
Selective esterification to produce microalgal biodiesel and enrich polyunsaturated fatty acid using zeolite as a catalyst
Abstract
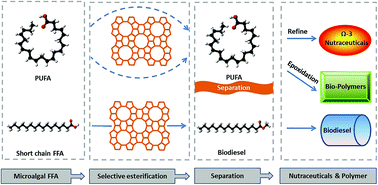
Microalgae can be both a promising biofuel feedstock and a source of polyunsaturated fatty acids (PUFA). This paper reports a novel integrated process that simultaneously produces biodiesel and enriches PUFA. It was accomplished by using zeolite as a selective catalyst that preferentially converts shorter-chain fatty acids (SCFA) into fatty acid methyl esters (FAME) (86% conversion for S. limacinum and 65% conversion for N. salina) and enriches high-value PUFA (70% for S. limacinum and 78% for N. salina) in the unreacted free fatty acid (FFA) stream. The esterification reaction rate was affected by acid strength and pore size, while the selectivity of zeolite increased as pore size of zeolite decreased. This approach allows production of high quality biodiesel and efficient PUFA enrichment. The unreacted PUFA can be further refined for nutraceutical or other applications to improve economic viability of microalgal biodiesel production.
 Please wait while we load your content...
Please wait while we load your content...

