ A·T*O2↑(w) and A+·T−(w) ↔ A*·T↑(w) tautomeric conversions are barrier-less in the continuum with ε = 1/ε = 4 (see Table S4†).
Comparison of the energy barriers of the mutagenic tautomerisation of the isolated A and T bases (45.7 and 39.2 kcal mol−1, respectively)51 with similar values in the composition of the pair (17.4 and 20.8 kcal mol−1, respectively (Table S4†)) clearly provides evidence that complementary H-bonded bases catalyze the tautomeric transformation of each other into the mutagenic tautomeric forms.
On the one hand, the population of the tautomerised states of the A·T(WC) DNA base pairs A*·T↑(w), A·T*O2↑(w) and A·T*↓(w) equals to 5.4 × 10−8/1.5 × 10−7, 9.9 × 10−9/3.3 × 10−8 and 2.5 × 10−10/6.1 × 10−9, respectively, as obtained at ε = 1/ε = 4 (a continuum with a low dielectric constant is typical for the hydrophobic interfaces of specific protein–nucleic acid interactions25,53) (Fig. 1), which fits into the range of the frequencies of the spontaneous point mutations observed experimentally (10−11 to 10−9).54–56
On the other hand, the wobble A*·T↑(w), A·T*O2↑(w) and A·T*↓(w) H-bonded mispairs satisfy all the necessary conditions in order to be successfully dissociated into the monomers by the replication machinery.19–23 Indeed, they incorporate well into the structure of the DNA double helix,57,58 the interaction energy between the bases in them does not exceed a similar value for the G·C(WC) DNA base pair (ΔEint = −29.28 and ΔGint = −15.97 kcal mol−1),21 and they have a fairly long lifetimes (τ = 1.6 × 10−8, 2.9 × 10−9 and 3.0 × 10−8 s, respectively) (Table 1†). This indicates that we have uncovered for the first time that it is an inherent property of the A·T(WC) DNA base pair to exist in a rapid tautomeric equilibrium with the A*·T↑(w), A·T*O2↑(w) and A·T*↓(w) mispairs (time τ99.9% necessary to reach 99.9% of the equilibrium concentration between the reactant and the product of the tautomerisation reactions is equal to 1.1 × 10−7/6.0 × 10−9, 2.0 × 10−8/1.4 × 10−9 and 2.1 × 10−7/9.2 × 10−9 s (ε = 1/ε = 4), respectively (Table S4†)), which has direct relevance to the occurrence of the spontaneous point replication errors in DNA.
The low population of the tautomerised states of the classical A·T(WC) DNA base pair complicates its direct observation by experimental physico-chemical methods. Meanwhile, the concepts developed in this work about the spontaneous mutagenic tautomerisation of the A·T(WC) base pair allow us to explain quite naturally the microstructural mechanism of the mutagenic pressure exerted by 2-aminopurine (2AP) on DNA. It is well known59–63 that this strong mutagen causes the A·T → G·C spontaneous point replication error in the DNA. According to our data, the tautomerisation reaction of the 2AP·T(WC)64 base pair into the 2AP·T*↓(w) mispair containing the T* mutagenic tautomer and back (Fig. 1 and 2) causes the increasing of the population of the latter in comparison with the A·T*↓(w) mispair by 1887/219 times under normal conditions in the continuum with ε = 1/ε = 4 (accordingly, the probability of the G·T*(WC) mismatch65 formation in the next cycle of the DNA replication increases in the appropriate number of times) (Fig. 1 and 2, Tables 1 and S1–S4†). One of the basic reasons for this is that the weak C2H⋯N3 H-bond (1.41 kcal mol−1) in the A·T*↓(w) mispair is replaced by a much stronger classical N2H⋯N3 H-bond (5.27 kcal mol−1) in the 2AP·T*↓(w) mismatch (Fig. 1 and 2, Tables 1 and S1†). In our opinion, the outlined above considerations are an important confirmation of the adequacy of our model of the tautomeric non-stability of the A·T(WC) DNA base pair.
Finally, we would like to note that in addition to the pathways analyzed in detail above, we have also detected two other A·T*↑(w) ↔ A·T(WC) ↔ A*·T↓(w) tautomerisation routes, controlled by the transition states that are H-bonded tight A−·T+ ion pairs (Fig. 4, S2, S3 and Tables S1–S4†). In view of their extreme slowness (τ99.9% = 6.6 × 1010 and 9.1 × 109 s, respectively (Table S4†), that by orders of magnitude exceeds the time of DNA replication in the cell2), these tautomerisation processes do not present any biological interest and can be attractive only from a theoretical point of view.
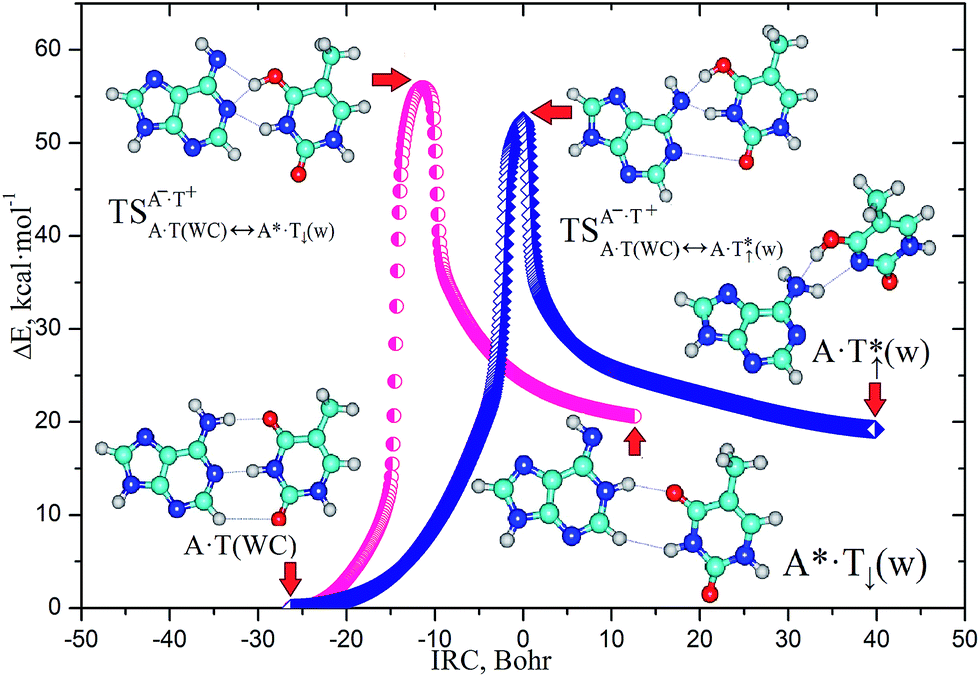 | ||
| Fig. 4 Energetic profiles combined with the stationary points of the high-energy A·T(WC) ↔ A·T*↑(w) and A·T(WC) ↔ A*·T↓(w) reaction pathways of the mutagenic tautomeric variability of the A·T(WC) DNA base pair, proceeding via the initial nipping off of the migrating proton from the A base and joining it to the T complementary base, accompanied with structural rearrangements of the bases relative to each other within the base pair into the major or minor groove sides of the DNA helix, and the terminal transfer of the mobile or other acidic proton to the N1/N6 nitrogen atoms of the A base (see also Fig. S2, S3 and Table S1–S4†). These data are obtained by following IRC at the B3LYP/6-311++G(d,p) level of theory in the continuum with ε = 1. For the detailed designations of the intermolecular bonds, atoms and their numeration see Fig. 1 caption. | ||
Mutagenic tautomeric wobbling of the canonical G·C Watson–Crick DNA base pair
We have managed to pioneeringly establish that the classical G·C(WC) DNA base pair stays in the rapid (in comparison with the rate of the DNA replication in the cell2) tautomeric equilibrium with the four wobble G·C*↑(w), G*·C↓(w), G·C*↓(w) and G*·C↑(w) pairs involving mutagenic tautomers of the DNA bases (Fig. 5, 7, Table 2 and Fig. S4, Table S6–S10†). Our QM computational experiment demonstrates that the forced dissociation of these mispairs into monomers by the DNA-polymerase machinery would generate mutagenic tautomers of the DNA bases that are able to cause spontaneous point mutations – replication errors.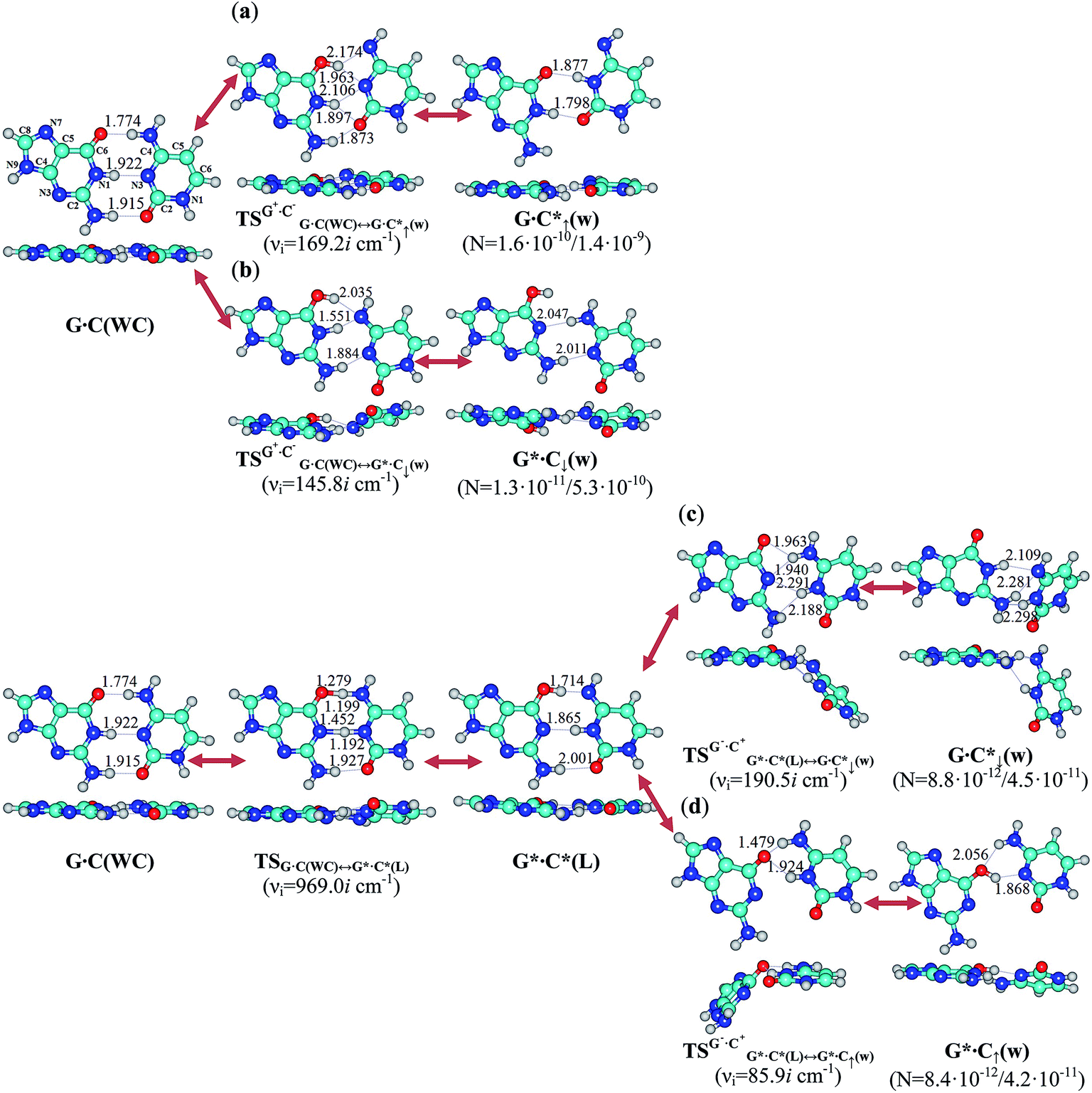 | ||
| Fig. 5 Stationary structures determined on the potential energy hypersurface of the canonical G·C(WC) DNA base pair and the pathways of their tautomeric interconversions: calculations at the B3LYP/6-311++G(d,p) level of theory (ε = 1). The (a) G·C(WC) ↔ G·C*↑(w), (b) G·C(WC) ↔ G*·C↓(w), (c) G·C(WC) ↔ G*·C*(L) ↔ G·C*↓(w) and (d) G·C(WC) ↔ G*·C*(L) ↔ G*·C↑(w) tautomerisations occur through the initial nipping off of the migrating proton from the G or C base, then shifting and significant rebuilding of the bases relative to each other within the base pair into the major or minor groove sides of the DNA helix, and the further addition of the mobile or other acidic proton to the neighboring nitrogen or oxygen atoms of the complementary base. All transition states of these processes represent tight G+·C− or G−·C+ ion pairs, the geometry of which is no longer Watson–Crick, but is not yet wobble. Perpendicular projections of all complexes are given below them. For the detailed designations of the intermolecular bonds, atoms, their numeration and complex populations see Fig. 1 caption. | ||
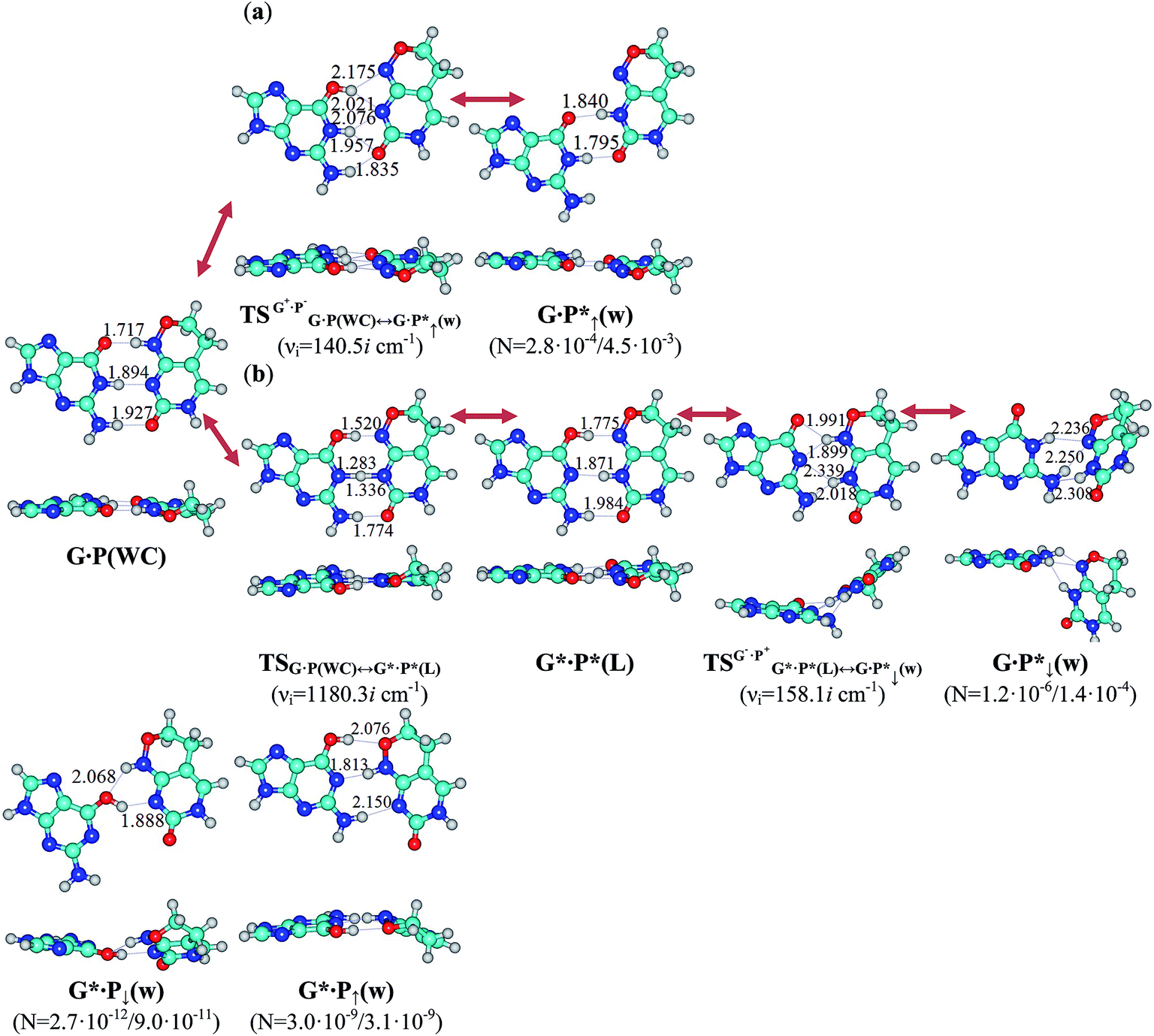 | ||
| Fig. 6 Stationary structures determined on the potential energy hypersurface of the G·P(WC) base pair involving the P cytosine analogue and their tautomerisation pathways into the mispairs containing the mutagenic tautomer of P: calculations at the B3LYP/6-311++G(d,p) level of theory (ε = 1). The (a) G·P(WC) ↔ G·P*↑(w) and (b) G·P(WC) ↔ G·P*↓(w) tautomerisations occur through the initial nipping off of the migrating proton from the G or P base, then shifting and significant rebuilding of the bases relative to each other within the base pair into the major or minor groove sides of the DNA helix, and further addition of the mobile or other acidic proton to the neighboring nitrogen or oxygen atoms of the complementary base. All transition states of these processes represent tight G+·P− or G−·P+ ion pairs, the geometry of which is no longer Watson–Crick, but is not yet wobble. Perpendicular projections of all complexes are given below them. For the designation of the intermolecular bonds, atoms, their numeration and complex populations see Fig. 1 caption. | ||
 | ||
| Fig. 7 Diversity of the reaction pathways of the (a) G·C(WC) ↔ G·C*↑(w), G·C(WC) ↔ G*·C↓(w), G·C(WC) ↔ G·C*↓(w), G·C(WC) ↔ G*·C↑(w) and (b) G·P(WC) ↔ G·P*↑(w), G·P(WC) ↔ G·P*↓(w) mutagenic wobbling tautomerisations depicted together with key stationary structures on the potential energy hypersurface (see also Fig. 5 and 6). The G·C(WC) ↔ G·C*↓(w), G·C(WC) ↔ G*·C↑(w) and G·P(WC) ↔ G·P*↓(w) processes proceed via short-lived and dynamically stable intermediates – the G*·C*(L) and G*·P*(L) Löwdin’s base mispairs with Watson–Crick geometry. The presented data are obtained by following IRC at the B3LYP/6-311++G(d,p) level of theory in the continuum with ε = 1. Various routes of the tautomerisation reactions are depicted in different colors. The peaks of the curves correspond to the transition states of the tautomerisation processes, while the minima correspond to the base pairs with Watson–Crick or wobble architectures. For more detailed physico-chemical characteristics of these tautomeric transformations see Table 2 and S6–S14.† | ||
| Tautomerisation | ε | ΔG | ΔE | ΔΔGTS | ΔΔETS | τ99.9% | N |
|---|---|---|---|---|---|---|---|
| a For the detailed designations see Table 1. See also summary Table S7 for the Gibbs and electronic energies of the mispairs and TSs relative to the global minimum – the G·C(WC) DNA base pair. | |||||||
| G·C(WC) ↔ G·C*↑(w) | 1 | 13.35 | 14.10 | 30.47 | 30.74 | 3.95 | 1.6 × 10−10 |
| 4 | 12.08 | 12.55 | 31.35 | 31.73 | 148.66 | 1.4 × 10−9 | |
| G·C(WC) ↔ G*·C↓(w) | 1 | 15.10 | 16.49 | 31.08 | 31.53 | 0.58 | 1.3 × 10−11 |
| 4 | 14.14 | 15.40 | 32.50 | 32.40 | 32.45 | 5.3 × 10−10 | |
| G·C(WC) ↔ G·C*↓(w) | 1 | 15.08 | 15.96 | 30.88 | 31.41 | 0.42 | 8.8 × 10−12 |
| 4 | 14.11 | 15.48 | 24.31 | 24.59 | 3.35 × 10−5 | 4.5 × 10−11 | |
| G·C(WC) ↔ G*·C↑(w) | 1 | 14.85 | 17.22 | 24.87 | 25.64 | 2.50 × 10−5 | 8.4 × 10−12 |
| 4 | 12.64 | 14.59 | 19.30 | 19.06 | 8.45 × 10−8 | 4.2 × 10−11 | |
| G·P(WC) ↔ G·P*↑(w) | 1 | 4.85 | 5.97 | 21.77 | 22.08 | 2.80 | 2.8 × 10−4 |
| 4 | 3.20 | 4.01 | 21.99 | 22.30 | 65.37 | 4.5 × 10−3 | |
| G·P(WC) ↔ G·P*↓(w) | 1 | 8.08 | 9.42 | 31.44 | 31.83 | 1.48 × 105 | 1.2 × 10−6 |
| 4 | 5.28 | 6.89 | 24.43 | 24.67 | 123.32 | 1.4 × 10−4 | |
It should be noted that all four G·C(WC) ↔ G·C*↑(w), G·C(WC) ↔ G*·C↓(w), G·C(WC) ↔ G·C*↓(w) and G·C(WC) ↔ G*·C↑(w) pathways of the mutagenic tautomerisation occur non-dissociatively (Fig. 8, S9 and Table S14†) via the highly stable and highly polar transition states – tight G+·C− and G−·C+ ion pairs – formed by the protonated and deprotonated bases (Fig. 5 and 7). Bases in the aforementioned complexes, which are involved in these tautomerisation processes, are tightly bound in view of the negative values of the Gibbs free energy of interaction (ΔGint < 0) (Table S9†), extremely high values of which (>100 kcal mol−1) are achieved at the TSs. This means that the impact of the water molecules and stacking interactions52,66,67 on the course of the considered processes is negligible.
 | ||
| Fig. 8 Exchange of the patterns of the intermolecular AH⋯B, NH⋯HN H-bonds and attractive N⋯O/N van der Waals contacts along the IRC of the biologically important tautomerisations of the G·C(WC) DNA base pair via the sequential DPT accompanied with structural rearrangements of the bases relative to each other: (a) G·C(WC) ↔ G·C*↑(w), (b) G·C(WC) ↔ G*·C↓(w), (c) G·C(WC) ↔ G*·C*(L) ↔ G·C*↓(w) and (d) G·C(WC) ↔ G*·C*(L) ↔ G*·C↑(w) (B3LYP/6-311++G(d,p) level of theory (ε = 1)). Detailed numerical characteristics of the unique patterns of the specific intermolecular interactions, in which sequential changing accompanies the non-dissociative processes of tautomerisation, are presented in Table S14.† | ||
This non-dissociative quantum mechanism of the sequential DPT in the nucleobase pairs is accompanied by their substantial rearrangement and sequential change of their intermolecular specific interactions (from 10 to 15 patterns), in particular AH⋯B, NH⋯HN H-bonds, attractive N⋯O/N van der Waals contacts and loosened A–H–B covalent bridges along the IRC of tautomerisation (Fig. 8 and Table S14†).
This approach12–16 has stood the test of time for other mispairs and was confirmed experimentally for the G·T(w)9,10 and G·P(WC)68,69 base mispairs.
It becomes clear from the comparison of the energy barriers of the mutagenic tautomerisation of the G and C bases in the isolated state51 with the corresponding values in the composition of the G·C(WC) DNA base pair (Tables 2 and S10†), that complementary H-bonded bases catalyze the tautomeric transformation of each other.
The G·C(WC) ↔ G·C*↑(w) and G·C(WC) ↔ G*·C↓(w) tautomerisation processes occurring without the involvement of the intermediates are significantly simpler than two others, G·C(WC) ↔ G·C*↓(w) and G·C(WC) ↔ G*·C↑(w) (Fig. 5 and 7, Tables 2 and S10†). These last two processes proceed via the dynamically stable intermediate – the G*·C*(L) Löwdin’s (L) base mispair with Watson–Crick geometry21 (Fig. 5 and 7, Tables 2 and S10†).
It should be emphasized that all four wobble G·C*↑(w), G*·C↓(w), G·C*↓(w) and G*·C↑(w) pairs do not tautomerise through the DPT along the neighboring intermolecular H-bonds, because the products of these reactions are dynamically unstable and short-lived structures, for which low-frequency intermolecular vibrations can’t develop during their lifetimes (Fig. S5, S6 and Table S12†). It also draws attention to the significant non-planarity of the 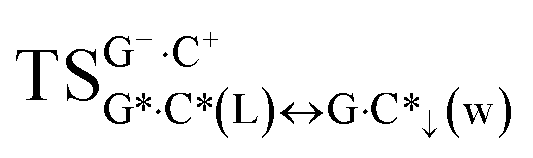 (∠C6N1(G+)N3C4(C−) = ±38.0°) and
(∠C6N1(G+)N3C4(C−) = ±38.0°) and  (∠C6N1(G−)N3C4(C+) = ±40.2°) transition states and also of the wobble G·C*↓(w) (∠N1C2(G)N3C4(C*) = ±42.3°) and G*·C↑(w) (∠C6N1(G*)N1C6(C) = ±31.0°) mispairs (Fig. S7, S8 and Table S13†). However, our simulation results show that it is not an obstacle for the incorporation of these complexes into the structure of the DNA double helix.
(∠C6N1(G−)N3C4(C+) = ±40.2°) transition states and also of the wobble G·C*↓(w) (∠N1C2(G)N3C4(C*) = ±42.3°) and G*·C↑(w) (∠C6N1(G*)N1C6(C) = ±31.0°) mispairs (Fig. S7, S8 and Table S13†). However, our simulation results show that it is not an obstacle for the incorporation of these complexes into the structure of the DNA double helix.
Detailed physico-chemical analysis revealed that the wobble G·C*↑(w), G*·C↓(w), G·C*↓(w) and G*·C↑(w) mismatches can be successfully dissociated into the monomers by the replication machinery, since they fulfill all the conditions which are necessary to accomplish this purpose.19,21,23 Thus, these tautomerised mispairs can be embedded well57,58 into the structure of the DNA double helix owing to the fact that their interaction energy between the bases is lower than a similar value for the canonical G·C(WC) DNA base pair;21 they have rather long lifetimes (21.5, 4.7, 4.9 × 10−6 and 1.2 × 10−8 s, respectively) and, in addition, the population of these tautomerised states of the canonical G·C(WC) DNA base pair – G·C*↑(w), G*·C↓(w), G·C*↓(w) and G*·C↑(w) – is 1.6 × 10−10/1.4 × 10−9, 1.3 × 10−11/5.3 × 10−10, 8.8 × 10−12/4.5 × 10−11 and 8.4 × 10−12/4.2 × 10−11, respectively, in the continuum with ε = 1/ε = 4 53,70 spanning the experimentally observed range of the frequencies of the spontaneous point mutations from 10−11 to 10−9.54–56 All these facts gathered together indicate that the intrinsic property, identified here for the first time, of the G·C(WC) DNA base pair to stay in rapid tautomeric equilibrium with the wobble G·C*↑(w), G*·C↓(w), G·C*↓(w) and G*·C↑(w) pairs is closely related to the occurrence of spontaneous point mutations – DNA replication errors.
The low population of the tautomerised states of the classical G·C(WC) base pair complicates their observation by the experimental physico-chemical methods. Meanwhile, using NMR spectroscopy the conversion of the G·P(WC) base pair including 6H,8H-3,4-dihydropyrimido[4,5-c][1,2]oxazin-7-one (P) mutagen6 into the wobble configuration and vice versa was observed in the DNA composition.68,69 In this case, according to the presented above model, the population of the tautomerised G·P*↑(w) and G·P*↓(w) states increased by orders of magnitude in comparison with similar G·C*↑(w) and G·C*↓(w) states, attaining values of 4.5 × 10−3 and 1.4 × 10−4, respectively (ε = 4), which allows them to be recorded experimentally (Fig. 5, 6, Tables 2, S8, S10 and S11†). In contrast, the population of the two other tautomerised G*·P↓(w) and G*·P↑(w) states of the G·P(WC) base pair increased only in 0.2 and 74.2 times, in comparison to analogous states of the G·C(WC) base pair in the continuum with ε = 4 (Fig. 5 and 6), and so remains inaccessible for experimental observation. All this testifies to the adequacy of the above presented representations of the mutagenic tautomerisation of the G·C(WC) base pair.
Results obtained in this paper significantly deepen and elaborate the classical Watson–Crick tautomeric hypothesis3 that links the nature of spontaneous point mutations occurring during DNA replication with the ability of the nucleotide bases to transfer from the canonical into the mutagenic tautomeric form. The developed representations significantly extend the possibility for the rational design of the promutagens by using nucleobases with targeted actions that are interesting from the point of view of both anticancer and antiviral therapy.
A rather promising task for the future is the clarification of the role of the local protein environment of the replisome71 on the discussed above tautomeric equilibrium, and also of the possible evolutionary value of the mutagenic tautomeric mobility of the canonical Watson–Crick DNA base pairs.
Acknowledgements
We gratefully appreciate the technical support and computational facilities of the joint computer cluster of the “SSI Institute for Single Crystals” of the National Academy of Sciences of Ukraine and the Institute for Scintillation Materials of the National Academy of Sciences of Ukraine, incorporated into the Ukrainian National Grid. This work was supported by the grant of the President of Ukraine to support the scientific research of young scientists for the year 2015 from the State Fund for Fundamental Research of Ukraine (project No. GP/F61/028), by the Grant of the NAS of Ukraine for young scientists for the years 2015–2016, and by the Scholarship of the President of Ukraine for young scientists for the years 2014–2016 given to O.O.B.References
- E. B. Freese, On the molecular explanation of spontaneous and induced mutations, Brookhaven Symp. Biol., 1959, 12, 63–75 Search PubMed.
- E. C. Friedberg, G. C. Walker, W. Siede, R. D. Wood, R. A. Schultz and T. Ellenberger, DNA repair and mutagenesis, ASM Press, Washington D.C, 2006 Search PubMed.
- J. D. Watson and F. H. C. Crick, The Structure of DNA, Cold Spring Harbor Symp. Quant. Biol., 1953, 18, 123–131 CrossRef CAS PubMed.
- V. I. Danilov, V. M. Anisimov, N. Kurita and D. Hovorun, MP2 and DFT studies of the DNA rare base pairs: the molecular mechanism of the spontaneous substitution mutations conditioned by tautomerism of bases, Chem. Phys. Lett., 2005, 412, 285–293 CrossRef CAS.
- C. Fonseca Guerra, F. M. Bickelhaupt, S. Saha and F. Wang, Adenine tautomers: relative stabilities, ionization energies, and mismatch with cytosine, J. Phys. Chem. A, 2006, 110, 4012–4020 CrossRef CAS PubMed.
- V. H. Harris, et al., The effect of tautomeric constant on the specificity of nucleotide incorporation during DNA replication: support for the rare tautomer hypothesis of substitution mutagenesis, J. Mol. Biol., 2003, 326, 1389–1401 CrossRef CAS PubMed.
- K. Bebenek, L. C. Pedersen and T. A. Kunkel, Replication infidelity via a mismatch with Watson–Crick geometry, Proc. Natl. Acad. Sci. U. S. A., 2011, 108, 1862–1867 CrossRef CAS PubMed.
- W. Wang, H. W. Hellinga and L. S. Beese, Structural evidence for the rare tautomer hypothesis of spontaneous mutagenesis, Proc. Natl. Acad. Sci. U. S. A., 2011, 108, 17644–17648 CrossRef CAS PubMed.
- I. J. Kimsey, K. Petzold, B. Sathyamoorthy, Z. W. Stein and H. M. Al-Hashimi, Visualizing transient Watson–Crick-like mispairs in DNA and RNA duplexes, Nature, 2015, 519, 315–320 CrossRef CAS PubMed.
- I. J. Kimsey, H. Zhou, H. Alvey and H. M. Al-Hashimi, Role of dynamic base pair polymorphism in the central dogma of molecular biology, J. Biomol. Struct. Dyn., 2015, 33(suppl. 1), 75–76 Search PubMed.
- O. O. Brovarets’ and D. M. Hovorun, Physicochemical mechanism of the wobble DNA base pairs Gua·Thy and Ade·Cyt transition into the mismatched base pairs Gua*·Thy and Ade·Cyt* formed by the mutagenic tautomers, Ukr. Bioorg. Acta, 2009, 8, 12–18 Search PubMed.
- O. O. Brovarets’ and D. M. Hovorun, Tautomeric transition between wobble A·C DNA base mispair and Watson–Crick-like A·C* mismatch: microstructural mechanism and biological significance, Phys. Chem. Chem. Phys., 2015, 17, 15103–15110 RSC.
- O. O. Brovarets’ and D. M. Hovorun, How many tautomerisation pathways connect Watson–Crick-like G*·T DNA base mispair and wobble mismatches?, J. Biomol. Struct. Dyn., 2015, 33, 2297–2315 Search PubMed.
- O. O. Brovarets’ and D. M. Hovorun, Novel physico-chemical mechanism of the mutagenic tautomerisation of the Watson–Crick-like A·G and C·T DNA base mispairs: a quantum-chemical picture, RSC Adv., 2015, 5, 66318–66333 RSC.
- O. O. Brovarets’ and D. M. Hovorun, A novel conception for spontaneous transversions caused by homo-pyrimidine DNA mismatches: a QM/QTAIM highlight, Phys. Chem. Chem. Phys., 2015, 17, 21381–21388 RSC.
- O. O. Brovarets’ and D. M. Hovorun, Wobble ↔ Watson–Crick tautomeric transitions in the homo-purine DNA mismatches: a key to the intimate mechanisms of the spontaneous transversions, J. Biomol. Struct. Dyn., 2015 DOI:10.1080/07391102.2015.1077737.
- D. Jacquemin, J. Zúñiga, A. Requena and J. P. Céron-Carrasco, Assessing the importance of proton transfer reactions in DNA, Acc. Chem. Res., 2014, 47, 2467–2474 CrossRef CAS PubMed.
- P.-O. Löwdin, Proton tunneling in DNA and its biological implications, Rev. Mod. Phys., 1963, 35, 724–732 CrossRef.
- O. O. Brovarets’ and D. M. Hovorun, Can tautomerisation of the A·T Watson–Crick base pair via double proton transfer provoke point mutations during DNA replication? A comprehensive QM and QTAIM analysis, J. Biomol. Struct. Dyn., 2014, 32, 127–154 Search PubMed.
- O. O. Brovarets’ and D. M. Hovorun, Proton tunneling in the A·T Watson–Crick DNA base pair: myth or reality?, J. Biomol. Struct. Dyn., 2015 DOI:10.1080/07391102.2015.1092886.
- O. O. Brovarets’ and D. M. Hovorun, Why the tautomerization of the G·C Watson–Crick base pair via the DPT does not cause point mutations during DNA replication? QM and QTAIM comprehensive analysis, J. Biomol. Struct. Dyn., 2014, 32, 1474–1499 Search PubMed.
- A. Padermshoke, Y. Katsumoto, R. Masaki and M. Aida, Thermally induced double proton transfer in GG and wobble GT base pairs: a possible origin of the mutagenic guanine, Chem. Phys. Lett., 2008, 457, 232–236 CrossRef CAS.
- O. O. Brovarets’, R. O. Zhurakivsky and D. M. Hovorun, DPT tautomerisation of the wobble guanine·thymine DNA base mispair is not mutagenic: QM and QTAIM arguments, J. Biomol. Struct. Dyn., 2015, 33, 674–689 Search PubMed.
- P. Strazewski and C. Tamm, Replication experiments with nucleotide base analogues, Angew. Chem., Int. Ed., 1990, 29, 36–57 CrossRef.
- O. O. Brovarets’, Y. P. Yurenko, I. Y. Dubey and D. M. Hovorun, Can DNA-binding proteins of replisome tautomerize nucleotide bases? Ab initio model study, J. Biomol. Struct. Dyn., 2012, 29, 1101–1109 Search PubMed.
- M. J. Frisch, G. W. Trucks, H. B. Schlegel, G. E. Scuseria, M. A. Robb, J. R. Cheeseman and J. A. Pople, GAUSSIAN 09 (Revision B.01), Gaussian Inc, Wallingford CT, 2010 Search PubMed.
- J. Tirado-Rives and W. L. Jorgensen, Performance of B3LYP density functional methods for a large set of organic molecules, J. Chem. Theory Comput., 2008, 4, 297–306 CrossRef CAS.
- R. G. Parr and W. Yang, Density-functional theory of atoms and molecules, Oxford University Press, Oxford, 1989 Search PubMed.
- C. Lee, W. Yang and R. G. Parr, Development of the Colle–Salvetti correlation-energy formula into a functional of the electron density, Phys. Rev. B: Condens. Matter Mater. Phys., 1988, 37, 785–789 CrossRef CAS.
- C. F. Matta, How dependent are molecular and atomic properties on the electronic structure method? Comparison of Hartree–Fock, DFT, and MP2 on a biologically relevant set of molecules, J. Comput. Chem., 2010, 31, 1297–1311 CAS.
- O. O. Brovarets’, R. O. Zhurakivsky and D. M. Hovorun, The physico-chemical mechanism of the tautomerisation via the DPT of the long Hyp*·Hyp Watson–Crick base pair containing rare tautomer: a QM and QTAIM detailed look, Chem. Phys. Lett., 2013, 578, 126–132 CrossRef.
- O. O. Brovarets’ and D. M. Hovorun, Atomistic understanding of the C·T mismatched DNA base pair tautomerization via the DPT: QM and QTAIM computational approaches, J. Comput. Chem., 2013, 34, 2577–2590 CrossRef PubMed.
- O. O. Brovarets’, R. O. Zhurakivsky and D. M. Hovorun, Is the DPT tautomerisation of the long A·G Watson–Crick DNA base mispair a source of the adenine and guanine mutagenic tautomers? A QM and QTAIM response to the biologically important question, J. Comput. Chem., 2014, 35, 451–466 CrossRef PubMed.
- O. O. Brovarets’ and D. M. Hovorun, How the long G·G* Watson–Crick DNA base mispair comprising keto and enol tautomers of the guanine tautomerises? The results of the QM/QTAIM investigation, Phys. Chem. Chem. Phys., 2014, 6, 15886–15899 RSC.
- S. P. Samijlenko, Y. P. Yurenko, A. V. Stepanyugin and D. M. Hovorun, Tautomeric equilibrium of uracil and thymine in model protein–nucleic acid contacts. Spectroscopic and quantum chemical approach, J. Phys. Chem. B, 2012, 114, 1454–1461 CrossRef PubMed.
- C. Peng, P. Y. Ayala, H. B. Schlegel and M. J. Frisch, Using redundant internal coordinates to optimize equilibrium geometries and transition states, J. Comput. Chem., 1996, 17, 49–56 CrossRef CAS.
- P. W. Atkins, Physical chemistry, Oxford University Press, Oxford, 1998 Search PubMed.
- H. P. Hratchian and H. B. Schlegel, Finding minima, transition states, and following reaction pathways on ab initio potential energy surfaces, in Theory and applications of computational chemistry: the first 40 years, ed. C. E. Dykstra, G. Frenking, K. S. Kim and G. Scuseria, Elsevier, Amsterdam, 2005, pp. 195–249 Search PubMed.
- O. O. Brovarets’ and D. M. Hovorun, DPT tautomerisation of the G·Asyn and A*·G*syn DNA mismatches: a QM/QTAIM combined atomistic investigation, Phys. Chem. Chem. Phys., 2014, 16, 9074–9085 RSC.
- O. O. Brovarets’, R. O. Zhurakivsky and D. M. Hovorun, Does the tautomeric status of the adenine bases change under the dissociation of the A*·Asyn Topal–Fresco DNA mismatch? A combined QM and QTAIM atomistic insight, Phys. Chem. Chem. Phys., 2014, 16, 3715–3725 RSC.
- O. O. Brovarets’ and D. M. Hovorun, Does the G·G*syn DNA mismatch containing canonical and rare tautomers of the guanine tautomerise through the DPT? A QM/QTAIM microstructural study, Mol. Phys., 2014, 112, 3033–3046 CrossRef.
- M. J. Frisch, M. Head-Gordon and J. A. Pople, Semi-direct algorithms for the MP2 energy and gradient, Chem. Phys. Lett., 1990, 166, 281–289 CrossRef CAS.
- P. C. Hariharan and J. A. Pople, The influence of polarization functions on molecular orbital hydrogenation energies, Theor. Chim. Acta, 1973, 28, 213–222 CrossRef CAS.
- R. Krishnan, J. S. Binkley, R. Seeger and J. A. Pople, Self-consistent molecular orbital methods. XX. A basis set for correlated wave functions, J. Chem. Phys., 1980, 72, 650–654 CrossRef CAS.
- R. A. Kendall, T. H. Dunning Jr and R. J. Harrison, Electron affinities of the first-row atoms revisited. Systematic basis sets and wave functions, J. Chem. Phys., 1992, 96, 6796–6806 CrossRef CAS.
- R. F. W. Bader, Atoms in molecules: a quantum theory, Oxford University Press, Oxford, 1990 Search PubMed.
- C. F. Matta, Modeling biophysical and biological properties from the characteristics of the molecular electron density, electron localization and delocalization matrices, and the electrostatic potential, J. Comput. Chem., 2014, 35, 1165–1198 CrossRef CAS PubMed.
- O. O. Brovarets’, Y. P. Yurenko and D. M. Hovorun, Intermolecular CH⋯O/N H-bonds in the biologically important pairs of natural nucleobases: a thorough quantum-chemical study, J. Biomol. Struct. Dyn., 2014, 32, 993–1022 Search PubMed.
- O. O. Brovarets’, Y. P. Yurenko and D. M. Hovorun, The significant role of the intermolecular CH⋯O/N hydrogen bonds in governing the biologically important pairs of the DNA and RNA modified bases: a comprehensive theoretical investigation, J. Biomol. Struct. Dyn., 2015, 33, 1624–1652 Search PubMed.
- V. Zoete and M. Meuwly, Double proton transfer in the isolated and DNA-embedded guanine–cytosine base pair, J. Chem. Phys., 2004, 121, 4377–4388 CrossRef CAS PubMed.
- O. O. Brovarets’ and D. M. Hovorun, How stable are the mutagenic tautomers of DNA bases?, Biopolym. Cell, 2010, 26, 72–76 CrossRef.
- Y. Lin, H. Wang, Y. Wu, S. Gao and H. F. Schaefer III, Proton-transfer in hydrogenated guanine–cytosine trimer neutral species, cations, and anions embedded in B-form DNA, Phys. Chem. Chem. Phys., 2014, 16, 6717–6725 RSC.
- J. Petrushka, L. C. Sowers and M. Goodman, Comparison of nucleotide interactions in water, proteins, and vacuum: model for DNA polymerase fidelity, Proc. Natl. Acad. Sci. U. S. A., 1986, 83, 1559–1562 CrossRef.
- J. W. Drake, B. Charlesworth, D. Charlesworth and J. F. Crow, Rates of spontaneous mutation, Genetics, 1998, 148, 1667–1686 CAS.
- M. Lynch, Rate, molecular spectrum, and consequences of human mutation, Proc. Natl. Acad. Sci. U. S. A., 2010, 107, 961–968 CrossRef CAS PubMed.
- I. J. Fijalkowska, R. M. Schaaper and P. Jonczyk, DNA replication fidelity in Escherichia coli: a multi-DNA polymerase affair, FEMS Microbiol. Rev., 2012, 36, 1105–1121 CrossRef CAS PubMed.
- O. O. Brovarets’ and D. M. Hovorun, Molecular logic of the spontaneous point mutagenesis: variation on a theme, Ukr. Bioorg. Acta, 2014, 12, 48–55 Search PubMed.
- E. Westhof, Isostericity and tautomerism of base pairs in nucleic acids, FEBS Lett., 2014, 588, 2464–2469 CrossRef CAS PubMed.
- M. Osborn, S. Person, S. Phillips and F. Funk, A determination of mutagen specificity in bacteria using nonsense mutants of bacteriophage T4, J. Mol. Biol., 1967, 26, 437–447 CrossRef CAS PubMed.
- E. B. Freese, The mutagenic effect of hydroxyaminopurine derivatives on phage T4, Mutat. Res., 1968, 5, 299–301 CrossRef CAS PubMed.
- M. F. Goodman, R. Hopkins and W. C. Gore, 2-Aminopurine-induced mutagenesis in T4 bacteriophage: a model relating mutation frequency to 2-aminopurine incorporation in DNA, Proc. Natl. Acad. Sci. U. S. A., 1977, 74, 4806–4810 CrossRef CAS.
- A. Ronen, 2-Aminopurine, Mutat. Res., 1979, 75, 1–47 Search PubMed.
- S. M. Watanabe and M. F. Goodman, On the molecular basis of transition mutations: frequencies of forming 2-aminopurine·cytosine and adenine·cytosine base mispairs in vitro, Proc. Natl. Acad. Sci. U. S. A., 1981, 78, 2864–2868 CrossRef CAS.
- E. Freese, The specific mutagenic effect of base analogue on phage T4, J. Mol. Biol., 1959, 1, 87–105 CrossRef CAS.
- O. O. Brovarets’ and D. M. Hovorun, The nature of the transition mismatches with Watson–Crick architecture: the G*·T or G·T* DNA base mispair or both? A QM/QTAIM perspective for the biological problem, J. Biomol. Struct. Dyn., 2015, 33, 925–945 Search PubMed.
- C. F. Matta, N. Castillo and R. J. Boyd, Extended weak bonding interactions in DNA: π-stacking (base–base), base–backbone, and backbone–backbone interactions, J. Phys. Chem. B, 2006, 110, 563–578 CrossRef CAS PubMed.
- J. P. Cerón-Carrasco, J. Zúñiga, A. Requena, E. A. Perpète, C. Michaux and D. Jacquemin, Combined effect of stacking and solvation on the spontaneous mutation in DNA, Phys. Chem. Chem. Phys., 2011, 13, 14584–14589 RSC.
- A. N. Nedderman, M. J. Stone, D. H. Williams, P. K. T. Lin and D. M. Brown, Molecular basis for methoxyamine-initiated mutagenesis: 1H nuclear magnetic resonance studies of oligonucleotide duplexes containing base-modified cytosine residues, J. Mol. Biol., 1993, 230, 1068–1076 CrossRef CAS PubMed.
- A. N. Nedderman, M. J. Stone, P. K. T. Lin, D. M. Brown and D. H. Williams, Base pairing of cytosine analogues with adenine and guanine in oligonucleotide duplexes: evidence for exchange between Watson–Crick and wobble base pairs using 1H NMR spectroscopy, J. Chem. Soc., Chem. Commun., 1991, 1357–1359 RSC.
- E. L. Mertz and L. I. Krishtalik, Low dielectric response in enzyme active site, Proc. Natl. Acad. Sci. U. S. A., 2000, 97, 2081–2086 CrossRef CAS PubMed.
- T. A. Baker and S. P. Bell, Polymerases and the replisome: machines within machines, Cell, 1998, 92, 295–305 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available: (i) computational details; (ii) profiles of the glycosidic parameters of the Watson–Crick DNA base pairs along the IRC of the tautomerisation; (iii) stationary structures on the reaction pathways of the tautomeric transformations of the A·T and G·C Watson–Crick DNA base pairs; (iv) energetic profiles of the reaction pathways of the tautomeric transformations of the A·T and G·C Watson–Crick DNA base pairs; (v) physico-chemical parameters of the specific intermolecular contacts in the investigated DNA base pairs and TSs of their wobbling tautomerisations; (vi) energetic and kinetic characteristics of the tautomerisations of the DNA base pairs containing canonical bases and mutagenic analogue of cytosine P, and also of the conformational interconversions of the mirror-symmetric enantiomers of the DNA base mispairs; (vii) interaction energies for the investigated DNA base pairs and TSs of their tautomeric wobbling; (viii) energetic and kinetic characteristics of the wobbling tautomerisations of the Watson–Crick DNA base pairs; (ix) ranges of the existence of the obtained patterns of the specific intermolecular interactions along the IRC of the Watson–Crick DNA base pairs tautomerisations; (x) movies of the tautomeric conversions of the A·T and G·C Watson–Crick DNA base pairs via the sequential DPT into the wobble mismatches containing mutagenic tautomers. See DOI: 10.1039/c5ra19971a |
| This journal is © The Royal Society of Chemistry 2015 |