
Genetically encoded fluorescence screening probe for MgrA, a global regulator in Staphylococcus aureus†
Yujie Wanga,
Hong Zhangb,
Qingzhou Zhanga,
Yujie Lianga,
Lin Maa,
Hongsheng Tanb,
Yuanzhi Lao*b,
Hongxi Xu*b and
Zigang Li*a
aSchool of Chemical Biology and Biotechnology, Peking University Shenzhen Graduate School, Shenzhen, Guangdong 518055, P. R. China. E-mail: lizg@pkusz.edu.cn
bSchool of Pharmacy, Shanghai University of Traditional Chinese Medicine, Shanghai, 201203, P. R. China. E-mail: xuhongxi88@gmail.com; laurence_ylao@163.com
First published on 8th October 2015
Abstract
Herein, we construct a novel cell-based fluorescence response screening system based on a single insertion of circularly permuted yellow fluorescent protein (cpYFP) to reflect the protein conformational change caused by MgrA–small molecules interaction. And this screening platform was applied for screening of Chinese herb extracts with two extracts identified from 351.
Staphylococcus aureus (S. aureus), a gram-positive bacteria, causes diseases ranging from minor skin infections to life-threatening endocarditis, pneumonia and sepsis.1 The repeated and improper use of antibiotics promotes drug-resistant bacteria including multiple methicillin-resistant (MRSAs) and vancomycin-resistant (VISAs and VRSAs) strains. A significant portion of postoperative infections are caused by these antibiotic-resistant strains, which are more difficult to treat and are associated with higher mortality.2 New strategies for combating these drug-resistant strains have become an increasingly important area of research. Inhibiting staphylococcal virulence expression by targeting non-essential virulence regulators represents an alternative approach to fight infections without selective pressure.3 Transcriptional regulator MgrA, a member of staphylococcal accessory A (SarA) and multiple antibiotic resistance regulator (MarR) protein family, plays a key role in the regulation of the expression of major virulence factors.4 Notably, mgrA-deleted S. aureus strains showed 1000–10
![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 000 fold virulence decrease in mouse infection model without showing detectable growth defects in cell based model.5
000 fold virulence decrease in mouse infection model without showing detectable growth defects in cell based model.5
We previously established a fluorescent anisotropy (FA)-based high-throughput screening platform and identified 5,5-methylenedisalicylic acid (MDSA) as an efficient blocker for MgrA–DNA interactions.6
As the first reported MgrA inhibitor, MDSA showed reduction of S. aureus virulence by modulating the expression of MgrA-regulating protein.6 Recently, α-methylene-γ-butyrolactone was also reported to lower S. aureus virulence via inhibition of SarA, SarR and MgrA.7 These reports validate the therapeutic potential of virulence-regulating transcriptional regulators such as MgrA.
Although the in vitro FA technology is broadly used in studying bio-molecule interactions, it easily generates false positive hits and is unsuitable for complex compound screenings such as herbal extracts. Ultimately, our aim is to identify compounds, which can suppress the toxicity of S. aureus without causing growth defects and this protein-based FA technology could not eliminate compounds with severe cell toxicity. Complex systems such as Chinese herbal medicinal extracts are valuable resources for identifying biologically active compounds. To date many herbal extracts, including single compounds isolated from these extracts, have been developed into drugs, such as Artemisia annua for the treatment of P. falciparum malaria and Fructus schisandrae chinensis (FSC) for hepatitis.8 The US Food and Drug Administration (FDA) approved plant extract mixtures Veregen for the treatment of human papillomavirus, and Fulyzaq for treatment of HIV-related diarrhea, in 2006 and 2012, respectively.9 Recently, danshen dripping pill, a traditional Chinese medicine mixture for cardiovascular diseases developed by Tasly Pharmaceutical Group, has entered a FDA approved phase III clinical trial.10 Based on these successes, plant extract mixtures are intriguing libraries for identifying new therapeutics.
Currently, most high throughput screening (HTS) systems depend on chemically labelled fluorescent or radioactive probes to screen small molecules.11
However, these labels can cast adverse effects on the binding properties being screened, which may cause false results or artifacts.12 Genetically encoded sensors based on the green fluorescent protein (GFP)-like proteins present another method showing significant advantages and are broadly utilized in living cells.13 In general, fluorescent biosensors contain two parts: a “sensor unit” that senses a cellular analyst or invades small molecules; and a “reporter unit” that generates a fluorescent readout. In the case of single cpFP-based sensors, the reporter cpFP's photophysical properties are altered in response to a conformational change in the sensor unit, which is caused by the small molecules being studied. The target proteins with cpFPs fused in sensitive domains would produce distinguishable fluorescent signals under different environments.14 Using this method, fluorescent sensors have been constructed based on GlnK1, Pericam, OxyR, OhrR, COSer and Frex for cell-based ATP, Ca2+, H2O2, organic hydroperoxides (OHPs), CO and NADH detections respectively.15 All these sensors showed high sensitivity and selectivity in cells. These properties indicate that genetically encoded fluorescent reporters may be utilized for HTS in living cells.
The molecular mechanism underlying the change in fluorescence intensity varied depending on cases. In study of Ca2+ sensor based on the crystal structure of GCaMP2–cpEGFP fusion protein, it was shown that a small hole existed at the site of permutation, which caused the chromosphere to be exposed to solvent. After binding to Ca2+ and M13, the conformational change in GCaMP2 induced sealing of the channel and restored fluorescence of cpEGFP.16
Herein, we report a cell-based screening platform of MgrA inhibitors with satisfying selectivity and sensitivity for screening complex chemical systems from Chinese herbal extracts. We constructed various MgrA–cpYFP fusion proteins as sensing probes in living bacterial cells and evolved the best candidate with significant in situ fluorescence changes upon treatment (Fig. 1).
 | ||
| Fig. 1 General design of screening method. cpYFP protein is inserted by two short linker between residues in MgrA. The structure changes caused by Chinese herbal extracts binding induce the increased fluorescence of this sensor. | ||
We have previously reported a near-UV (250–320 nm) circular dichroism (CD) spectra of MgrA and MgrA–MDSA that revealed upon binding, MDSA could cause significant conformational changes to MgrA.6 Computational docking indicated that MDSA can bind to the DNA-binding lobe flanked by Trp-48.6 MgrA shares some similar structural features with OhrR (Fig. S1†).17 In the organic hydroperoxide (OHP) sensor based on OhrR, cpYFP was inserted at the α-helix bundle 5, which breaks into two shorter helices (α-5a and α-5b) when OHPs are present.18 MgrA has a similar linkage between α-helix 5′ and α-helix 6′ (residues 109–111) (Fig. S1†). With this foundation we constructed a series of fluorescent probes that are MgrA–cpYFP fusion proteins with cpYFP insertion between S110/N111(SN), D114/K115(DK), S118/A119(SA) at helix 5′ and helix 6′. In addition, we also constructed fusion proteins with cpYFP inserted at helix 3′ (V59/T60, VT), helix 4′ (S71/P72, SP) at the helix-turn-helix (HTH) domain, which was crucial for DNA binding (Fig. 2a and b). The E. coli strains expressing fusion proteins such as SN, DK and SA were tested directly with MDSA. The SN, DK and SA expressing strains showed 43%, 30% and 20% fluorescence increases upon 400 μM MDSA treatment while fusion proteins VT and SP showed fluorescence decreases (Fig. 2c). The decrease in fluorescence in VT and SP is likely caused by permutation site channel expanding instead of channel sealing after MDSA binding to the sensor part of MgrA.
 | ||
| Fig. 2 The design and construction of fluorescent probe for MgrA inhibitor screening. (a) cpYFP insertion sites in MgrA (PDB No.2 BV6), MgrA dimer is shown and the insertion sites are only marked on blue monomer for clearance. (b) Constructed plasmid of fusion protein, VT means cpYFP is inserted between the 59V and 60T, same for SP, SN, DK and SA. (c) E. coli cells expressing cpYFP–MgrA fusion proteins are treated with 400 μM MDSA and relative emissions are recorded. (Excitation: 485 nm, emission: 520 nm). | ||
Similar to OhrR and OxyR, MgrA has a conserved cysteine (C12), which is sensitive to oxidation. The E. coli expressed SN protein and the purified protein both showed enhanced fluorescence upon treatment with tBHP (Fig. S2 and S3†). The fusion protein SN–MgrA–cpYFP (C12S) was then constructed by mutating cysteine into serine to reduce its oxidant sensitivity. As expected, this fusion protein showed minimal fluorescence increase upon oxidation (tBHP, CHP and H2O2) but the fluorescence response upon treatment with MDSA remained (Fig. S3†). When living E. coli cells expressing SN–MgrA–cpYFP (C12S) fusion protein were treated with MDSA and oxidants, the cells showed a fluorescence increase of up to 2.5 fold upon 400 μM MDSA treatment and only minimal fluorescence response with oxidants (Fig. 3b). Compared to SN–MgrA–cpYFP, the mutated SN–MgrA–cpYFP (C12S) system displayed higher sensitivity (Fig. 2c). The mutation eliminated background caused by endogenous or stimulated ROS (reactive oxidative species) in E.coli cells.19 Thus, the SN–MgrA–cpYFP (C12S) system was chosen for screening, and the Z′ of this platform was measured to be 0.78 (Fig. S5†).
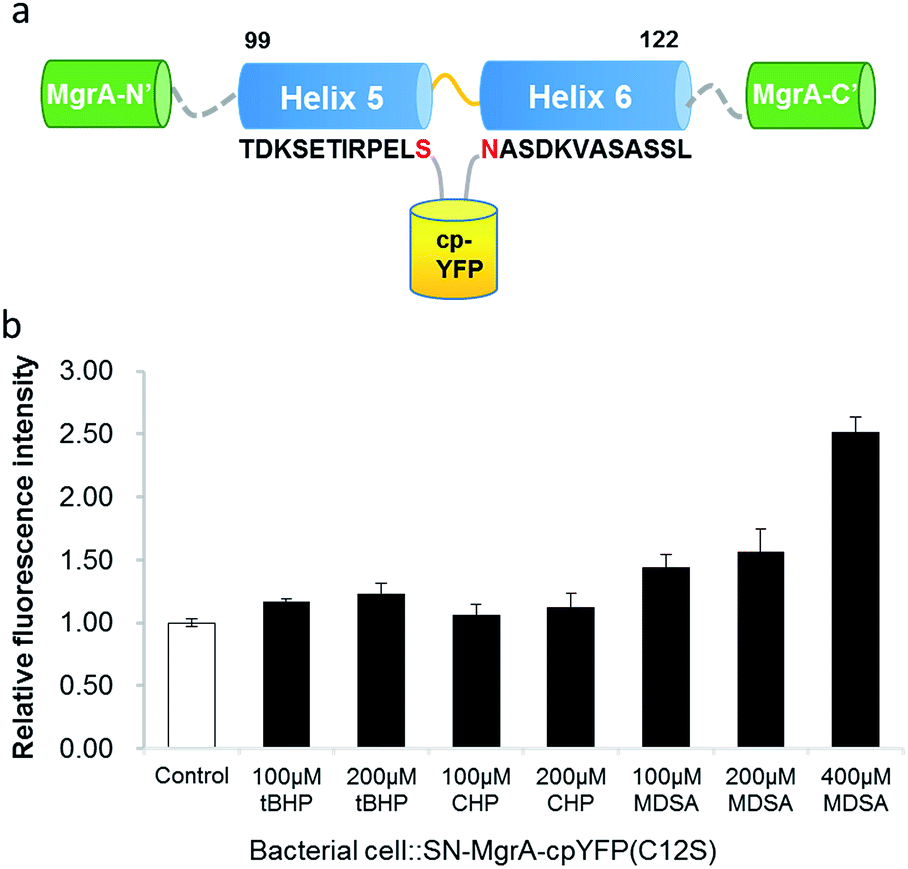 | ||
| Fig. 3 SN–MgrA–cpYFP–C12S is selected as fluorescent probe for inhibitor screening. (a) SN–MgrA–cpYFP–C12S contains a cpYFP inserted into the linker region between residues 110 and 111 in each monomer. (b) Fluorescence response of E. coli cells expressing SN–MgrA–cpYFP–C12S to different small molecule treatments. | ||
To test the selectivity of the screening method, we synthesized and purchased 11 MDSA derivatives with different substitute groups or structural modifications. All of these compounds lost most of their binding activity to MgrA and showed no fluorescent response to E. coli expressing SN–MgrA–cpYFP (C12S) (Fig S6 and S7†). While MgrA inhibitors 12 (IC50 30 μM) and 13 (IC50 32 μM),6 which have quite different structures compared with that of MDSA, could lead to moderate increase in fluorescence (about 1.2 fold), our result proved that, despite some imprecisions, the increase in fluorescence intensity observed was consistent with binding activity. These results also indicated that E. coli expressing SN–MgrA–cpYFP (C12S) is qualified to be used for MgrA binding molecular screening with good selectivity (Fig. S6 and S7†).
To test the performance of our screening method in the complex system, a library of 351 Chinese herbal extracts were screen screened in triplicates. 19 hits could induce over 1.9 fold fluorescent increase while 200 μM MDSA only get 1.5 fold increases. The reliability of this platform was then tested by the electrophoretic mobility shift assay (EMSA). A previous study showed that MgrA up-regulates the virulence factor α-toxin (hla) via recognizing its promoter region (5′-GTGTACAAACGAAAAAGTATCGTATGTATTTTTAATATAG-3′). In our EMSA assays, 15 out of the 19 hits could partially or fully dissociate the binding of MgrA to the hla promoter at 0.8 mg mL−1 (Fig. S8†), suggesting these 15 hits disrupt the MgrA–DNA binding by interacting with MgrA. Our aim is to identify virulence regulating agents which don't disrupt the growth of S. aureus, all the hits were then subjected to growth curve assay examination. None of 19 hits showed intermediate toxicity towards E. coli cells and S. aureus Newman strain, two hits showed detrimental effects on S. aureus mgrA-deleted strain growth after 12 hour treatment. Both assays confirmed our method's screening efficiency (Table S1†).
After excluding 4 false positives by EMSA and 2 cell toxic hits by a growth inhibition assay, 13 hits remained. 2 most effective extract mixtures from the bark of Cinnamomum cassia Presl (LG-026) and Sargassum pallidum (Turn.) C. Ag (LG-312) were picked for furthers studies.
C. cassia (Lauraceae) is an evergreen tree originating in southern China, and widely cultivated in India, Indonesia, Laos, Malaysia, Thailand, and Vietnam. Pharmacological studies of the dried bark of C. cassia (cortex cinnamomi), showed anti-diabetes, anti-virus, anti-inflammatory and anticancer effects; cortex cinnamomi also shows xanthine oxidase inhibitory activity and immunosuppressive activities.20 Sargassum pallidum (Turn) C. Ag is another kind of TCM, distributed extensively in the Yellow Sea and the East China Sea, it is widely used for phlegm elimination and detumescence.21 Pharmacological investigation of S. pallidum showed antitumor, antioxidant and immunity-enhancing activities.22 In our study, LG-026 and LG-312 showed strong interaction with MgrA and effectively dissociate the MgrA-hla interaction at 0.1 and 0.4 mg mL−1 respectively (Fig. 4a, similar results are obtained with MgrA C12S mutant, shown in Fig. S10†). These EMSA results suggest these two extracts may inhibit the expression of hla in S. aureus, which was further confirmed by western blot assays. The treatment with both LG-026 and LG-312 extracts could repress the expression of α-hemolysin in S. aureus Newman strain. The expression of α-hemolysin (hla), which is up-regulated by MgrA, is remarkably inhibited at 400 μg mL−1 for LG-026 and 900 μg mL−1 for LG-312 (Fig. 4b). There is no significant hla expression change in mgrA-deleted strain when treated with LG-026 and LG-312 (Fig. S11†). The α-hemolysin released by S. aureus can dissolve sheep blood in agar and produce a clearance zone. When Newman strains treated with LG-026 (1 mg mL−1) and LG-312 (1 mg mL−1), the clearance zones on 5% sheep blood agar significantly decreases, which further confirms the inhibition of hla expression (Fig. 4c). Real-time polymerase chain reaction (RT-PCR) experiments further validated the effects of these two extracts on two selected genes hla and spa (MgrA up-regulates hla and down-regulates spa).23 Quantitative-PCR (qPCR) results showed that the expression levels of hla in the Newman strains significantly decrease while spa expression levels increase with treatment of LG-026 or LG-312 (Fig. 4d). MgrA negatively regulated the efflux pumps such as NorA, NorB, NorC and Tet38, which account for the antibiotic resistance.5 In the plate sensitivity assay, increased antibiotic resistance was found in the wild type and the complementary strain in the presence of LG-026 and LG-312, which was consistent with observations from our previous work on MDSA (Fig. S12†).6 All these biochemical and phenotypic studies suggest that both herbal extracts LG-026 and LG-312 alter the virulence factor expression levels via an MgrA-dependent manner.
 | ||
| Fig. 4 LG-026 and LG-312 regulate the expression of virulence factors in an MgrA-dependent way. (a) EMSA of LG-026 and LG-312 with hla promoter. (b) Expression of hla in S. aureus shows a concentration dependent manner of LG-026 and LG-312. (c) Hemolysis of hla on 5% sheep blood agar plate is inhibited by 1 mg ml−1 LG-026 and LG-312. (d) mRNA expression of hla and spa decreases and increases respectively in a dose dependent manner when treated with LG-026 and LG-312, means and standard errors of triplicates are shown. *P < 0.05, **P < 0.01 and ***P < 0.001, when compared with control. | ||
Here, we constructed a fluorescent probe for MgrA inhibitor screening, which is facile and can be applied for complex system screening such as Chinese herbal extracts. This probe will precisely reflect the conformational change of MgrA upon binding of small molecules with high sensitivity and selectivity. Among 19 hits picked from the initial screening, 15 of them were effective in EMSA validation, while none of them showed immediate toxicity and only two showed detrimental effects for cell growth in 12 hour growth experiments. 2 best hits were further validated with biochemical and phenotypic assays and both of them decreased S. aureus virulence via an MgrA-depended pathway. Virulence regulation factors such as SarA, SarZ share similar functions and structural features with MgrA and deletion of these genes resulted in virulence reduction of S. aureus. This platform is also promising for screening of SarA and SarZ inhibitors.4
In summary, to the best of our knowledge, this is the first demonstration of a conformation sensitive fluorescent senor of regulator proteins being used for small molecule inhibitor screening with high sensitivity and selectivity. Notably, this method could significantly eliminate off-target effects comparing with previous screening method based on FA and yield highly reliable results from screening of complex systems as Chinese medicinal herbal extracts. Further separation and characterization of effective compounds may have potential to be further developed into novel anti-microbials.
Acknowledgements
This work was financially supported by Natural Science Foundation of China Grant 21102007 and 21372023, MOST 2015DFA31590, the Shenzhen Science and Technology Innovation Committee SW201110018, SGLH20120928095602764, ZDSY20130331145112855, the Shenzhen Peacock Program (KQTD201103), and Basic Research Program of Shanghai University of Traditional Chinese Medicine (L1364).Notes and references
- (a) A. L. Panlilio, D. H. Culver, R. P. Gaynes, S. Banerjee, T. S. Henderson, J. S. Tolson and W. J. Martone, Infect. Control Hosp. Epidemiol., 1992, 13, 582–586 CrossRef CAS; (b) F. D. Lowy, N. Engl. J. Med., 1998, 339, 520–532 CrossRef CAS PubMed.
- A. J. Alanis, Arch. Med. Res., 2005, 36, 697–705 CrossRef PubMed.
- (a) Y. M. Lee, F. Almqvist and S. J. Hultgren, Curr. Opin. Pharmacol., 2003, 3, 513–519 CrossRef CAS PubMed; (b) A. E. Clatworthy, E. Pierson and D. T. Hung, Nat. Chem. Biol., 2007, 3, 541–548 CrossRef CAS PubMed.
- (a) C. P. Gordon, P. Williams and W. C. Chan, J. Med. Chem., 2013, 56, 1389–1404 CrossRef CAS PubMed; (b) A. L. Cheung, K. A. Nishina, M. P. Trotonda and S. Tamber, Int. J. Biochem. Cell Biol., 2008, 40, 355–361 CrossRef CAS PubMed; (c) I. C. Perera and A. Grove, J. Mol. Cell Biol., 2010, 2, 243–254 CrossRef CAS PubMed.
- (a) P. R. Chen, T. Bae, W. A. Williams, E. M. Duguid, P. A. Rice, O. Schneewind and C. He, Nat. Chem. Biol., 2006, 2, 591–595 CrossRef CAS PubMed; (b) T. Bae, A. K. Banger, A. Wallace, E. M. Glass, F. Åslund, O. Schneewind and D. M. Missiakas, Proc. Natl. Acad. Sci. U. S. A., 2004, 101, 12312–12317 CrossRef CAS PubMed.
- (a) F. Sun, L. Zhou, B. C. Zhao, X. Deng, H. Cho, C. Yi, X. Jian, C. X. Song, C. H. Luan, T. Bae, Z. G. Li and C. He, Chem. Biol., 2011, 18, 1032–1041 CrossRef CAS PubMed; (b) W. H. Zheng, Y. J. Liang, H. Zhao, J. N. Zhang and Z. G. Li, ChemBioChem, 2015, 16, 1035–1040 CrossRef CAS PubMed.
- M. H. Kunzmann, N. C. Bach, B. Bauer and S. A. Sieber, Chem. Sci., 2014, 5, 1158–1167 RSC.
- (a) S. Y. Pan, S. F. Zhou, S. H. Gao, Z. L. Yu, S. F. Zhang, M. K. Tang, J. N. Sun, D. L. Ma, Y. F. Han and W. F. Fong, J. Evidence-Based Complementary Altern. Med., 2013, 2013 Search PubMed; (b) N. White, Antimicrob. Agents Chemother., 1997, 41, 1413 CAS.
- X. Lei, J. Chen, C. X. Liu, J. Lin, J. Lou and H. C. Shang, Chin. J. Integr. Med., 2014, 20, 403–408 CrossRef PubMed.
- L. Zhou, Z. Zuo and M. S. Chow, J. Clin. Pharmacol., 2005, 45, 1345–1359 CrossRef CAS PubMed.
- (a) J. Inglese, R. L. Johnson, A. Simeonov, M. Xia, W. Zheng, C. P. Austin and D. S. Auld, Nat. Chem. Biol., 2007, 3, 466–479 CrossRef CAS PubMed; (b) D. A. Bachovchin, S. J. Brown, H. Rosen and B. F. Cravatt, Nat. Biotechnol., 2009, 27, 387–394 CrossRef CAS PubMed.
- L. M. Mayr and D. Bojanic, Curr. Opin. Virol., 2009, 9, 580–588 CAS.
- (a) A. E. Palmer, Y. Qin, J. G. Park and J. E. McCombs, Trends Biotechnol., 2011, 29, 144–152 CrossRef CAS PubMed; (b) W. B. Frommer, M. W. Davidson and R. E. Campbell, Chem. Soc. Rev., 2009, 38, 2833–2841 RSC.
- G. S. Baird, D. A. Zacharias and R. Y. Tsien, Proc. Natl. Acad. Sci. U. S. A., 1999, 96, 11241–11246 CrossRef CAS.
- (a) V. V. Belousov, A. F. Fradkov, K. A. Lukyanov, D. B. Staroverov, K. S. Shakhbazov, A. V. Terskikh and S. Lukyanov, Nat. Methods, 2006, 3, 281–286 CrossRef CAS PubMed; (b) J. Berg, Y. P. Hung and G. Yellen, Nat. Methods, 2009, 6, 161–166 CrossRef CAS PubMed; (c) B. Simen Zhao, Y. Liang, Y. Song, C. Zheng, Z. Hao and P. R. Chen, J. Am. Chem. Soc., 2010, 132, 17065–17067 CrossRef PubMed; (d) Y. Zhao, J. Jin, Q. Hu, H. M. Zhou, J. Yi, Z. Yu, L. Xu, X. Wang, Y. Yang and J. Loscalzo, Cell Metab., 2011, 14, 555–566 CrossRef CAS PubMed; (e) T. Nagai, A. Sawano, E. S. Park and A. Miyawaki, Proc. Natl. Acad. Sci. U. S. A., 2001, 98, 3197–3202 CrossRef CAS PubMed; (f) Y. G. Ermakova, D. S. Bilan, M. E. Matlashov, N. M. Mishina, K. N. Markvicheva, O. M. Subach, F. V. Subach, I. Bogeski, M. Hoth and G. Enikolopov, Nat. Commun., 2014, 5, 5222 CrossRef CAS PubMed.
- (a) Q. Wang, B. Shui, M. I. Kotlikoff and H. Sondermann, Structure, 2008, 16, 1817–1827 CrossRef CAS PubMed; (b) J. Akerboom, J. D. Rivera, M. M. Guilbe, E. C. Malave, H. H. Hernandez, L. Tian, S. A. Hires, J. S. Marvin, L. L. Looger and E. R. Schreiter, J. Biol. Chem., 2009, 284, 6455–6464 CrossRef CAS PubMed; (c) R. H. Newman, M. D. Fosbrink and J. Zhang, Chem. Rev., 2011, 111, 3614–3666 CrossRef CAS PubMed.
- M. Hong, M. Fuangthong, J. D. Helmann and R. G. Brennan, Mol. Cell, 2005, 20, 131–141 CrossRef CAS PubMed.
- K. J. Newberry, M. Fuangthong, W. Panmanee, S. Mongkolsuk and R. G. Brennan, Mol. Cell, 2007, 28, 652–664 CrossRef CAS PubMed.
- D. O. de Orué Lucana, I. Wedderhoff and M. R. Groves, J. Signal Transduction, 2012, 2012, 605905 Search PubMed.
- (a) Y. M. Yan, P. Fang, M. T. Yang, N. Li, Q. Lu and Y. X. Cheng, J. Ethnopharmacol., 2015, 165, 141–147 CrossRef CAS PubMed; (b) Y. Han, H. W. Jung, H. S. Bae, J. S. Kang and Y. K. Park, Pharm. Biol., 2013, 51, 961–967 CrossRef PubMed; (c) C. F. Yeh, J. San Chang, K. C. Wang, D. E. Shieh and L. C. Chiang, J. Ethnopharmacol., 2013, 147, 321–326 CrossRef CAS PubMed; (d) A. M. Reddy, J. H. Seo, S. Y. Ryu, Y. S. Kim, K. Min and Y. Kim, Planta Med., 2004, 70, 823–827 CrossRef CAS PubMed; (e) J. C. Liao, J. S. Deng, C. S. Chiu, W. C. Hou, S. S. Huang, P. H. Shie and G. J. Huang, J. Evidence-Based Complementary Altern. Med., 2012, 2012, 129152 Search PubMed; (f) M. A. Lee, H. J. Park, H. J. Chung, W. K. Kim and S. K. Lee, J. Nat. Prod., 2013, 76, 1278–1284 CrossRef CAS PubMed; (g) T. M. Ngoc, N. M. Khoi, N. X. Nhiem, B. H. Tai, D. van Don, H. van Luong, D. C. Son and K. Bae, Bioorg. Med. Chem. Lett., 2012, 22, 4625–4628 CrossRef CAS PubMed.
- L. Liu, M. Heinrich, S. Myers and S. A. Dworjanyn, J. Ethnopharmacol., 2012, 142, 591–619 CrossRef PubMed.
- (a) H. Ye, K. Wang, C. Zhou, J. Liu and X. Zeng, Food Chem., 2008, 111, 428–432 CrossRef CAS PubMed; (b) R. L. Zhang, W. D. Luo, T. N. Bi and S. K. Zhou, Molecules, 2012, 17, 8419–8429 CrossRef CAS PubMed.
- S. Ingavale, W. van Wamel, T. T. Luong, C. Y. Lee and A. L. Cheung, Infect. Immun., 2005, 73, 1423–1431 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available: Methods and ESI figures. See DOI: 10.1039/c5ra11455a |
| This journal is © The Royal Society of Chemistry 2015 |