
Effect of azo dye on ammonium oxidation process and ammonia-oxidizing bacteria (AOB) in soil
Abstract
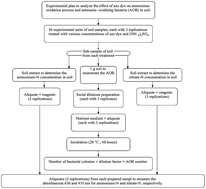
Ammonia-oxidizing bacteria (AOB) play a key role in the production of nitrate-N (NO3−-N) in terrestrial ecosystems. A study was planned with the aim of assessing the effect of azo dyes released by textile and dyestuff industries on the NH4+-N oxidation process in soil. The data was analyzed statistically using a two factorial completely randomized design (CRD). The results of the study demonstrated that higher doses of reactive black 5 (RB5) significantly suppressed the NH4+-N oxidation process throughout incubation. Average percent inhibition rates (%) were in the following order: coarse > fine > medium soil. Overall average percent inhibition rates (%) of nitrification in soils exposed to 30 mg-N kg−1 soil ammonium sulfate [(NH4)2SO4] were 46–53% higher than those from 90 mg-N kg−1 soil. This may be attributed to (NH4)2SO4 that acts as a substrate for the proliferation of AOB. NO3−-N concentration was strongly negatively correlated (r = −0.86) with various amounts of RB5, whereas a strong positive response was observed for the inhibition rate (r = 0.92). A considerable decrease in AOB population (up to 92.58%) was detected for >200 mg kg−1 soil plus N fertilizer, which differed with soil type. This study could be helpful to investigate the effect of contaminants on biochemical processes occurring in soil. Furthermore, the inhibitory effect of azo dye on the NH4+-N oxidation process suggests that critical concentrations of organic dyes may be used as an inhibitor to release NO3−-N in soil at a slow rate in order to further reduce NO3−-N contamination in terrestrial and aquatic ecosystems and to allow less frequent application of ammonium fertilizer in soil as well.
 Please wait while we load your content...
Please wait while we load your content...

