
Gallic acid and methyl-3-O-methyl gallate: a comparative study on their effects on prostate cancer stem cells
G. Civenni†
a,
M. G. Iodice†b,
S. F. Nabavic,
S. Habtemariamd,
S. M. Nabavic,
C. V. Catapano‡
a and
M. Daglia‡*b
aInstitute of Oncology Research (IOR), Oncology Institute of Southern Switzerland (IOSI), Via Vela 6, 6500 Bellinzona, Switzerland. E-mail: carlo.catapano@ior.iosi.ch
bDepartment of Drug Sciences, Medicinal Chemistry and Pharmaceutical Technology Section, Pavia University, Via Taramelli 12, 27100 Pavia, Italy. E-mail: maria.daglia@unipv.it
cApplied Biotechnology Research Center, Baqiyatallah University of Medical Sciences, Tehran, Iran
dPharmacognosy Research Laboratories, Medway School of Science, University of Greenwich, Chatham-Maritime, Central Avenue, Kent ME4 4TB, UK
First published on 21st July 2015
Abstract
Growing evidence shows that gallic acid (3,4,5-trihydroxybenzoic acid, GA) and methyl-3-O-methyl gallate (M3OMG) possess physiological and pharmacological activities as antioxidant and anti-inflammatory agents. At the molecular level, many chronic diseases, including cancer, are caused by oxidative stress and deregulated inflammatory responses. Several lines of evidence support a role for oxidative stress and inflammation in cancer. Moreover, one of the most important links between inflammation and cancer is nuclear factor κB (NF-κB), a transcription factor regulating the expression of genes involved in inflammation and immune responses. The aim of the present study is twofold: to evaluate and compare the ability of GA and M3OMG to inhibit NF-κB transcriptional activity, and to address their properties in different prostate cancer cell subpopulations. NF-κB transcriptional activity was found to be higher in prostatosphere than in prostate cancer cells cultured as an adherent monolayer and was efficiently reduced by GA and M3OMG. M3OMG exhibited stronger inhibitory activity in cancer cells with stem-like properties, whereas GA exhibited higher potency in the more differentiated cancer cells and was more effective in blocking cellular proliferation. Moreover, M3MOG was a stronger inhibitor of prostatosphere formation than GA. These results show that GA and M3OMG inhibit NF-κB transcriptional activity and growth of prostate cancer cells, with differential effects on cells with different proliferative, self-renewal and tumourigenic potential.
1. Introduction
Gallic acid (3,4,5-trihydroxybenzoic acid, GA) is a secondary metabolite ubiquitously distributed in plant foods. The amounts of GA vary according to plant species and, within a plant species, according to different environmental factors, such as UV radiation, microbial infections, insect attack and chemical stressors. Rubus, Fragaria (Rosaceae) and Vaccinium (Ericaceae) genera are important sources of GA, which is also present in green, oolong and black tea (Camellia sinensis L.), and in grapes and wine (Vitis vinifera L.).1 In addition to GA in free form, many plant foods and beverages contain GA in esterified forms. Green and semi-fermented or fermented teas are the most important sources of the GA esterified forms of (−) epicatechin.2 Recent studies have reported the isolation of methyl-3-O-methyl gallate (M3OMG), another GA esterified derivative extracted from Peltiphyllum peltatum (Torr.) Engl. (Saxifragaceae), which is a rhizomatous perennial herb, used as food and in traditional medicine.3 A comparative in vitro study on the antioxidant/prooxidant activities of GA and M3OMG revealed that M3OMG, in contrast to GA, shows antioxidant activity without showing prooxidant activity. Previous in vivo studies reported that GA reduces oxidative stress and enhances the levels of glutathione (GSH), glutathione peroxidase (GPX), GSH reductase and GSH S-transferase, in hepatic tissue, and it reduces catalase (CAT) in serum.4,5 Moreover, M3OMG mitigates the NaF-induced cardiotoxicity by modificating abnormality in the oxidant/antioxidant status, measured as changes in thiobarbituric acid reactive substance (TBAR) levels, and it reduced GSH and antioxidant enzyme activities (i.e. superoxide dismutase (SOD) and CAT), in rat cardiac tissues.6,7 Our recent research on peripheral blood mononuclear cells (PBMCs) and EVC-304 cells, treated with sub-toxic concentrations of M3OMG, has shown that M3OMG influences the expression levels of miR-17-3p.8 The miR-17-3p is a microRNA with important roles in the regulation of cellular redox status, interfering with the transcription of the mRNAs coding for SOD, glutathione peroxidase-2 (GPX2) and thioredoxin reductase-2 (TRX2). Therefore, these results suggest the presence of a potential mechanism of action of M3OMG, which, by inducing the reduction of miR-17-3p levels in cells, allows the antioxidant enzymes to perform their effects. Moreover, GA acts as competitive inhibitor of COX-1 and COX-2 (with more affinity toward COX-2),9 which play important role in inflammation, because they are key enzymes in the synthesis of prostaglandins from arachidonic acid. Another crucial effect of GA is the induction of apoptosis.10 In fact, GA was able to activate the proapoptotic proteins Fas, FasL and p53.Prostate cancer is the most common form of cancer in men. It is considered one of the leading causes of cancer-related death in Western countries.11 On the other hand, the lowest incidence rate of prostate cancer is typically found in Asian countries, such as China.12 Apart from age, the differences between industrialized Western countries and non-industrialized East Asian countries suggest that lifestyle is a very important factor, which contributes to prostate cancer development. Given these premises, diet may play a major role in prostate cancer as well as in other types of cancer. Many epidemiological and animal studies have shown that a diet rich in fruits and vegetables decreased the incidence of some types of cancer.13,14 This suggests that good dietary habits could reduce the risk of certain types of cancer.
At molecular level, many chronic diseases, including cancer, are caused by deregulated inflammatory responses.15 One of the most important links between inflammation and cancer is NF-κB.16 NF-κB is an ubiquitous transcription factor that regulates the expression of genes implicated in tumour cells' transformation, survival, proliferation, invasion, angiogenesis and metastasis.16 We recently reported that constitutive activation of NF-κB has a major role in prostate cancer progression, and it is linked to aggressive disease with early recurrence, metastases and poor clinical outcome.17 Metastases are the leading cause of death in cancer, and the discovery of new therapeutic strategies for highly metastatic and invasive cancers is very important in cancer research.
GA has various biological activities. Increasing evidence suggests the effectiveness of GA as anticancer agent, due to its antimetastatic effects through the inhibition of cancer cell migration and invasion. Recently, the extract of Phullanthus urinaria L. (PUE, rich in GA and derivatives) has been shown to inhibit the invasion and migration of lung cancer cell lines.17 Moreover, PUE exerted inhibitory effects on NF-κB nuclear translocation and DNA-binding activity. GA was reported to increase the level of the inhibitory subunit IκB, in a dose-dependent manner, with consequent inhibition of NF-κB signalling. These studies suggest that the suppression of NF-κB activation by GA and derivatives could play a role in chemoprevention and treatment of cancer.18–20
The research to date has shown potentially useful biological activity and interesting physical–chemical properties (such as lipophilicity) of GA and M3OMG, which make them attractive candidates for therapeutic applications in cancer research. However, there has been limited information on the activity of M3OMG in human cancer cell lines and on human experimental models. Therefore, the aim of this investigation was to verify the potential anti-cancer effect of M3OMG. The effects of M3OMG on human prostate cancer-cell cultures were examined in comparison with the effects of GA. The effects of these compounds were further investigated in relation to the activation state of the NF-κB pathway in the bulk cancer cell population and in the subpopulation of cancer stem-like cells (CSC). The latter, according to emerging evidence, are involved in tumour maintenance and progression, therapy resistance, and metastasis.21,22
2. Results and discussion
2.1 NF-κB activity in bulk cancer cells and cancer stem-cell compartment
DU145 prostate cancer cells, derived from a brain metastasis and considered highly malignant and tumourigenic cells, were cultured in adherent and non-adherent sphere-forming conditions, as previously described.23,24 Our previous studies showed that prostatospheres of DU145 cells, grown in sphere-forming conditions, are enriched in cells with CSC properties and are more tumourigenic than DU145 cells maintained as adherent monolayers.23,24 Moreover, the activity of NF-κB transcription factor is higher in the stem-cell enriched subpopulation than in bulk prostate cancer cells.25,26 In recent years, there has been an increasing interest in the inhibition of NF-κB pathway as potential tool to target CSC. Therefore, to address the status of NF-κB pathway in our experimental system, a set of known NF-κB target genes (i.e. IL-6, COX-2 and ESE-1) was assessed.16,17 Gene expression was measured by qRT-PCR in DU145 cells, grown either in adherent conditions or as floating spheres. After 7 days of growth either in adherent monolayer or non-adherent conditions, DU145 cells were collected and lysed to isolate total mRNA. The level of mRNA coding for IL-6, COX-2, and ESE-1 was significantly upregulated in cells grown in non-adherent conditions, confirming the hyper-activation of NF-κB in the CSC subpopulation (Fig. 1). | ||
| Fig. 1 IL-6, COX-2 and ESE-1 mRNA expression levels in DU145 adherent (Adh) cells and in prostatospheres (PS). Data are presented as fold change of mRNA level relative to adherent cells. | ||
2.2 Evaluation of the effect of M3OMG on NF-κB transcriptional targets
To determine the effect of M3OMG on the NF-κB pathway in CSCs, DU145 cells were cultured in sphere-forming conditions, and then the prostatospheres were treated with M3OMG (Fig. 2A). In parallel, DU145 cells, grown in adherent conditions, were treated with M3OMG in the same conditions (Fig. 2B). At the end of the treatment, cells were collected and mRNA was isolated to measure the expression of mRNA coding for IL-6 and ESE-1 by qRT-PCR. M3OMG significantly decreased the expression levels of IL-6 and ESE-1 mRNA in cells grown in sphere-forming conditions at tested concentrations. By contrast, when treatment was done in adherent conditions, at the low concentration (2.5 μM) the expression of these genes was slightly, but not significantly, higher, compared to control cells (Fig. 2B). Only at the highest concentration (10 μM) M3OMG decreased significantly IL-6 and ESE-1 mRNA expression levels in adherent growing cells (Fig. 2B). | ||
| Fig. 2 IL-6 and ESE-1 mRNA expression levels in DU145 cells cultured (A) in sphere forming conditions (PS, black bars) and (B) in adherent conditions (Adh, empty bars) after the treatment with M3OMG at the concentrations of 2.5 and 10 μM. | ||
2.3 Evaluation of the effect of GA on NF-κB transcriptional targets
To determine the effect of GA on the NF-κB signalling in CSCs, DU145 cells were grown either in sphere-forming (Fig. 3A) or in adherent (Fig. 3B) conditions and afterwards treated with GA. At the end of the incubation time, the levels of mRNA coding for IL-6 and ESE-1 were reduced by GA at the tested concentrations, in both groups of cells. In contrast to M3OMG, GA showed greater effect on cells grown in adherence compared to prostatospheres (Fig. 3B). In fact, at the concentration of 2.5 μM, GA reduced the expression levels of mRNA coding for IL-6 and ESE-1 by 74% and 62% in adherence, and by 35% and 42% in sphere cells, respectively. These results show that GA at the lowest concentration (2.5 μM) is a potent inhibitor of NF-κB transcriptional activity and might have higher efficiency on the more differentiated cancer cells grown in adherent monolayer cultures. | ||
| Fig. 3 IL-6 and ESE-1 mRNA expression levels in DU145 cells cultured (A) in sphere forming conditions (PS, black bars) and (B) in adherent conditions (Adh, empty bars) after the treatment with GA at the concentrations of 2.5 and 10 μM. | ||
GA has been reported to interfere with the NF-κB pathway and to have anti-carcinogenic effects, related to its antioxidant activity and ability to interfere with several cellular systems.18–20 M3OMG has different physical–chemical and biological properties compared to GA, such as increased antioxidant activity, stability and lipophilicity.8 However, previous studies have not investigated the ability of M3OMG to interfere with the NF-κB pathway and to act as anticancer agent. The results reported so far has shown that GA and M3OMG suppress transcription of NF-κB target genes in the CSC compartment and in more differentiated bulk cancer cells. In particular, GA decreased the expression of IL-6 and ESE-1 more effectively in adherent than in prostatosphere cells. Conversely, M3OMG showed greater inhibitory activity in prostatospheres than in adherent cells, and it was more active than GA in CSC-enriched prostatospheres.
2.4 Evaluation of the effect of GA and M3MOG on the CSC compartment
From the previous experiments, it is clear that, though with different efficacy, GA and M3MOG are able to block the transcription of NF-κB target genes in DU145 cells grown both in adherent and in non-adherent conditions. Since NF-κB activity is higher in CSC-enriched prostatospheres derived from DU145 cells, we evaluated the effects of GA and M3OMG on the ability of these cells to survive in non-adherent conditions and to grow as prostatospheres. DU145 cells were cultured in sphere-forming conditions, in the presence of different concentrations of GA (Fig. 4A) and M3OMG (Fig. 4B). GA and M3OMG were able to reduce prostatosphere formation in a dose-dependent manner, with half maximal inhibitory concentration (IC50) values of 730 nM and 330 nM, respectively. The lower IC50 registered for M3OMG compared with that determined for GA indicated that the former had potentially a greater impact on the sphere-forming ability of DU145 cells, perhaps in line with the greater effect on the NF-κB signalling in prostatosphere cells. Overall, these results has shown that both compounds were able to prevent prostatosphere growth. However M3OMG is more efficient in affecting the cancer stem-like component than GA. | ||
| Fig. 4 Effect of GA (A, black bars) and M3OMG (B, empty bars), tested at different concentrations (ranging from 0 to 10 μM), on the proliferation of DU145 cells cultured in sphere forming conditions. Data are presented as percentage of untreated control cells. | ||
2.5 Effect of GA and M3MOG on the proliferation of DU145 cells cultured in adherent conditions
To characterise further the effects of GA and M3MOG, their ability to reduce the proliferation of DU145 cells grown as adherent monolayer was evaluated. Adherent cells were treated with different concentrations of GA and M3MOG, and the cellular proliferation was examined (Fig. 5). The proliferation of DU145 cells grown in adherent condition was impaired with greater efficiency by GA (IC50 1.710 μM, Fig. 5A) than by M3OMG (IC50 > 10 μM, Fig. 5B). These results showed that GA was a more potent inhibitor of cellular proliferation than M3OMG on the bulk tumour cell population in adherent culture conditions. | ||
| Fig. 5 Effect of GA (A, black bars) and M3MOG (B, empty bars), tested at different concentrations (ranging from 0 to 10 μM), on the proliferation of DU145 cells cultured in adherent conditions. Data are presented as percentage of untreated control cells. | ||
Overall, the results has shown that when GA was applied to cells cultured in differentiation permissive conditions, the proliferation of prostate cancer cells was strongly impaired. Interestingly, M3OMG was less effective in preventing proliferation of adherent growing cells.
2.6 Effect of GA and M3MOG on the clonogenic capacity of DU145 cells
Another measure of the proliferative capacity of cancer cells is given by their ability to grow in vitro at low cell density and to generate single cell derived colonies. To address the ability of GA and M3MOG to affect the clonogenic capacity, DU145 cells were plated at low cell density. GA and M3MOG were added to the media at different concentrations and, after 10 days, cell colonies were counted. GA was a potent inhibitor of clonogenicity, with a half maximal effective concentration (EC50) value of 0.852 μM (Fig. 6). In the same experimental conditions, M3OMG inhibited the clonogenic capacity of DU145 cells at higher concentrations than GA, with a EC50 value of 5.817 μM (Fig. 6).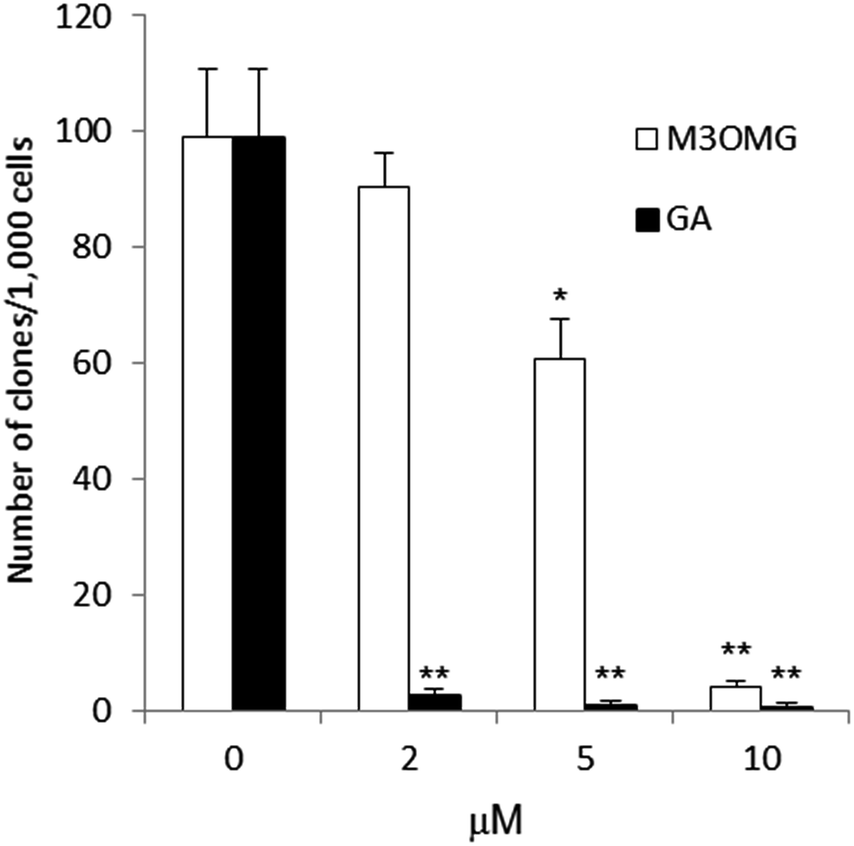 | ||
| Fig. 6 Clonogenic ability of DU145 cells, plated at low density in the presence of GA (A, black bars) and M3OMG (B, empty bars) at the concentrations of 2, 5, and 10 μM after 10 days. The initial cell density was 1000 cells per well in 6-wells plates. | ||
The results reported so far has shown that when GA and M3OMG were tested on cells plated at clonal density, GA decreased the clonogenic capacity of DU145 cells more efficiently than M3OMG. Thus, the activity of M3OMG seems to be more selective for cells with stem-like properties than for differentiated cancer cells.
2.7 Determination of GA and M3OMG interactions with cells and culture-medium components
The differences observed in the activity of GA and M3OMG in DU145 cells, cultured as adherent monolayer or as prostatospheres, suggested further evaluation of the interaction between the two compounds and cell culture-medium components, in presence or absence of FBS and cells. Thus, we determined the amount of free GA and M3OMG in cell culture medium, cell culture medium supplemented with FBS (10%), and in presence of cells. Then we compared these results with free GA and M3OMG aqueous solutions at the same concentration. In all these conditions, the concentration of free GA was lower than the limit of detection, indicating complete interaction between GA and the constituents of the cell culture medium. These results could be justified with the high reactivity of the carboxyl group and high affinity with culture-medium constituents. The concentration of free M3OMG in cell culture medium and medium supplemented with FBS was reduced by 53.5 ± 0.5% and 51.0 ± 0.2%, respectively. By contrast, M3OMG showed a complete cellular absorption, since its concentration in the culture medium in presence of cells was lower than detection limit. These data suggests that M3OMG has less affinity with culture-medium constituents than GA, probably due to its ester functional group, and was completely absorbed by the cells.Overall, these results indicated that GA showed higher affinity for culture-medium components than M3OMG, which remained in higher concentration as free form in cell culture medium, perhaps allowing increased cellular uptake. Growing evidence indicates that the CSC-enriched subpopulations, from human tumours and cell lines, exhibit a drug-resistant phenotype compared with the adherent counterparts. Thus, the greater efficacy of M3OMG against prostatospheres might be due to the higher concentration of free M3OMG in cell culture medium and to higher cell absorption compared to GA.
3. Experimental
3.1 Reagents
Methyl gallate, borax, dimethyl sulphate, sodium hydroxide, sulphuric acid, chloroform, HPLC-grade water, methanol, ethanol 96%, formic acid, agarose, PBS, EDTA, poly-hema, tri-chloroacetic acid, sulforhodamine B (SRB), acetic acid, Tris base, RPMI 1640 medium, FBS (fetal bovine serum), poly(2-hydroxyethylmethacrylate), insulin, FGF2, EGF, β-actin, and crystal violet were purchased from Sigma-Aldrich. Mammary Epithelial Basal Medium (MEBM) was purchased from Lonza. 1xB-27 Supplement, Pen/Strep, and Superscript III One-Step RT-PCR Kit were purchased from Invitrogen. Millex-HV PVDF membranes were purchased from Merck Millipore. M3OMG was synthetized as reported in Curti et al. 2014.83.2 Cell cultures and culture conditions
DU145 human prostate cancer cell line was maintained in RPMI 1640 medium as described by Albino et al.23 Conditional media were generated by incubating exponentially growing cells in RPM1 1640 medium, supplemented with FBS (10%). Prostatosphere cultures of DU145 cells were established as previously described.23,24 In brief, single cells were plated within flasks coated with poly-hema (at a density of 10![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 000 viable cells per mL) in MEBM serum-free medium, containing 1xB-27 supplement, 4 μg mL−1 insulin, 20 ng mL−1 FGF2, 10 ng mL−1 EGF, and 5% Pen/Strep. The resulting spheres were collected after 7–10 days by centrifugation (800 rpm), dissociated using PBS, supplemented with 2 mM EDTA, and finally replated into the poly-hema coated culture flasks. To guarantee plating of viable single cells, the dissociated cells were coated with Trypan blue. To assess sphere formation over time, the cells were seeded at the density of 1000 cells per mL, in the presence of different concentrations of GA or M3OMG. After 7 days, the spheres were counted using an optical microscope. To assess the effect on the expression levels of target genes, cells were seeded for 7–10 days to generate prostatospheres and subsequently treated with GA or M3OMG. After 16 hours, prostatospheres were collected and total RNA was extracted.
000 viable cells per mL) in MEBM serum-free medium, containing 1xB-27 supplement, 4 μg mL−1 insulin, 20 ng mL−1 FGF2, 10 ng mL−1 EGF, and 5% Pen/Strep. The resulting spheres were collected after 7–10 days by centrifugation (800 rpm), dissociated using PBS, supplemented with 2 mM EDTA, and finally replated into the poly-hema coated culture flasks. To guarantee plating of viable single cells, the dissociated cells were coated with Trypan blue. To assess sphere formation over time, the cells were seeded at the density of 1000 cells per mL, in the presence of different concentrations of GA or M3OMG. After 7 days, the spheres were counted using an optical microscope. To assess the effect on the expression levels of target genes, cells were seeded for 7–10 days to generate prostatospheres and subsequently treated with GA or M3OMG. After 16 hours, prostatospheres were collected and total RNA was extracted.
3.3 RNA extraction and qRT-PCR
Total RNA was extracted with SV Total RNA Isolation System (Promega). Quantitative real-time RT-PCR (qRT-PCR) was performed using the ABI 7000 sequence detection system and SYBR Green chemistry. Forward (_f) and reverse (_r) primers for the NF-κB target genes IL-6, COX-2 and ESE-1 were custom-made and their sequences are reported in Table 1. Total RNA (100 ng) was reverse-transcribed and amplified using Superscript III One-Step RT-PCR Kit. β-Actin was used as control of the input RNA level. Verification of specific product amplification was determined by dissociation curve analysis. Data were analyzed by StepOne software v2.0 (Applied Biosystems) and reported as RQ with respect to the appropriate calibrator sample using the 2-ΔΔCt method.23| Gene/region | Sequence 5′ to 3′ | Primer |
|---|---|---|
| ACTIN | GGATGCCACAGGACTCCAT | ACTIN_f |
| ACTIN | ATTGGCAATGAGCGGTTC | ACTIN_f |
| IL-6 | GTACATCCTCGACGGCATCTC | IL-6_f |
| IL-6 | GCTGCTTTCACACATGTTACTCTTG | IL-6_f |
| ESE-1 | GGTACTGACCCTGAGCAACC | ESE-1_f |
| ESE-1 | AGTCCAGAACCTGCGTCTTC | ESE-1_f |
| COX-2 | GCAATAACGTGAAGGGCTGT | COX-2_f |
| COX-2 | CGGGAAGAACTTGCATTGAT | COX-2_f |
3.4 Colorimetric cell proliferation assay
Cellular proliferation was evaluated with SRB assay. Cells were plated at a density of 1000 cells per well in 96-wells plate. After 24 hours, cells were treated with GA and M3OMG at different concentrations. After 72 hours, cells were fixed with 10% (w/v) trichloroacetic acid and stained for 30 minutes with SRB solution (0.057% in acetic acid 1%). The excess dye was removed by washing repeatedly with 1% (v/v) acetic acid. The protein-bound dye was then dissolved in 10 mM Tris base solution (pH 10.5), and optical density (OD) was determined at 570 nm using a microplate reader.3.5 Clonogenic assay
For clonogenic assay, cells were plated at a density of 1000 cells per well in a 6-wells plate. After 24 hours, cells were treated with GA and M3OMG at different concentrations. After 10 days, the colonies were fixed and stained with crystal violet 1% in 20% of ethanol and counted under an optical microscope.3.6 Determination of GA and M3OMG interactions with culture-medium components and cells
To evaluate the potential interaction between GA and M3OMG and culture-medium components and/or cells, GA and M3OMG solutions (5 μM) were prepared using different solvents: (i) H2O/MeOH: 50/50 (% v/v); (ii) serum-free cell culture medium; (iii) cell culture medium including FBS; (iv) cell culture medium including cells. Each solution was filtered through 0.45 μm and 0.20 μm PVDF membranes (Millex – HV) and injected in triplicate into a RP-HPLC-DAD system. All experiments were performed using an Agilent 1100 HPLC system (Agilent, Waldbronn, Germany) equipped with a gradient quaternary pump and a diode array detector. The Agilent Chemstation software was used for control of the HPLC system and data processing. Chromatographic separation was carried out on a Zorbax Eclipse XDB-C18 analytical column (150 × 4.6 mm; 5 μm; Agilent) coupled with a Hypersil Gold RP18 guard column (10 × 2.1 mm, 5 μm, Agilent), both maintained at 25 °C. Injection volume was 10 μL and autosampler temperature was set at 4 °C. The mobile phase was a mixture of deionized water containing the 0.1% of formic acid and methanol. Two different gradient programs were used. The first method, used for the determination of GA, consisted of methanol concentration: 0 min, 1%; 0–30 min 15%; 30–42 min, 40%; 42–48 min, 60%; 48–55 min, 90%; 55–60 min, 100%; 60–70 min, 100%; 70–80 min, 1%; 80–85 min, 99%. The second method, used for the determination of M3OMG, consisted of methanol concentration: 0 min, 1%; 30–42 min, 40%; 42–48 min, 60%; 48–55 min, 90%; 55–60 min, 100%; 60–70 min, 100%; 70–80 min, 1%; 80–85 min, 99%). The mobile phase flow rate was 0.3 mL min−1. GA and M3OMG were identified by comparing their retention times and UV spectra with those of standard compounds.3.7 Statistical analysis
Differences between groups were assessed with an unpaired two-tailed t-test and were considered statistically significant for P < 0.05 (*) and P < 0.01 (**).4. Conclusions
In conclusion, this study reveals the different ability of GA and M3OMG to inhibit the NF-κB activity and the growth of PCa cells with stem-like properties. The underlined differences may be related to the differential impact of NF-κB inhibition on the two cell populations (i.e. more differentiated bulk and less differentiated stem-like cancer cells). Additionally, the different ability of GA and M3OMG to selectively interfere with NF-κB independent processes may affect the preferential cytotoxic activity of the two compounds for differentiated proliferating cancer cells or for less differentiated CSC-like cells. In this regard, M3OMG might be more selective in targeting the NF-κB pathway and consequently more effective against CSC-like cells, which might depend more on the activation of the pathway. GA might have more promiscuous effects interfering with other cellular targets in addition to the NF-κB pathway, resulting in increased cytotoxicity against the more differentiated proliferating cancer cells. Therefore, our results show that both GA and M3OMG affect pathways and biological processes associated with the CSC phenotype and tumour progression, supporting further investigation on the potential use of these compounds for the prevention and treatment of prostate cancer.Acknowledgements
We thank the EPSRC UK National Mass Spectrometry Facility at Swansea University for their support in accurate mass analysis of our test compounds.References
- M. Daglia, A. Di Lorenzo, S. F. Nabavi, Z. S. Talas and S. M. Nabavi, Curr. Pharm. Biotechnol., 2104, 15, 362–372 CrossRef.
- M. Daglia, R. Antiochia, A. P. Sobolev and L. Mannina, Food Res. Int., 2014, 63, 275–289 CrossRef CAS PubMed.
- S. Habtemariam, J. Med. Food, 2011, 14, 1412–1418 CrossRef CAS PubMed.
- C. L. Hsu and G. C. Yen, Br. J. Nutr., 2007, 98, 727–735 CrossRef CAS PubMed.
- S. M. Nabavi, S. Habtemariam, S. F. Nabavi, A. Sureda, M. Daglia, A. H. Moghaddam and M. A. Amani, Mol. Cell. Biochem., 2013, 372, 233–239 CrossRef CAS PubMed.
- S. F. Nabavi, S. M. Nabavi, S. Habtemariam, A. H. Moghaddam, M. Daglia and F. Abolhasani, Fluoride, 2012, 45, 262–268 Search PubMed.
- S. F. Nabavi, S. M. Nabavi, S. Habtemariam, A. M. Moghaddam, A. Sureda and M. Mirzaei, Cell. Mol. Neurobiol., 2013, 33, 261–267 CrossRef CAS PubMed.
- V. Curti, E. Capelli, F. Boschi, S. F. Nabavi, A. I. Bongiorno, S. Habtemariam, S. M. Nabavi and M. Daglia, Mol. Nutr. Food Res., 2014, 58, 1776–1784 CAS.
- A. Abdelwahed, I. Bouhlel and I. Skandrani, Chem.-Biol. Interact., 2007, 165, 1–13 CrossRef CAS PubMed.
- C. Hsu, W. Lo and G. Yen, J. Agric. Food Chem., 2007, 55, 7359–7365 CrossRef CAS PubMed.
- D. M. Parkin and C. S. Muir, IARC Sci. Publ., 1992, 45–173 CAS.
- H. G. Sim and C. W. Cheng, Eur. J. Cancer, 2005, 41, 834–845 CrossRef PubMed.
- L. Reddy, B. Odhav and K. D. Bhoola, Pharmacol. Ther., 2003, 99, 1–13 CrossRef CAS.
- H. Vainio and E. Weiderpass, Nutr. Cancer, 2006, 54, 111–142 CrossRef CAS PubMed.
- B. B. Aggarwal and P. Gehlot, Curr. Opin. Pharmacol., 2009, 9, 351–369 CrossRef CAS PubMed.
- N. D. Perkins, Nat. Rev. Cancer, 2012, 12, 121–132 CAS.
- N. Longoni, M. Sarti, D. Albino, G. Civenni, A. Malek, E. Ortelli, S. Pinton, M. Mello-Grand, P. Ostano, G. D'Ambrosio, F. Sessa, R. Garcia-Escudero, G. N. Thalmann, G. Chiorino, C. V. Catapano and G. M. Carbone, Cancer Res., 2013, 73, 4533–4547 CrossRef CAS PubMed.
- H. H. Tseng, P. N. Chen, W. H. Kuo, J. W. Wang, S. C. Chu and Y. S. Hsieh, Integr. Cancer Ther., 2012, 11, 267–278 CrossRef CAS PubMed.
- T. C. Reddy, D. B. Reddy, A. Aparna, K. M. Arunasree, G. Gupta, C. Achari, G. V. Reddy, V. Lakshmipathi, A. Subramanyam and P. Reddanna, Toxicol. In Vitro, 2012, 26, 396–405 CrossRef PubMed.
- C. Hsiang, Y. Hseu, Y. Chang, K. J. S. Kumar, T. Ho and H. Yang, Food Chem., 2013, 136, 426–434 CrossRef CAS PubMed.
- J. Marx, Cancer Res., 2003, 301, 1308–1310 CAS.
- S. K. Singh, C. Hawkins, I. D. Clarke, J. A. Squire, J. Bayani, T. Hide, R. M. Henkelman, M. D. Cusimano and P. B. Dirks, Nature, 2004, 432, 396–401 CrossRef CAS PubMed.
- D. Albino, N. Longoni, L. Curti, M. Mello-Grand, S. Pinton and G. Civenni, et al., Cancer Res., 2012, 72, 2889–2900 CrossRef CAS PubMed.
- G. Civenni, A. Malek, D. Albino, R. Garcia-Escudero, S. Napoli, S. Di Marco, S. Pinton, M. Sarti, G. M. Carbone and C. V. Catapano, Cancer Res., 2013, 73, 6816–6827 CrossRef CAS PubMed.
- V. K. Rajasekhar, L. Studer, W. Gerald, N. D. Socci and H. I. Scher, Nat. Commun., 2010, 2, 162 CrossRef PubMed.
- G. Civenni, P. Costales, C. García-Inclán, D. Albino, N. Longoni, L. E. Nuñez, F. Moris, G. M. Carbone and C. V. Catapano, Eur. J. Cancer, 2014, 50, 159 CrossRef.
Footnotes |
| † These two authors share first authorship. |
| ‡ These two authors share the senior authorship. |
| This journal is © The Royal Society of Chemistry 2015 |