
Alkyl mono- and di-glucoside sugar vesicles as potential drug delivery vehicles: detecting drug release using fluorescence
Abstract
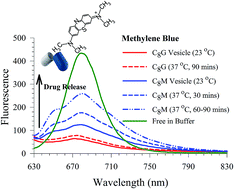
The use of alkyl glycosides as alternatives to other surfactants for pharmaceutical applications has received much interest due to their specific carbohydrate interactions, low inherent toxicity, and biodegradability. We have formulated sugar-based vesicles that carry mono- and disaccharide groups using octyl glucoside and maltoside, respectively. The physicochemical properties of the vesicles including size, surface charge, shape, and effect of the cholesterol content on the vesicle stability were investigated using dynamic light scattering and transmission electron microscopy. Stable glucoside (C8G) and maltoside (C8M) vesicles were achieved by incorporation of 20 mol% cholesterol, and 8 mol% dicetyl phosphate charge additives. A model small hydrophilic methylene blue (MB) dye was encapsulated into C8G and C8M, and the leakage of the dye was determined by examining the corresponding fluorescence signal in frequency and time domains. The results indicate a tight binding of MB in C8G, as reflected in the short fluorescence lifetime (τ < 0.10 ns) compared to MB in C8M (τ = 0.28 ns). This was in agreement with the leakage efficiency of MB which was measured at physiological temperature to be almost zero in MB–C8G, while a ca. 17% leakage was detected for MB–C8M after 30 minutes. The latter shows a ca. 44% leakage after 60 minutes. The larger head group in C8M increases the membrane fluidity, leading to a more flexible vesicle with more leakage efficiency. A local basic environment was observed in dye-encapsulated vesicles, and was attributed to through space interactions between MB and each of the sugar polar sites and the localized water molecules. The vesicles were non-cytotoxic at low concentrations. The current work shows the usefulness of using C8M as a carrier intended for fast release of hydrophilic drugs.
 Please wait while we load your content...
Please wait while we load your content...

