
Violet pigment production from liquid pineapple waste by Chromobacterium violaceum UTM5 and evaluation of its bioactivity
Abstract
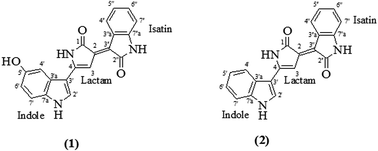
Synthetic pigments have been utilized in numerous industries including textile, cosmetic, food and pharmaceuticals. However, the drawbacks of these pigments, namely toxicity problems have kindled interest in natural pigments. In view of this, the use of natural pigments such as those from a bacterial origin offers an interesting alternative for industrial application. However, large scale applications of natural pigments are often hindered by the high production cost. This study evaluates the feasibility of using liquid pineapple waste for the production of violacein by a locally isolated Chromobacterium violaceum UTM5 both in a shake flask and a 50 L bioreactor. The use of optimized growth parameters including culture conditions, concentration of liquid pineapple waste and supplementation of L-tryptophan resulted in a violacein yield of 16 256 ± 440 mg L−1. Post treatment of the effluent effectively reduced the COD, turbidity and TSS contents to less than 1 mg L−1, 1.57 ± 0.2 NTU and 2.7 ± 0.6 mg L−1 respectively. The violet pigment exhibited good stability during the entire storage period of 30 days at pH 7, temperature 25–30 °C and under dark conditions. The violet pigment has a good antimicrobial activity against selected microorganisms. Of interest, the pigment was active against Staphylococcus aureus ATCC 29213 and methicillin-resistant Staphylococcus aureus (MRSA) ATCC 43300 with a MIC value of 7.8 and 15.6 μg mL−1, respectively. However, the pigment is toxic to the V79-4 Chinese hamster lung cells with low selectivity index. The purified compounds were determined as violacein and deoxyviolacein respectively using FT-IR, LC-MS and NMR. The results confirmed the feasibility of using liquid pineapple waste as a potential low cost growth medium for the large-scale cultivation of violet pigment using C. violaceum UTM5.
 Please wait while we load your content...
Please wait while we load your content...

