
The screening of microalgae mutant strain Scenedesmus sp. Z-4 with a rich lipid content obtained by 60Co γ-ray mutation†
Bingfeng Liu‡
*,
Chao Ma‡,
Ruonan Xiao,
Defeng Xing,
Hongyu Ren and
Nanqi Ren
State Key Laboratory of Urban Water Resource and Environment, School of Municipal and Environmental Engineering, Harbin Institute of Technology, P.O. Box 2614, 202 Haihe Road, Harbin 150090, China. E-mail: lbf@hit.edu.cn; Fax: +86 451 86282008; Tel: +86 451 86282110
First published on 2nd June 2015
Abstract
In this study, a microalgae mutant Scenedesmus sp. Z-4 with a lipid content of 28.86% and biomass of 2.876 g L−1 was obtained using 60Co γ-ray mutation. The lipid productivity (138 mg L−1 per day) and content of mutant Z-4 were enhanced by 113% and 71.3% compared to that of the wild strain, respectively. In addition, unlike the wild strain, the microalgae cells were larger and the surface was rougher in mutant Z-4.
Energy shortages and environmental pollution caused by continuous dependence on fossil fuels have attracted worldwide attention.1 Therefore, the development of clean, renewable green fuels is extremely urgent. Currently, biodiesel is becoming well-known as an environmentally-friendly biofuel due to its non-toxic and biodegradable characteristics, and it will be a potential substitute for fossil fuels in the future.2 So far, the major feedstocks for biodiesel production are from high-oil crops and animal fats.3 However, the widespread use of crops containing oil will occupy a large amount of arable land and this will lead to competition between fuel and food, and the sustainability of animal fats did not meet the demands of the rapid development of biodiesel production.4 So, the selection of raw materials is critical for biodiesel production.
Recently, microalgae as a new raw material have become a focus of biodiesel production because of the use of carbon dioxide, higher photosynthetic efficiency, rapid growth rate, high lipid content, etc.5 Although biodiesel production from microalgae is considered as a promising and efficient technique, there are still some barriers limiting its practical application. One of the key limiting factors is the high production cost compared to that of traditional fossil fuels.6 The obtaining of microalgae strains with high lipid productivity using an efficient screening method can make a contribution to resolving the above problem and help accelerate the pace of industrial production of microalgal biodiesel. This also is one of the major challenges in the algal biofuels production chain.7 However, screening microalgae with high biomass and lipid content directly from the natural environment is difficult. The application of mutagenesis technology shortens the screening period and improves the screening efficiency, making it possible to find more microalgae strains with an excellent lipid content.8
Compared to selective breeding and genetic engineering, mutation breeding proves to be a more mature and effective method for microalgae screening.9 Currently, ultraviolet radiation is widely used in induced mutation of microalgae, but its intensity and time of radiation are hard to control, in addition to the unstable characteristics after mutation. Therefore, it is vital to establish an effective and stable mutation system. In contrast to ultraviolet radiation, 60Co γ-rays have a higher energy density, and are widely available for the breeding of agricultural crops.10,11 A few studies have reported the effect of gamma ray radiation on Escherichia coli.12,13 However, information about the application of 60Co γ-rays in the screening of microalgae strains containing a rich lipid content is extremely limited.
So, the objective of this study is to screen a lipid-rich microalgae mutant using 60Co γ-ray irradiation after two rounds of mutation and determination of the optimal dosage of the 60Co γ-rays. The morphology, physiological and biochemical characteristics of the microalgal mutant are investigated.
The wild microalgae strain MC-1 is from one of 88 microalgal colonies, which were isolated using an improved Nile red staining method, employed in this study. After optimization of lipid production according to previously described parameters,14 the lipid content of strain MC-1 was 15.7%. The observations using a microscope and SEM showed that the cells of strain MC-1 were green and oblong, and the dimensions of the cells approximately ranged from 4 to 14 μm in length and 2 to 8 μm in width, and that 2–4 microalgal cells usually flocked together along the width direction (Fig. 1f), which are the typical characteristics of the genus Scenedesmus. In addition, the microalgae strain MC-1 with Nile red staining exhibited a weak yellow fluorescence intensity, this also indicated that a low lipid yield was present in the cells of strain MC-1 (Fig. 1e).
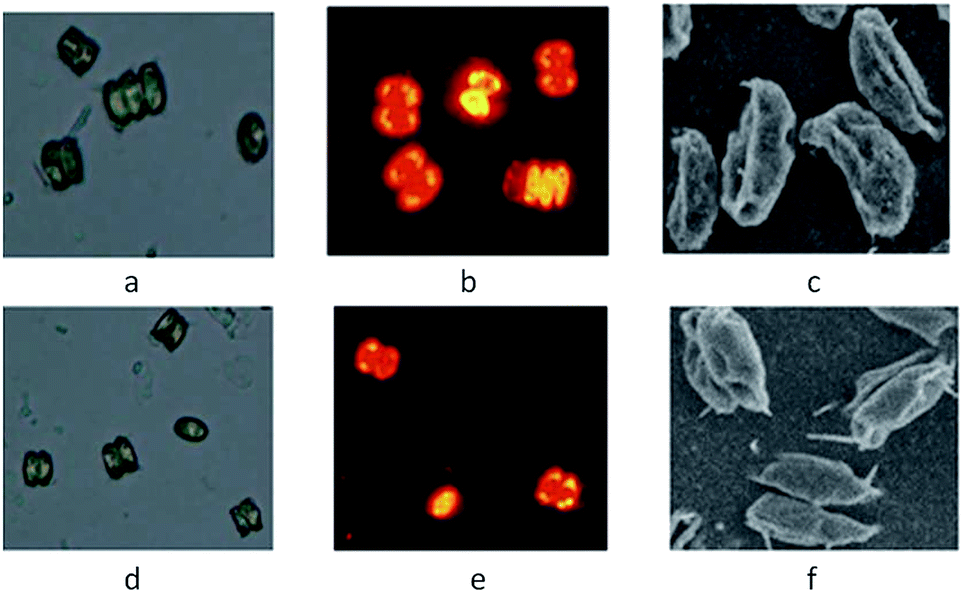 | ||
| Fig. 1 Microscopic morphological observations and SEM images of mutant Z-4 (a–c) and strain MC-1 (d–f). (a) Light micrograph; (b) fluorescence micrograph of the microalgae stained with Nile red; (c) SEM image (5000×); (d) light micrograph; (e) fluorescence micrograph of the microalgae stained with Nile red; (f) SEM image (5000×). | ||
The 18S rDNA complete sequence of strain MC-1 consisting of 1292 bp was determined and submitted to the GenBank (accession no. KJ740712). The 18S rDNA complete sequence of strain MC-1 exhibited 100% similarity to those of Scenedesmus sp. R-16 (accession no. KC859922)14 and Scenedesmus abundans UTEX 343 (Accession no. X73995.1), and the nucleotides of these two microalgae strains (1419 bp and 1794 bp, respectively) covered all nucleotides of strain MC-1 (1292 bp). So, the microalgae strain MC-1 was designated as a new strain of Scenedesmus sp. and it was used as the original microalgae for further mutation by 60Co γ-ray irradiation.
Using the appropriate dosage range of 60Co γ-rays was vitally important for the microalgal mutation and determined the microalgal fatalities. In this work, initial dosages of 0, 50, 100, 500, 1000, 2000 and 3000 Gy were chosen to explore the effect of different dosages of 60Co γ-rays on the microalgal fatalities and lipid productivity. The results showed that microalgal colonies with great differences in the numbers grown on BG-11 agar plates were obtained after the wild strain MC-1 was irradiated using the above dosages. No colonies formed on the agar plates when the dosage was in the range of 1000 to 3000 Gy, suggesting that the high dosages had a severely lethal effect on the cells of the strain. Rays at dose rates exceeding a certain threshold might cause serious damage to the organism DNA and lead to cell dead.12 In addition, only a few microalgal colonies were formed on the plate with the dosage of 500 Gy. Compared to the control, the numbers of colonies were less, but the microalgal colonies were slightly larger under 50 and 100 Gy radiation. This indicated that the low dosages may have a positive effect on the microalgal cell growth. The fatalities of the microalgal colonies for the dosages of 50 and 100 Gy were approximately 5% and 60%, respectively. This reached 98% for 500 Gy. Therefore, a dosage range of 0 to 500 Gy was applied for the second mutation using 60Co γ-rays to further enhance the lipid production yield.
The colonies from different agar plates were inoculated into 50 mL Erlenmeyer flasks containing 30 mL BG-11 medium to screen the microalgae strains with a rich lipid content. Compared to the control, the lipid content was improved to some extent after the 60Co γ-ray mutation when the low dosages of 50 and 100 Gy were used. Among the 18 tested microalgae strains (Fig. 2), the strain numbered as 50 Gy-2 (it was named Y-2) was determined as the best strain because of its high lipid content of 21.80% and biomass of 2.667 g L−1, which were higher than the lipid content (16.84%) and biomass (2.311 g L−1) of the control. The lipid productivity was enhanced by 49.3% from 64.8 mg L−1 per day for the wild strain MC-1 to 97 mg L−1 per day for the mutant Y-2. Thus, the mutant Y-2 was selected as the original strain for the second round mutation. The mutant Y-2 was irradiated using 60Co γ-rays at the dosage range from 0 to 500 Gy, and the dosages were 0, 50, 100, 200, 300, 400 and 500 Gy, respectively. Unlike the results from the first mutation, the fatality levels of this test were 34.2%, 76.5%, 92.2% and 99.8% corresponding to a dosage of 50, 100, 200 and 300 Gy, respectively. It is worth noting that mutant strain Y-2 could only survive at the low dosage conditions (50–300 Gy), indicating that it was more sensitive to high dosages of 60Co γ-rays in the second mutation. It may be concluded that the tolerance of mutant Y-2 to 60Co γ-rays decreased because of DNA and protein damage to mutant Y-2 in the above first mutation. Compared to mutant Y-2, the colonies were larger at the dosage of 50 Gy and 100 Gy, which is similar to the results of the first mutation. After the second mutation, a mutant strain Z-4 (100 Gy-4) with a lipid content of 28.86%, biomass of 2.876 g L−1 and lipid productivity of 138 mg L−1 per day was obtained (Fig. 3). The lipid content, biomass and lipid productivity were enhanced by 28.85%, 18.76% and 47.8% respectively compared to that of mutant Y-2 (21.80%, 2.667 g L−1 and 93.4 mg L−1 per day). Finally, the lipid content and productivity of mutant Z-4 were obviously enhanced by 71.3% and 113% compared to the wild strain MC-1. The lipid production yield and cell growth of the mutant can be promoted by γ-ray irradiation.15 The genetic stability of mutant Z-4 was also analyzed by obtaining subcultures many times, and the lipid content of mutant Z-4 was maintained at 29%, while that of strain MC-1 was 17%, respectively, suggesting that the mutant Z-4 is an ideal lipid-rich microalgal mutant and the reasons for the rapid growth and high lipid productivity of mutant Z-4 will be studied in future work.
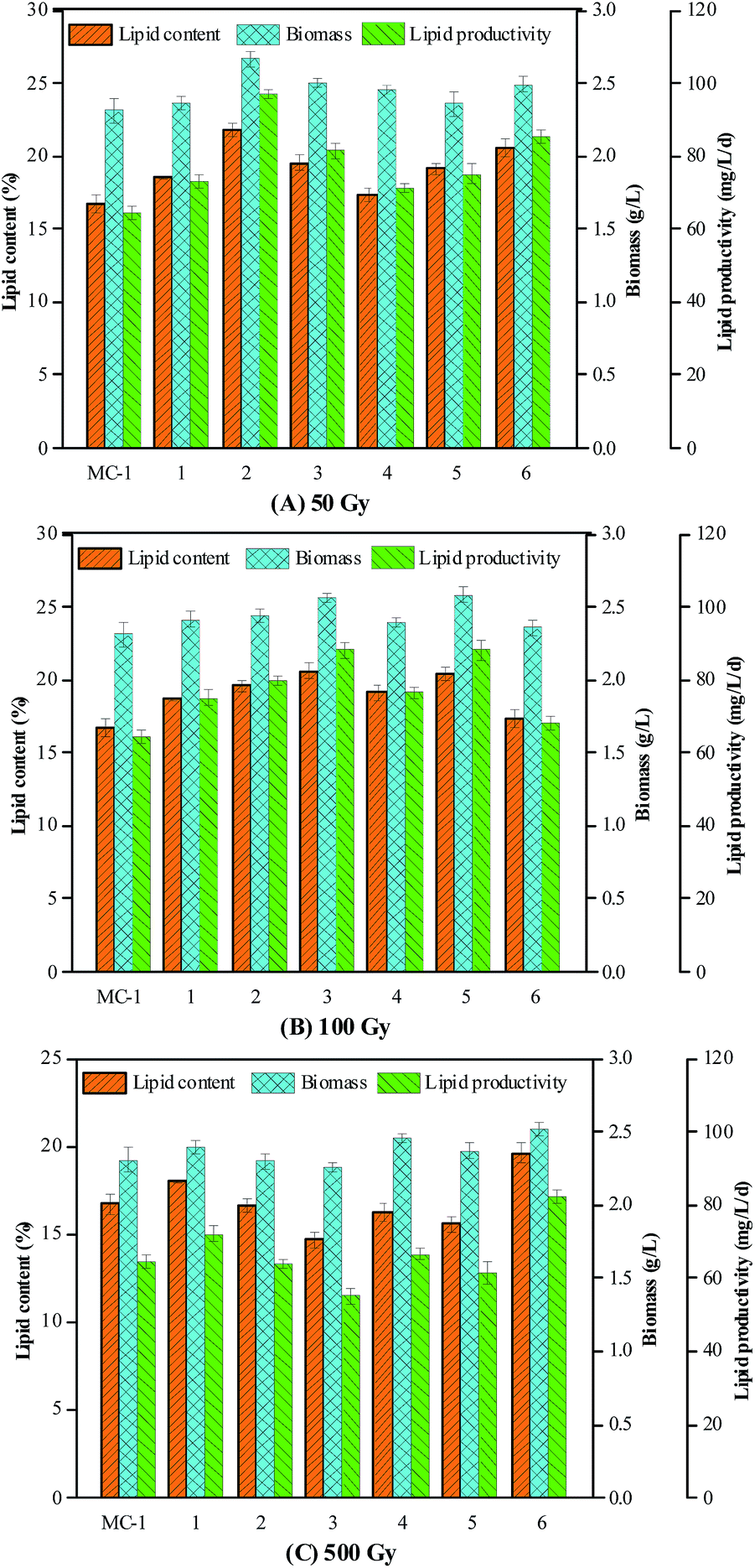 | ||
| Fig. 2 The lipid content of the selected 18 strains used in the first mutation under different dosages (m Gy-n: m is the value of the dosage and n is the number of the strain). | ||
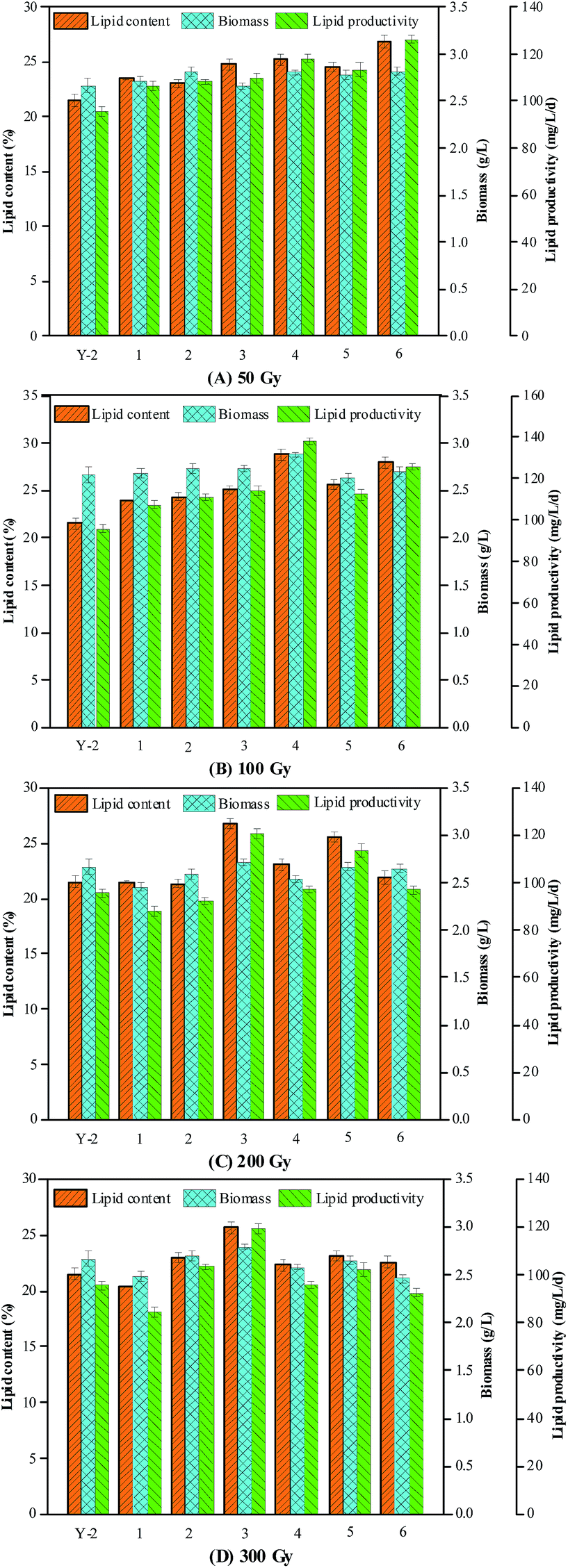 | ||
| Fig. 3 The lipid content of the selected 24 strains used in the second mutation under different dosages (m Gy-n: m is the value of the dosage and n is the number of the strain). | ||
After two mutations, mutant Z-4 with high content and productivity of lipid was screened. This result indicates that using 60Co γ-rays is a useful and efficient manner of obtaining a microalgal mutant strain containing a rich lipid content. The microorganism was exposed to radiation that can induce the damage of cellular DNA.17 Excessive radiation intensity (high dosage of 500 to 3000 Gy) had a strong lethal effect on the microalgal cells. In this case, the degree of cellular DNA damage was huge, which caused cell death. So no colonies grew on the agar plates and the fatality level was 100%. However, an appropriate radiation intensity can maintain normal cell growth, even some excellent traits were found. In this study, the lipid production ability was remarkably improved by two mutations using 60Co γ-rays. This indicated that the capacity for lipid biosynthesis of the microalgae increased and it is suspected that some key genes related to lipid biosynthesis were mutated which led to positive expression, for example, the gene expression of acetyl-CoA carboxylase (ACCase) increased five fold after mutagenesis.15 Our further work will focus on the relationship between gene expression and lipid biosynthesis in microalgae by using DNA recombination methods to determine their functions.
The 18S rDNA complete sequence (1292 bp) of mutant Z-4 was absolutely the same as that of strain MC-1, indicating that the species characteristics did not change after mutation. So, the microalgal mutant of wild strain MC-1was named Scenedesmus sp. mutant Z-4.
The morphological features of the mutant strain Z-4 were observed. The formed colonies are green and round, and had diameters of 1 to 3 mm, and the surface and edge of the colony are smooth and neat, respectively. The growth in the BG-11 medium was in a green suspension and the biomass would sink to the bottom after a period of standing. In addition, the color of the microalgal cells turned from yellow-green to dark green as they grew. The photographs of the mutant strain Z-4 by optical microscopy and SEM are shown in Fig. 1a–c, respectively. According to the observations, the characteristics of the mutant Z-4 were similar to that of the wild strain MC-1. The main distinction was found to be that the microalgal cells (4–15 μm long and 2–8 μm wide) of mutant Z-4 were larger than that of strain MC-1 (Fig. 1), which may explain the reason why the mutant Z-4 could accumulate more biomass and lipid. However, in contrast to our results, after mutation the size of the cells of mutant decreased.15,16 This also indicated that different strains, species or mutation conditions caused obvious differences in microalga traits. Furthermore, the surface of the mutant strain Z-4 in the SEM observation was rougher, which probably resulted from changed surface features by 60Co γ-radiation (Fig. 1c). This may enhance their environmental adaptation, such as tolerating high salinity, high CO2, high temperature, nitrogen depletion, and so on.
Several physiological and biochemical characteristics, including substrate utilization, optimal temperature and optimal pH, are listed in Table 1. The optimal carbon source was glucose for the strain Z-4, and fructose, sucrose, maltose and sodium acetate were also appropriate substrates. However, the growth of the strain Z-4 was not satisfactory in BG-11 medium complemented with sodium butyrate, sodium propionate and sodium malate, and sodium bicarbonate, citric acid, xylose and alcohol could not even be utilized. The strain Z-4 could grow in a wide range of temperature (15–35 °C) and pH (4.0–12.0), and the optimal temperature and pH were 25 °C and 7.0, respectively.
| Characteristic | Mutant Z-4 |
|---|---|
| Glucose | ++ |
| Fructose | + |
| Sucrose | + |
| Maltose | + |
| Xylose | − |
| Sodium acetate | + |
| Propionate | ± |
| Butyrate | ± |
| Malate | − |
| Bicarbonate | − |
| Citric acid | − |
| Alcohol | − |
| Temperature range | 15–35 °C |
| Optimal temperature | 25 °C |
| pH range | 4.0–12.0 |
| Optimal pH | 7.0 |
| Nutrition type | Photoheterotrophic |
Conclusions
A microalgae mutant strain containing a rich lipid content was screened successfully using a 60Co γ-ray mutation method. After a second mutation, a mutant Z-4 with high lipid content and productivity was obtained. The productivity (138 mg L−1 per day) and content (28.84%) of lipid for mutant Z-4 were increased by 113% and 71.3% compared with that of wild strain MC-1. Thus, mutant strain Z-4 has great potential as an ideal raw material for biodiesel. This study also demonstrated that using 60Co γ-ray mutation technology is an efficient and useful way to obtain excellent microalgae strains.Acknowledgements
This study is supported by the National Natural Science Foundation of China (no. 51478139), Fundamental Research Funds for Central Universities (no. HIT.BRETIII.201418), State Key Laboratory of Urban Water Resource and Environment (Harbin Institute of Technology) (no. 2015TS02) and Harbin Innovation Talents Funding of Science and Technology (no. 2014RFQXJ084).Notes and references
- J. Liu, L. Chen, J. Wang, J. Qiao and W. Zhang, Proteomic analysis reveals resistance mechanism against biofuel hexane in Synechocystis sp. PCC 6803, Biotechnol. Biofuels, 2012, 5, 68 CrossRef CAS PubMed
.
- A. L. Ahmad, N. H. M. Yasin, C. J. C. Derek and J. K. Lim, Microalgae as a sustainable energy source for biodiesel production: A review, Renewable Sustainable Energy Rev., 2011, 15(1), 584–593 CrossRef CAS PubMed
- X. D. Deng, Y. J. Zhou, Y. J. Li and X. W. Fei, Optimization of the culture conditions of a Chlamydomonas high oil content ultraviolet mutant CC124-M25 and polymorphism analysis by inter-simple sequence repeat (ISSR), Afr. J. Microbiol. Res., 2012, 6(15), 3604–3619 CAS
- T. M. Mata, A. A. Martins and N. S. Caetano, Microalgae for biodiesel production and other applications, Renewable Sustainable Energy Rev., 2010, 14, 221–232 CrossRef PubMed
- L. Gouveia and A. C. Oliveira, Microalgae as a raw material for biofuels production, J. Ind. Microbiol. Biotechnol., 2009, 36, 269–274 CrossRef CAS PubMed
- F. Yang, M. Hanna and R. Sun, Value-added uses for crude glycerol–a byproduct of biodiesel production, Biotechnol. Biofuels, 2012, 5, 13 CrossRef CAS PubMed
- M. Hannon, J. Gimpel, M. Tran, B. Rasala and S. Mayfield, Biofuels from algae: challenges and potential, Biofuels., 2010, 1(5), 763–784 CrossRef CAS
- G. R. Hu, Y. Fan, L. Zhang, C. Yuan, J. F. Wang, W. J. Li, Q. Hu and F. L. Li, Enhanced lipid productivity and photosynthesis efficiency in a Desmodesmus sp. mutant induced by heavy carbon ions, PLoS One, 2013, 8(4), e60700 CAS
- K. Manandhar-Shrestha and M. Hildebrand, Development of flow cytometric procedures for the efficient isolation of improved lipid accumulation mutants in a Chlorella sp., Microalga. J. Appl. Phycology., 2013, 25(6), 1643–1651 CrossRef CAS PubMed
- L. Zhao, L. Liu, J. Wang, H. Guo, J. Gu, S. Zhao, J. Li and Y. Xie, Development of a new wheat germplasm with high anther culture ability by using a combination of gamma-ray irradiation and anther culture, J. Sci. Food Agric., 2015, 95(1), 120–125 CrossRef CAS PubMed
- J. E. Hwang, J. W. Ahn, S. J. Kwon, J. B. Kim, S. H. Kim, S. Y. Kang and D. S. Kim, Selection and molecular characterization of a high tocopherol accumulation rice mutant line induced by gamma irradiation, Mol. Biol. Rep., 2014, 41(11), 7671–7681 CrossRef CAS PubMed
- J. Min, C. W. Lee and M. B. Gu, Gamma-radiation dose-rate effects on DNA damage and toxicity in bacterial cells, Radiat. Environ. Biophys., 2003, 42, 189–192 CrossRef PubMed
- T. Atsumi, E. Fujimoto, M. Furuta and M. Kato, Effect of gamma-ray irradiation on Escherichia coli motility, Cent. Eur. J. Biol., 2014, 9(10), 909–914 CrossRef CAS
- H. Y. Ren, B. F. Liu, C. Ma, L. Zhao and N. Q. Ren, A new lipid-rich microalga Scenedesmus sp. strain R-16 isolated using Nile red staining: effects of carbon and nitrogen sources and initial pH on the biomass and lipid production, Biotechnol. Biofuels, 2013, 6, 143 CrossRef CAS PubMed
- J. Cheng, J. Feng, R. L. Cheng, C. X. Zhang, J. H. Zhou and K. F. Cen, Gene expression and metabolic pathways related to cell growth and lipid synthesis in diatom Nitzschia ZJU2 after two rounds of mutagenesis by γ-rays, RSC Adv., 2014, 4, 28463 RSC
- S. G. Lynn, S. S. Kilham, D. A. Kreeger and S. J. Interlandi, Effect of nutrient availability on the biochemical and elemental stoichiometry in the freshwater diatom Stephanodiscus minutulus (bacillariophyceae), J. Phycol., 2000, 36, 510–522 CrossRef CAS
- J. Cadet, T. Douki, D. Gasparutto and J. L. Ravanat, Radiation-induced damage to cellular DNA measurement and biological role, Radiat. Phys. Chem., 2005, 72, 293–299 CrossRef CAS PubMed
Footnotes |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/c5ra07263h |
| ‡ These authors contributed to the work equally. |
| This journal is © The Royal Society of Chemistry 2015 |