
Manipulating multi-system of NADPH regulation in Escherichia coli for enhanced S-adenosylmethionine production†
Yawei Chenab,
Duanbin Xua,
Lihai Fana,
Xu Zhanga and
Tianwei Tan*a
aNational Energy R&D Center for Biorefinery, College of Life Science and Technology, Beijing University of Chemical Technology, North Third Ring Road 15, Chaoyang District, Beijing, 100029, PR China. E-mail: twtan@mail.buct.edu.cn; Fax: +86 10 64715443; Tel: +86 10 64416691
bCollege of Chemical and Pharmaceutical Engineering, Henan University of Science and Technology, Luoyang 471003, PR China
First published on 29th April 2015
Abstract
S-adenosylmethionine (SAM) is an essential regulatory compound in many biological reactions and an active small molecule of high biomedical value. Cofactor engineering as an advantageous and emerging strategy was introduced to improve the SAM production. In this work, we constructed two kinds of NADPH regenerators of heterologous NADH kinase (Pos5p) and NADH kinase-like enzymes (combination of transhydrogenase PntAB and NAD kinase YfjB) to increase the availability of NADPH in SAM biosynthesis. Modulating of Pos5p resulted in a superior SAM production and led to approximately 5.30 mg L−1 of SAM in Escherichia coli without L-methionine addition. In addition, the synthetic sRNA-based NADPH regulation strategy was also implemented for enhancing the NADPH supply coupled with decreasing the by-products in the SAM synthesis pathway. The SAM titer of the recombinant strains harboring synthetic sRNA could exceed 1 mg L−1, which increased approximately 70% than that of the control. It was also proposed that the production of SAM is not only dependent on the absolute value of NADPH but also dependent on the NADPH/NADP+ ratio. These NADPH regulation strategies are potentially applicable to various processes for enhancing of NADPH-dependent products.
1. Introduction
S-adenosylmethionine (SAM) is an active small molecule of high biomedical value,1 and it is used widely in the treatment of various diseases, such as arthritis,2 depression,3,4 liver disease,1,5 and Alzheimer's disease.6 In recent years, SAM is also added into cosmetics and health care products as an essential ingredient. Owing to the commercial demand of SAM, several attempts have been made to improve SAM production with many kinds of microorganisms.7,8 Conversion of L-methionine, catalyzed by adenosyltransferase is the final step in the SAM synthesis, and the intracellular L-methionine concentration was confirmed to be a limited precursor in the SAM biosynthesis. Optimization of process on adding L-methionine was applied in SAM production.9 As SAM is not the final product in the cell and could be further converted to S-adenosyl-L-homocysteine and other metabolites, it's difficult to accumulate SAM in a high content.10,11 However, fewer reports had focused on the increasing the intracellular concentration of L-methionine using cofactor manipulation. In L-methionine biosynthesis, homoserine dehydrogenase requires NADPH as a cofactor for the reduction of L-aspartate 4-semialdehyde and aspartate semialdehyde dehydrogenase requires NADPH as reducing power for the reduction of L-4-aspartyl phosphate in E. coli (Fig. 1). The availability of NADPH and its redox balance seems to be important in L-methionine production. | ||
| Fig. 1 Simplified SAM synthetic pathway and constructed NADPH regulation strategies in E. coli. NADPH regeneration strategy (1) using heterologous NADH kinase Pos5p as the NADPH regenerator; NADPH regeneration strategy (2) using NADH kinase-like enzymes (combination of transhydrogenase PntAB and NAD kinase YfjB) as the NADPH regenerator; synthetic sRNAs-based NADPH generation strategy: applying sRNA-based strategy to repress the expression of genes involved in the NADPH competitive pathway. | ||
Compared with the direct pathway metabolic engineering, cofactor engineering developed recently could be a promising approach for further enhancing SAM production. Cofactors of NADPH/NADP+, ATP/ADP and NADH/NAD+ are the most highly connected metabolites in metabolic networks,12 and they provide redox carriers for biosynthetic reactions, catabolic reactions and act as principal agents in transfer of energy to the cell. It has been found that cofactors could regulate the expression level of many genes and redirect the flux of central carbon metabolism.13,14 Cofactor regulation has been applied successfully for enhancing the production of the target in many cases.15–18 However, it was not even considered in the production of SAM.
A synthetic small regulatory RNA (sRNA) based strategy is a highly efficient gene knock-down approach that can precisely control the expression of target genes in prokaryotes at the post-transcriptional level. Synthetic sRNA has gained great attraction because of their relatively short length (∼100 bp) that facilitates the implementation. Owing to their advantages, such as screening strain with highest metabolic capacity, identifying best repression target in chromosomal genes, and fine-tuning the expression level of repression target, there have been studies on the metabolic engineering using the synthetic sRNA-based strategy. Na et al. isolated an E. coli capable of producing 2 g L−1 tyrosine using synthetic sRNAs.19 Kim et al. developed an E. coli strain for phenol production by simultaneous metabolic engineering of 18 different E. coli strains using sRNA technology.20 To our knowledge, the synthetic sRNA-based strategy has not been used in NADPH regulating for target production.
In this work, strategies of NADPH regeneration and synthetic sRNA-based NADPH regulation were applied in SAM production respectively (Fig. 1). Two kinds of NADPH regenerators of heterologous NADH kinase (Pos5p) and co-overexpression of the transhydrogenase (PntAB) with NAD kinase (YfjB) were employed to verify the effect of NADPH regeneration on SAM production. Five genes involved in both NADPH consumption and by-products formation in SAM biosynthesis process were selected as candidates for applying the sRNA-based strategy to improve SAM production. The results indicated that increasing of NADPH level and NADPH/NADP+ ratio led to the improvement of SAM production. The synthetic sRNA-based cofactor regulation strategy was also a promising approach to the target production.
2. Materials and methods
2.1 Strains, plasmids and growth conditions
All strains and plasmids are summarized in Table 1. E. coli seed cultures were grown in Luria-Bertani (LB) medium. The recombinant strains for producing SAM were cultivated at 37 °C with 200 rpm orbital shaking in M9 mineral medium. A defined M9 medium (per liter) contained glucose 20 g, Na2HPO4·2H2O 7.52 g, KH2PO4 3 g, NaCl 0.5 g, NH4Cl 0.5 g, MgSO4·2H2O 0.25 g, CaCl2·2H2O 0.15 g, biotin 1 g, thiamine-HCl 1 g and 10 mL 100× trace elements solution. The 100× trace elements solution (per liter) contained EDTA 5 g, FeCl3·6H2O 0.83 g, ZnCl2 84 mg, CuCl2·2H2O 13 mg, CoCl2·2H2O 10 mg, H3BO3 10 mg, MnCl2·4H2O 1.6 mg. Appropriate antibiotics were added at the following concentrations: 100 μg mL−1 of ampicillin, 50 μg mL−1 of kanamycin. For the induction of the protein expression, the appropriate concentration of IPTG was added after 3 h cultivation and the strains were then grown at 30 °C.| Description | Source | |
|---|---|---|
| Strains | ||
| E. coli TOP10 | Cloning host | Biomed |
| E. coli BL21 (DE3) | Host of plasmids | Biomed |
| Control-N | E. coli BL21 harboring pRSFDuet-1 | This study |
| SSP-1 | E. coli BL21 harboring pRSFD-POS5Δ17 | This study |
| SSP-2 | E. coli BL21 harboring pRSFD-yfjB-pntAB | This study |
| Control-S | E. coli BL21 harboring pSA | This study |
| PRSCA-RFP | E. coli BL21 harboring pSA-rfp | This study |
| AT-RFP | E. coli BL21 harboring pSA-anti-rfp | This study |
| AT-AP1 | E. coli BL21 harboring pSA-anti-aroE | This study |
| AT-AP2 | E. coli BL21 harboring pSA-anti-argC | This study |
| AT-AP3 | E. coli BL21 harboring pSA-anti-proA | This study |
| AT-AP4 | E. coli BL21 harboring pSA-anti-ilvC | This study |
| AT-AP5 | E. coli BL21 harboring pSA-anti-proC | This study |
![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) |
||
| Plasmids | ||
| PRSFuet-1 | KmR, T7 promoter | Novagen |
| PpRFP | Harboring rfp | Lab collection |
| PE76 | yfjB gene in pMD19 T | [21] |
| pRSFD-POS5Δ17 | KmR, T7 promoter, POS5Δ17 cloned into pRSFuet-1 | This study |
| pRSFD-yfjB-pntAB | KmR, T7 promoter, yfjB and pntAB cloned into pRSFuet-1 | This study |
| pSA | KmR, Pr promoter, T7 terminator, encoding MicC | This study |
| pSA-rfp | rfp cloned into pSA | This study |
| pSA-anti-rfp | pSA-rfp harboring anti-rfp synthetic sRNA | This study |
| pSA-anti-aroE | pSA harboring anti-aroE synthetic sRNA | This study |
| pSA-anti-argC | pSA harboring anti-argC synthetic sRNA | This study |
| pSA-anti-proA | pSA harboring anti-proA synthetic sRNA | This study |
| pSA-anti-ilvC | pSA harboring anti-ilvC synthetic sRNA | This study |
| pSA-anti proC | pSA harboring anti-proC synthetic sRNA | This study |
2.2 Plasmids construction
| Constructs | Template | Oligo name (and restriction enzyme) | Oligonucleotides (5′–3′) |
|---|---|---|---|
| POS5Δ17 | S. cerevisiae | POS5Δ17 F (Bgl II) | GAAGATCTCAGTACGTTGGATTCACATTCCCT |
| POS5Δ17 R (Xho I) | CCGCTCGAGTTAATCATTATCAGTCTGTCTCTTGGTC | ||
| yfjB | pE76 | yfjB-F (EcoR V) | TGCAGGATATCGATGAATAATCATTTCAAGTGTATTGGCATTGTG |
| yfjB-R (Kpn I) | CGGGGTACCTTAGAATAATTTTTTTGACCAGCCG | ||
| pntAB | E.coli BL21 | pntAB-F (Sac I) | CGAGCTCGATGCGAATTGGCATACCAAG |
| anti-aroE | pRA | anti-aroE-F | ATAGGTTTCCATTTTCTGTTGGGCCATTGCATTG |
| anti-aroE-R | ATAGGTTTCCATGCAACCATTATCACCGCCAGAG | ||
| anti-argC | pRA | anti-argC-F | CGTATTCAACATTTTCTGTTGGGCCATTGCATTG |
| anti-argC-R | CTGATTGTGGGTGCAACCATTATCACCGCCAGAG | ||
| anti-proA | pRA | anti-proA-F | TTGTTCCAGCATTTTCTGTTGGGCCATTGCATTG |
| anti-proA-R | ATGGGCATTGCCGCAACCATTATCACCGCCAGAG | ||
| anti-ilvC | pRA | anti-ilvC-F | GTAGTTAGCCATTTTCTGTTGGGCCATTGCATTG |
| anti-ilvC-R | TTCAATACACTGGCAACCATTATCACCGCCAGAG | ||
| anti-proC | pRA | anti-proC-F | TTTCTTTTCCATTTTCTGTTGGGCCATTGCATTG |
| anti-proC-R | ATCGGTTTTATTGCAACCATTATCACCGCCAGAG | ||
| pSA-rfp | ppRFP | rfp-F(Bgl II) | GAAGATCTAACAAGAATTGGGACAACTCC |
| rfp-R (Xho I) | CCGCTCGAGAATTGTATAAACGCAGAAAGG | ||
| pSA-anti-rfp | pSA-rfp | anti-rfp-F | GGAGGAAGCCATTTTCTGTTGGGCCATTGCATTG |
| anti-rfp-R | GAAGACGTTATCGCAACCATTATCACCGCCAGAG |
| qPCR primers | Oligo name | Oligonucleotides (5′–3′) |
|---|---|---|
| qPCR-gapdh-F | CTCCGCTGGCTAAAGT | |
| qPCR-gapdh-R | GGACGGGATGATGTTCT | |
| qPCR-aroE-F | GTGGTAAAGGTGCGAATG | |
| qPCR-aroE-R | AGGGCGGATAAAAGACAG | |
| qPCR-argC-F | GCCTGAAATCGGGTGT | |
| qPCR-argC-R | GTTGTCTTCGGTCGCC | |
| qPCR-proA-F | AGCGAAATCATCCTCAAC | |
| qPCR-proA-R | AATCACGCCAATAACCCC | |
| qPCR-ilvC-F | GCGTGATTCTGGTCT | |
| qPCR-ilvC-R | GAGTGCTGCTTGTCC | |
| qPCR-proC-F | CGAACACTCCCGCAC | |
| qPCR-proC-R | ACCGCTCACACCGAC |
2.3 Analytical procedures
Cell growth was detected by measuring the optical density at 600 nm. The fluorescence intensity was measured by flow cytometer (BD, USA). SAM was extracted from the fermentation broth with 10% (w/v) perchloric acid for 1 h at 30 °C. The mixture was centrifuged at 13000 rpm for 10 min and filtered through a 0.22 μm membrane. Subsequent analysis of SAM was carried out by HPLC (Shimadzu, Japan) equipped with a C18 column (Agela Technologies, China) and a UV detector (260 nm). The mobile phase consisted of 0.01 mol L−1 ammonium formate at pH 3.5 adjusted with acetate acid at a flow rate of 1 mL min−1. Acetate and lactate were determined by HPLC (Thermo Scientific, USA) using the following apparatus and operating conditions: HPX-87H column (Bio-Rad, USA) with RID and UV detectors (254 nm), 50 °C oven temperature, and 5 mM H2SO4 as mobile phase, 0.6 mL min−1 flow rate. The intracellular NADPH, NADP+, NADH and NAD+ levels were determined using a spectrophotometric cycling assay according to previous procedures.23
The mRNA levels of the sRNA-based strains were determined by real-time quantitative PCR (qPCR). The qPCR primers were listed in Table 2. The E. coli cells were collected by centrifugation of 2 mL cultures after 6 h cultivation. All the kits involved in the RNA preparation, cDNA synthesis and qPCR were purchased from TIANGEN Biotech (Beijing, China). The qPCRs were performed using the method described by previous study.24
3. Results
3.1 Verification of the viability of sRNA-based repression system and design of the target synthetic sRNAs
As shown in Fig. 1, NADPH is required as reducing power to produce SAM and maintain intracellular redox balance. As sRNA-based repression strategy has many advantages in controlling gene expression at post-transcriptional level, we attempted to use this strategy to increase NADPH level coupled with by-products decreased for enhancing SAM production. At first, rfp was used as a reporter gene to insert into the plasmid pSA and constructed the control strain PRSCA-RFP. Strain AT-RFP which contains the anti-rfp synthetic sRNA with the complementary sequence that binded partially to the TIR of rfp was used to testify the effect of sRNA-based down-regulation. The FACS histogram (Fig. 2a) showed that the strain AT-RFP gave significantly lower fluorescence signals than strain PRSCA-RFP, which means synthetic sRNA-based down-regulation system was an efficient known-down strategy in E. coli BL21 (DE3). In Fig. 2b, the relative fluorescence data showed that the strain AT-RFP displayed a distinct repression effect and enabled up to 85% repression efficiency comparing with strain PRSCA-RFP. Then this sRNA-mediated repression system was applied to knock-down genes involved in the NADPH consumption of by-products in the SAM synthesis process.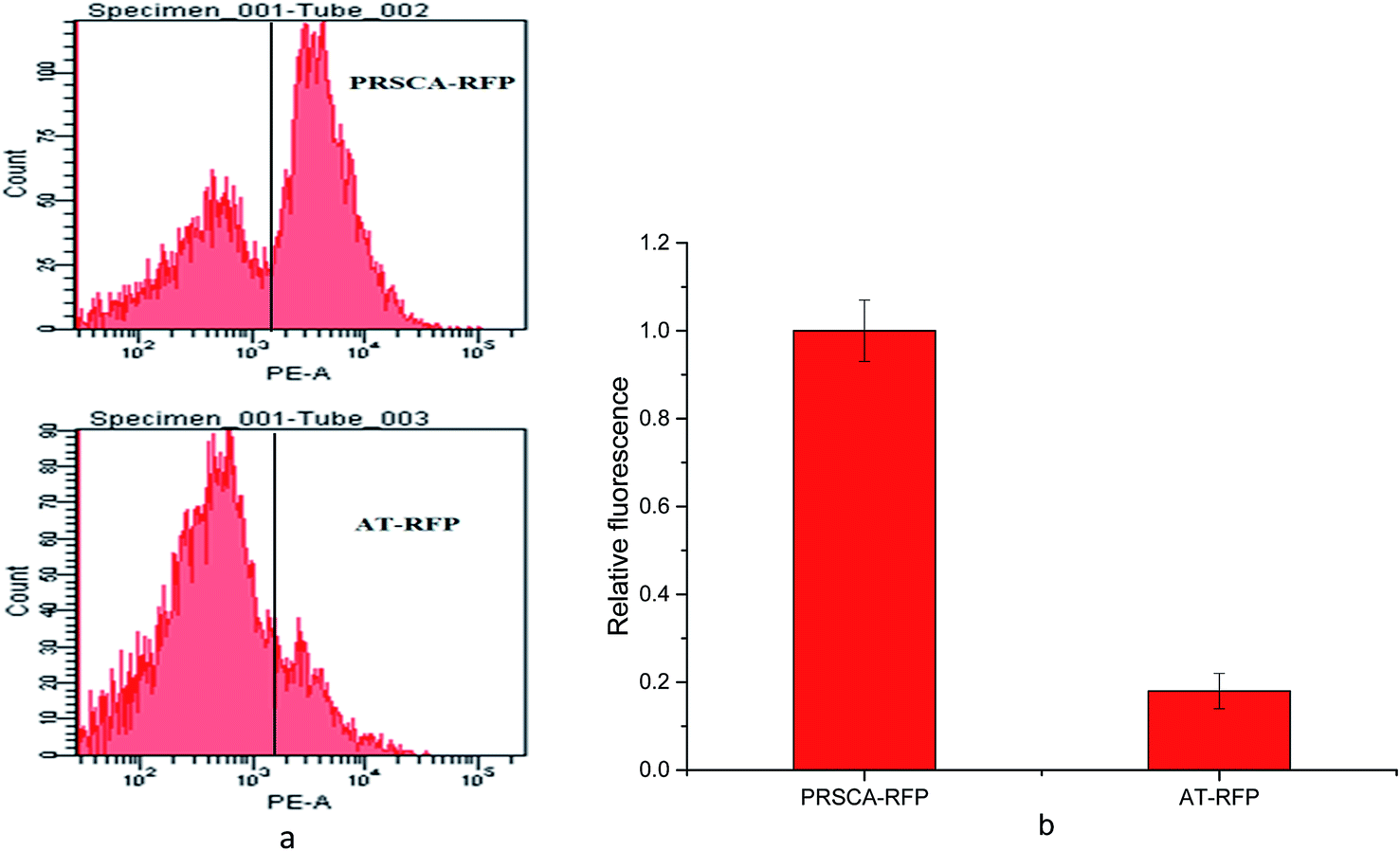 | ||
| Fig. 2 FACS analysis of synthetic sRNAs on repression efficiency. (a) FACS histogram of the strain PRSCA-RFP and AT-RFP; (b) effect of the synthetic sRNA-based strategy on repression efficiency of red fluorescent protein. PRSCA-RFP (strain containing red fluorescent protein and the synthetic sRNA cassette without the target binding sequence); AT-RFP (strain containing red fluorescent protein and the synthetic sRNA cassette with the target binding sequence). | ||
On the basis of the results above, five genes aroE (encoding dehydroshikimate reductase), argC (encoding N-acetyl-gamma-glutamyl phosphate reductase), proA (encoding gamma-glutamyl phosphate reductase), ilvC (encoding ketol-acid reductoisomerase), proC (encoding pyrroline-5-carboxylate reductase) responsible for NADPH consumption were selected as candidate targets for repression to improve SAM production. All five synthetic sRNAs (anti-aroE, anti-argC, anti-proA, anti-ilvC and anti-proC) were designed to have a target-binding sequence of 20–30 nt in length according to the principles of synthetic sRNAs design.22 The binding free energy of the synthetic sRNAs with target-binding sequence and the coding sequence of the target gene was calculated on a web-based service (DINAMelt).25 The calculated binding free energy was of −29 to −45 kcal mol−1, which was exactly in the high repression efficiency range of synthetic sRNAs (Fig. 3).
 | ||
| Fig. 3 The mRNA levels and the calculated binding energy with respect to strains harboring different synthetic sRNAs. aroE: encoding dehydroshikimate reductase; argC encoding N-acetyl-gamma-glutamyl phosphate reductase; proA: encoding gamma-glutamyl phosphate reductase; ilvC: encoding ketol-acid reductoisomerase; proC: encoding pyrroline-5-carboxylate reductase. | ||
In order to further confirm the down-regulation effect of the sRNA-based strategy on the target genes, qPCR was performed to determine the mRNA levels of the target genes. The E.coli glyceraldehyde-3-phosphate dehydrogenase (GAPDH) transcript was chosen to normalize the cDNA samples during expression analysis. The normalized mRNA level in control-S was assumed to be 1.0. As shown in Fig. 3, the mRNA levels of the target genes were affected significantly by the heterogenous sRNA-based system. The strains harboring anti-argC, anti-proA, and anti-proC were observed to reduce relative mRNAs by more than 70%. The smallest efficiency of down-regulation happened in anti-ilvC, and the expression level of the gene ilvC was 81%. Further study should be carried out to explore the effect of sRNA-based NADPH regulation strategy on the SAM production.
3.2 Redistribution of metabolic flux by increasing the NADPH pool
In addition to the sRNA-based strains, there were also NAD (H) kinase-mediated strains to regenerate NADPH. NADH kinase (Pos5p) introduced into plasmid pRSFDuet-1 constructed one kind of NADPH regenerator. Transhydrogenase (PntAB) and NAD kinase (YfjB) were utilized in combination to construct another kind of NADPH regenerator. These NADPH regenerator carrying strains (SSP-1, SSP-2) and sRNA-based strains (AT-AP1, AT-AP2, AT-AP3, AT-AP4, and AT-AP5) were both utilized to increase the intracellular NADPH level for enhancing SAM production. All the recombinant strains showed a higher SAM titer than their control strains. The SAM titer in strains harboring the NADPH regeneration system was much higher than sRNA-based strains, and the strain SSP-1 obtained the highest SAM titer of 3.46 mg L−1 which is approximately 6-fold than that of the Control-N under 50 μg mL−1 of IPTG (Fig. 4). In addition, the recombinant strains showed lower by-products of acetate and lactate than the control strain during flask fermentation. Notably, the sum of acetate and lactate in recombinant strains SSP-1, SPP-2, AT-AP1, AT-AP2, At-AP3, AT-AP4 and AT-AP5 was reduced by 29.94%, 37.20%, 61.80%, 59.14%, 36.92%, 42.91% and 48.50%, respectively (Fig. 4). The reason was mostly contributed to the change of the intracellular redox balance caused by introduced NADPH generation systems. Clearly, disturbance of NADPH pool triggered the redistribution of metabolic flux in E. coli and strengthen biosynthesis of target SAM. | ||
| Fig. 4 Flask fermentation results of by-products concentration and the SAM titer increase in the control and recombinant strains. Control-N: E. coli BL21 harboring pRSFDuet-1; SSP-1: E. coli BL21 harboring pRSFD-POS5Δ17; SSP-2: E. coli BL21 harboring pRSFD-yfjB-pntAB; Control-S: E. coli BL21 harboring pSA; AT-AP1: E. coli BL21 harboring pSA-anti-aroE; AT-AP2: E. coli BL21 harboring pSA-anti-argC; AT-AP3: E. coli BL21 harboring pSA-anti-proA; AT-AP4: E. coli BL21 harboring pSA-anti-ilvC; AT-AP5: E. coli BL21 harboring pSA-anti proC. | ||
3.3 Effect of the NADPH improvement on the cell growth and SAM production
The growth rate of the strains carrying NADPH regeneration system was slower than the strain Control-N. It implied that an excess of NADPH is deleterious to cell, as the high concentration of NADPH hampers its re-oxidation capacity in recombinant strains. However, the cell growth of the strains harboring synthetic sRNAs was almost the same comparing with the strain Control-S. It indicated that synthetic sRNA-based NADPH regulation strategy did not generate extra NADPH but repressed the NADPH consumption of by-products in SAM production. In addition, synthetic sRNAs did not exert burden to cells. The SAM production profiles under 50 μg mL−1 and 100 μg mL−1 of IPTG were measured to explore the impact of different NADPH levels on the SAM biosynthesis. The SAM titer of SSP-1 and SSP-2 strain was much higher than the Control-N strain under different IPTG concentrations according to Fig. 5. As expected, the rate of SAM titer increased under 100 μg mL−1 of IPTG was greater than that of under 50 μg mL−1 of IPTG. Strain SSP-1 obtained the highest SAM titer of 5.30 mg L−1 at 12 h without L-methionine addition, which was more than 13-fold compared with the Control-N strain. All five strains harboring synthetic sRNAs improved the SAM production, and the SAM titer could exceed 1 mg L−1 (Fig. 5f). It was obvious that NADPH regulation strategy was beneficial for SAM production, although the SAM titer was not high enough. | ||
| Fig. 5 Flask fermentation results of the control and the recombinant E. coli. (a and d) NADPH regeneration system on the cell growth and SAM titer under IPTG concentration of 50 μg mL−1; (b and e) NADPH regeneration system on the cell growth and SAM titer under IPTG concentration of 100 μg mL−1; (c and f) the sRNA-based NADPH generation system on the cell growth and SAM production. | ||
3.4 Determination of intracellular pyridine nucleotide pools in SAM production
To further verify the role of NADPH on the production of SAM, the concentrations of intracellular pyridine nucleotides were determined every 6 h. For the recombinant strain SSP-1, the intracellular NADPH and NADP+ levels were increased by 43% and 22% while the NADH and NAD+ levels were decreased by 57% and 16% at 12 h (Fig. 6). In addition, the ratio of NADPH to NADP+ was increased in both of the recombinant strain SSP-1 and SSP-2. It is interesting that the sum of the NADPH, NADP+, NADH and NAD+ level in recombinant strains was almost the same as in the control strain. Comparing with the Control-S, the NADPH/NADP+ ratio of the five sRNA-based NADPH regulation strains was increased (Fig. 7). However, the NADPH/NADP+ ratio of sRNA-based strains was lower than the strains harboring NADPH regenerators, and strain SSP-1 possessed the highest NADPH/NADP+ ratio of 0.70 at 12 h. These results indicated that either NAD (H) kinase-mediated NADPH regeneration strategy or sRNA-based NADPH regulation strategy could change the ratio of NADPH to NADP+ efficiently. It was also proposed that SAM production is not only dependent on the absolute value of NADPH but also on the NADPH/NADP+ ratio. | ||
| Fig. 6 Effects of the NADPH regeneration system on the disturbance of intracellular pyridine nucleotide level in the control and recombinant strains. (a) Intracellular NADPH concentration; (b) intracellular NADP+ concentration; (c) ratio of NADPH to NADP+; (d) intracellular NADH concentration; (e) intracellular NAD+ concentration; (f) ratio of NADH to NAD+. | ||
 | ||
| Fig. 7 Effect of synthetic sRNA-based NADPH regulation systems on the NADPH/NADP+ ratio. | ||
4. Discussion
Microbial fermentation method was expected to be used for production of SAM owing to its lower cost for industrial. As L-methionine is a limited factor during SAM production, most of the researches focused on optimizing fermentation process, manipulating the enzymes that involved in the central pathways, or adding L-methionine directly to the medium to satisfy the SAM synthesis. However, recently developed cofactor engineering has enabled more efficient construction of metabolic pathways for value-added chemicals. It has been proven that the manipulation of cofactors is an effective strategy in various microorganisms.15–18 Nevertheless, there have been fewer reports on SAM production by cofactor engineering at present. According to the biosynthesis pathway of SAM, NADPH is an important cofactor for the biosynthesis of precursor L-methionine which is the limited in SAM production.The oxidative PPP and the TCA cycle were conformed to be the primary sources of NADPH generation in E. coli,26 and the NADPH supplement was still insufficient to NADPH-dependent products. Enhancing the carbon flux through the PPP was the most helpful way of increasing the NADPH concentration for target products.27–30 However, direct enhancement of the PPP, coupled with CO2 formation may result in over-consumption of one carbon and decrease the production yield.
In this study, NADPH regeneration systems and synthetic sRNA-based NADPH regulation systems in the recombinant E. coli strains were firstly developed for the SAM production. NADH kinase (Pos5p) introduced into plasmid pRSFDuet-1 constructed one kind of NADPH regenerator. Transhydrogenase (PntAB) and NAD kinase (YfjB) were utilized in combination to construct another kind of NADPH regenerator. Five genes involved in the NADPH consumption of by-products in SAM synthesis pathway were selected for synthetic sRNA-mediated repression: aroE, argC, proA, ilvC, and proC.
After conducting the cofactor engineering manipulations, we investigated the effect triggered by the disturbance of pyridine nucleotides pool and demonstrated that it has a positive correlation between NADPH availability and SAM production within a certain range. Strategies for NADPH regeneration and sRNA-based NADPH regulation both increased intracellular NADPH concentration and thus improved SAM production. It was also proposed that SAM production is not only dependent on the absolute value of NADPH but also on the NADPH/NADP+ ratio. The significant increase of the NADPH/NADP+ ratio due to the cofactor regulation triggered considerable metabolic flux redistribution (Fig. 4). The decreased concentration of lactate and acetate which contributed to the increased yield of recombinant strains was tightly associated with the perturbation of NADH and NAD+. The excess NADH generated by the glycolysis that would be transferred to the respiratory chain to produce ATP transformed to NADPH catalyzed by the NAD (H) kinase in strains harboring NADPH regeneration system. Generally, the metabolic flux of acetate would be enhanced by this situation as it could alternatively contribute to producing ATP that was essential for cell growth. However, as shown in Fig. 4, the concentration of acetate was reduced in recombinant strains, suggesting that direct phosphorylation and transhydrogenation of NADH might lead to a faster NADPH regeneration and minimized acetate formation that is known to be related to the redox balance of pyridine nucleotides.26 Lactate formation was also inhibited due to the reduced concentration of NADH. Although NADH and NAD pools were not directly disturbed in sRNA-based strains, the concentration of lactate and acetate was also found lower than in the control. It was reasoned that increased NADPH, as highly connected metabolites in the metabolism networks, resulted in the redirection and reduction of the glucose flux.31 The cell growth rate was inhibited in strains with the NADPH regulation system while the growth rate was almost the same in synthetic sRNA-based strains. It implied that the synthetic sRNAs did not exert burden to cells. However, as an excess of NADPH is deleterious to cell, since the high concentration of NADPH hampers its re-oxidation capacity in recombinant strains.32
Pos5p from S. cerevisiae is one of the well-characterized NADH kinases, which presents a more than 50 times higher affinity toward NADH than NAD+.14,33 As expected, strain SSP-1 carrying NADH kinase exhibited the highest NADPH level and NADPH/NADP+ ratio, and it obtained the highest SAM titer and yield. In addition, the decreasing NADH level induced the synthesis of amino acids and also improved the SAM production. Although overexpression of either PntAB or YfjB individually had been commonly used to increase NADPH supply,27,34 these two enzymes had seldom been used in combination. NAD+ produced by PntAB can be phosphorylated to NADP+ by YfjB which is a critical enzyme for controlling the NAD (H) and NADP (H) balance, pushing the reaction direction towards NADPH formation (Fig. 1). Activating transhydrogenase and NAD kinase in combination led to an 88% and 94% increase of SAM titer and yield, respectively, compared to activate transhydrogenase alone. The obtained results suggested that transhydrogenase and NAD kinase had a synergistic effect on increasing the production of SAM. In the NADPH regeneration strains, NADPH was formed through consumption of ATP and NADH. Intracellular concentration ATP was also decreased with the increase of the SAM production (Fig. S1†). In addition, the concentration of methionine and cystathionine which are the intermediates in SAM biosynthesis pathway was also measured to further confirm the effect of the NADPH regeneration strategy. The results showed that production of theses intermediates were both increased comparing with the control strain (Fig. S2†).
The recently developed synthetic sRNA-based gene knock-down strategy was applied for NADPH regulation in E. coli strains for enhanced SAM production. The approach resulted in both enhancing the intracellular NADPH level and reducing the by-products formation. As a result, synthetic sRNA-based strains obtained a higher NADPH/NADP+ ratio and a higher SAM titer, which was increased by approximately 70%, in comparison with the control.
The results showed that these NADPH generation strategies are beneficial to enhance SAM production. Although the SAM titer in the fermentation broth was still low, the relative increase of the SAM titer and yield was significant. Especially, these strategies of NADPH regulation will serve as a platform for additional metabolic engineering to further increase the productivity of NADPH-dependent products.
5. Conclusion
Different NADPH regulation strategies were applied in this work for enhancing production of SAM which is of high biomedical value. Especially, the synthetic sRNA-based NADPH regulation strategy is a potential tool for conveniently balancing the redox status in recombinant strains. We have further demonstrated the relationship of NADPH/NADP+ ratio with SAM production. These strategies of NADPH regulation will serve as a platform for additional metabolic engineering to further increase the productivity of NADPH-dependent products.More research should be carried out to enhance the SAM production through cofactor engineering. Regulation of ATP which is also an essential cofactor in SAM synthesis should be studied. Disturbance of cofactor could affect the whole metabolic network, and the metabolism of amino acid itself is very complex. Therefore, metabolomics and metabolic flux should be analyzed for further improving the SAM production. In addition, the synthetic sRNA-based strategy can be used for screening the effective and non-obvious target genes that could be manipulated for enhanced of the SAM production.
Acknowledgements
The authors gratefully acknowledge the support from the National Basic Research Program of China (973 program) (2013CB733600, 2012CB725200), the National Nature Science Foundation of China (21390202, 21436002) and the National Key Scientific Instruments and Equipment Development Special Fund (2012YQ0401400302).References
- C. S. Lieber, J. Hepatol., 1999, 30, 1155–1159 CrossRef CAS.
- H. J. Hosea Blewett, Crit. Rev. Food Sci. Nutr., 2008, 48, 458–463 CrossRef CAS PubMed.
- D. Mischoulon and M. Fava, Am. J. Clin. Nutr., 2002, 76, 1158S–1161S CAS.
- G. Van der Watt, J. Laugharne and A. Janca, Curr. Opin. Psychiatr., 2008, 21, 37–42 CrossRef PubMed.
- Q. M. Anstee and C. P. Day, J. Hepatol., 2012, 57, 1097–1109 CrossRef CAS PubMed.
- L. D. Morrison, D. D. Smith and S. J. Kish, J. Neurochem., 1996, 67, 1328–1331 CrossRef CAS.
- M. Kanai, M. Masuda, Y. Takaoka, H. Ikeda, K. Masaki, T. Fujii and H. Iefuji, Appl. Microbiol. Biotechnol., 2013, 97, 1183–1190 CrossRef CAS PubMed.
- M. H. Li, X. M. Meng, E. J. Diao, F. L. Du and X. Y. Zhao, J. Chem. Technol. Biotechnol., 2012, 87, 1379–1384 CrossRef CAS PubMed.
- H. Hu, J. C. Qian, J. Chu, Y. H. Wang, Y. P. Zhuang and S. L. Zhang, Appl. Microbiol. Biotechnol., 2009, 83, 1105–1114 CrossRef CAS PubMed.
- T. Bottiglieri, Am. J. Clin. Nutr., 2002, 76, 1151S–1157S CAS.
- N. Martinez-Lopez, M. Varela-Rey, U. Ariz, N. Embade, M. Vazquez-Chantada, D. Fernandez-Ramos, L. Gomez-Santos, S. C. Lu, J. M. Mato and M. L. Martinez-Chantar, Biochem. Soc. Trans., 2008, 36, 848–852 CrossRef CAS PubMed.
- J. Nielsen, J. Bacteriol., 2003, 185, 7031–7035 CrossRef CAS.
- A. K. Holm, L. M. Blank, M. Oldiges, A. Schmid, C. Solem, P. R. Jensen and G. N. Vemuri, J. Biol. Chem., 2010, 23, 17498–17506 CrossRef PubMed.
- J. Hou, N. F. Lages, M. Oldiges and G. N. Vemuri, Metab. Eng., 2009, 11, 253–261 CrossRef CAS PubMed.
- X. J. Ji, Z. F. Xia, N. H. Fu, Z. K. Nie, M. Q. Shen, Q. Q. Tian and H. Huang, Biotechnol. Biofuels, 2013, 6, 7 CrossRef CAS PubMed.
- L. Y. Jiang, Y. Y. Zhang, Z. Li and J. Z. Liu, J. Ind. Microbiol. Biotechnol., 2013, 40, 1143–1151 CrossRef CAS PubMed.
- S. Siedler, S. N. Lindner, S. Bringer, V. F. Wendisch and M. Bott, Appl. Microbiol. Biotechnol., 2013, 97, 143–152 CrossRef CAS PubMed.
- J. A. Sun, L. Y. Zhang, B. Rao, Y. L. Shen and D. Z. Wei, Bioresour. Technol., 2012, 119, 94–98 CrossRef CAS PubMed.
- D. Na, S. M. Yoo, H. Chung, H. Park, J. H. Park and S. Y. Lee, Nat. Biotechnol., 2013, 31, 170–174 CrossRef CAS PubMed.
- B. Kim, H. Park, D. Na and S. Y. Lee, Biotechnol. J., 2014, 9, 621–629 CrossRef CAS PubMed.
- Z. J. Li, L. Cai, Q. Wu and G. Q. Chen, Appl. Microbiol. Biotechnol., 2009, 83, 939–947 CrossRef CAS PubMed.
- S. M. Yoo, D. Na and S. Y. Lee, Nat. Protoc., 2013, 8, 1694–1707 CrossRef CAS PubMed.
- Y. Gibon and F. Larher, Anal. Biochem., 1997, 251, 153–157 CrossRef CAS PubMed.
- M. M. Wang, L. J. Hu, L. H. Fan and T. W. Tan, Energy Fuels, 2015, 29, 1823–1829 CrossRef CAS.
- N. R. Markham and M. Zuker, Nucleic Acids Res., 2005, 33, W577–W581 CrossRef CAS PubMed.
- U. Sauer, F. Canonaco, S. Heri, A. Perrenoud and E. Fischer, J. Biol. Chem., 2004, 279, 6613–6619 CrossRef CAS PubMed.
- W. H. Lee, Y. K. Chin, N. S. Han, M. D. Kim and J. H. Seo, Appl. Microbiol. Biotechnol., 2011, 91, 967–976 CrossRef CAS PubMed.
- W. H. Lee, J. W. Kim, E. H. Park, N. S. Han, M. D. Kim and J. H. Seo, Appl. Microbiol. Biotechnol., 2013, 97, 1561–1569 CrossRef CAS PubMed.
- A. Q. Shi, X. N. Zhu, J. Lu, X. L. Zhang and Y. H. Ma, Metab. Eng., 2013, 16, 1–10 CrossRef CAS PubMed.
- Z. N. Xu, K. J. Jing, Y. Liu and P. L. Cen, J. Ind. Microbiol. Biotechnol., 2007, 34, 83–90 CrossRef CAS PubMed.
- S. Siedler, S. Bringer and M. Bott, Appl. Microbiol. Biotechnol., 2011, 92, 929–937 CrossRef CAS PubMed.
- F. Canonaco, T. A. Hess, S. Heri, T. Wang, T. Szyperski and U. Sauer, FEMS Microbiol. Lett., 2001, 204, 247–252 CrossRef CAS PubMed.
- M. K. Strand, G. R. Stuart, M. J. Longley, M. A. Graziewicz, O. C. Dominick and W. C. Copeland, Eukaryotic Cell, 2003, 2, 809–820 CrossRef CAS.
- H. C. Lee, J. S. Kim, W. Jang and S. Y. Kim, J. Biotechnol., 2010, 149, 24–32 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/c5ra02937f |
| This journal is © The Royal Society of Chemistry 2015 |