
Chronic toxicity of crude ricinine in rats assessed by 1H NMR metabolomics analysis†
Pingping Guo‡
a,
Dandan Wei‡a,
Junsong Wang*b,
Ge Donga,
Qian Zhanga,
Minghua Yanga and
Lingyi Kong*a
aState Key Laboratory of Natural Medicines, Department of Natural Medicinal Chemistry, China Pharmaceutical University, 24 Tong Jia Xiang, Nanjing 210009, PR China. E-mail: cpu_lykong@126.com; Fax: +86-25-8327-1405; Tel: +86-25-8327-1405
bCenter for Molecular Metabolism, Nanjing University of Science & Technology, 200 Xiao Ling Wei Street, Nanjing 210094, PR China. E-mail: wang.junsong@gmail.com; Tel: +86-25-8431-5512
First published on 26th February 2015
Abstract
Ricinine is a toxic alkaloid found in the leaves and seeds of Ricinus communis L. It can cause vomiting and various other toxic reactions, including liver and kidney damage, convulsions, and hypotension, and can even lead to death. In this study, rats were orally administrated with an extract of castor bean shell (crude ricinine) once a day for eight consecutive weeks to study its chronic toxicity. Urine, serum and kidney samples were collected and subjected to 1H NMR metabolomics analysis. This approach, complemented by histopathological inspection and a biochemical assay, demonstrated that crude ricinine produced obvious nephrotoxicity and severe metabolic alterations in rats. These changes were related with oxidative stress, energy metabolism, amino acid metabolism, renal function and the gut bacteria system. This work provides a molecular basis for the chronic toxicity of crude ricinine and shows the power of a 1H NMR-based metabolomic approach to study the toxicity of drugs dynamically and systematically.
Introduction
Ricinus communis L. (Euphorbiaceae) is a perennial shrub or small arbor distributed throughout temperate and tropical regions. It is an important economical plant, as its seeds are the raw material for producing castor oil, which can be used as a food additive, flavoring, lubricant, cosmetic ingredient, and vulneraria.1 The seeds of R. communis (also called the castor bean) contain toxic components, including the alkaloid ricinine2 and the protein lectins ricin. Ricinine (Nr-methyl-3-cyano-4-methoxy-2-pyridone) is a neutral alkaloid found mainly in the leaves and castor bean shell of this plant. As an insecticidal agent,3 ricinine has proven its activity against the Hymenoptera Atta sexdens rubropilosa,4 and the Lepidoptera Spodoptera frugiperda.5 Ricinine can cause vomiting and various other toxic reactions, including liver and kidney damage, convulsions, hypotension, and can even lead to death.6 However, the mechanisms for the toxicity of crude ricinine remain uncertain and have not been fully explored, especially from a holistic perspective.Metabolomics has been widely applied to examine the progression, generation, and recovery from toxic lesions,7–9 and can provide an insight into the integrated function of a complex biosystem at a systemic level.10 As an unbiased, noninvasive, and rapid analysis technique, 1H NMR has been one of the most widely utilized approaches in metabolomics analyses. The 1H NMR spectra of biofluids or tissues are also rich in structural information and could provide a rapid, non-destructive and high-throughput means for metabolomic profiling. Pattern recognition techniques, such as orthogonal signal correction partial least squares discriminate analysis (OSC-PLS-DA) and other statistic analyses could be used to denote and assess the holistic biochemical changes.11,12 Metabolomics is consistent with the holistic approach of traditional Chinese medicine (TCM) and is sensitive to stimuli, and thus is suitable for the mechanistic and systematic study of the toxicities of TCMs,13–15 which to date has been complicated due to the complex components contained in TCMs.
In our previous study, the toxicity of crude protein ricin from castor bean kernels on rats was successfully assessed by an NMR-based metabolomics approach.16 In a continuation of our studies on the toxic components of castor beans, the chronic toxicity of crude alkaloid ricinine was investigated. Rats were orally administrated with crude ricinine from castor bean shell once a day for eight consecutive weeks. Urine, serum, and kidney samples were collected and recorded for their 1H NMR spectra, which were then analyzed by multivariate OSC-PLS-DA and univariate techniques. This NMR-based metabolomics approach, complemented with biochemical and histopathological observations, revealed a series of metabolic pathway perturbations concerning oxidative stress, amino acid metabolism, energy metabolism, renal function, and the gut bacteria system produced in rats after eight weeks treatment with crude ricinine.
Experimental
Chemicals, reagents, and herbal materials
The seeds of R. communis were purchased from Anguo Qirui Chinese Herbal Medicine Company (Hebei, China) and identified by Professor Mian Zhang, Department of Medicinal Plants, China Pharmaceutical University, Nanjing, China. The voucher specimen was deposited in the Department of Natural Medicinal Chemistry, China Pharmaceutical University. The kits of blood urea nitrogen (BUN), urine urea nitrogen (UUN), serum creatinine (SCR), urine creatinine (UCR), urine protein (UP), urine N-acetyl-β-D-glucosaminidase (NAG), and the ELISA Assay kit of rat retinol binding protein (RBP) were bought from Nanjing Jiancheng Bioengineering Institute (Nanjing, China). Deuterium oxide (D2O, 99.9%) was purchased from Sigma Chemical Co. (St. Louis, MO, USA). Distilled water was purified using a Milli-Q system (Millipore, Bedford, MA, USA).Herbal material process
Dried and crushed castor bean shell (3600 g) was extracted with 50% ethanol (1![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :8 w/v) under reflux for 2 h for three times. The filtrates were combined, concentrated under reduced pressure, and lyophilized to give yellow crude extracts (9.5% yield). The dried extracts were suspended in 0.5% (w/v) sodium carboxymethylcellulose (CMC-Na) and the doses were calculated as raw material weights for the animal experiments. Ricinine from the yellow residue was indentified by 1H-NMR and LC-MS.
:8 w/v) under reflux for 2 h for three times. The filtrates were combined, concentrated under reduced pressure, and lyophilized to give yellow crude extracts (9.5% yield). The dried extracts were suspended in 0.5% (w/v) sodium carboxymethylcellulose (CMC-Na) and the doses were calculated as raw material weights for the animal experiments. Ricinine from the yellow residue was indentified by 1H-NMR and LC-MS.
Analysis of ricinine by 1H NMR and LC-MS
The 1H NMR spectrum of ricinine was recorded on a Bruker Avance 500 MHz spectrometer (Bruker Biospin, Germany).The HPLC analyses were performed using an Agilent 1290 HPLC instrument (Agilent, Waldbronn, Germany) equipped with a binary pump, an online egasser, an autosampler, and a thermostatically controlled column compartment. The samples were separated on an Agilent ZorBax Eclipse XDB-C18 column (4.6 × 220 mm, 5 μm, Agilent Corporation, Santa Clara, CA, USA). The mobile phase consisted of methanol (solvent A) and water (solvent B) prepared using a gradient elution according to the following profile: 0.0–10.0 min, 10–25% B; 10.5–16.5 min, 50% B; 17.0–25.0 min, 70–100% B. The flow rate was 1 mL min−1 and the column temperature was set at 30 °C.
Detections were performed using an Agilent 6520 QTOF mass spectrometer (Agilent Corporation), which was connected to the Agilent 1290 UHPLC instrument via an ESI interface. The operating parameters were as follows: drying gas (N2); flow rate, 8.0 L min−1; temperature, 320 °C; nebuliser, 35 psig; capillary, 4000 V; fragmentor, 175 V; skimmer, 65 V; OCT RF V, 750 V. All the operation, acquisition, and analysis of data were performed using Masshunter workstation software Version B.04.00 (Agilent). The sample was analysed in positive ion mode. The [M + H]+ ion of interest in the positive mode was selected as the precursor ion. The collision energy (CE) was adjusted from 30 eV to 55 eV, and the mass range from m/z 100 to 1000.
Animals and treatment
A total of 36 male Sprague-Dawley rats (220 ± 10 g) were purchased from the Experimental Animal Center of Yangzhou University (Yangzhou, China). The rats were housed in a climate-controlled room at a temperature of 25 ± 3 °C and a relative humidity of 50 ± 10%, with a 12 h light/12 h dark cycle. Food and tap water were provided ad libitum. The rats were acclimatized for one week in stainless steel wire-mesh cages before treatment. The study was approved by the Jiangsu Animal Care and Use Committee and followed the national and institutional rules considering animal experiments.The rats were randomly divided into three groups, 12 animals each, as follows: those administered with crude ricinine corresponding to a raw castor bean shell at a dose of 10 g and 4 g/(kg per day) were classed as the high dose group (HD) and the low dose group (LD), respectively, and those treated with the same volume of CMC-Na were the control group (NC). The oral gavage administration was adopted and performed once a day for eight consecutive weeks.
Collection and preparation of samples
Urine samples were collected into vials using metabolic cages overnight (from 8:00 pm to 8:00 am): the rats were deprived of food to avoid solid debris pollution, but were allowed free access to tap water. The collected urine samples were centrifuged at 12000 rpm for 10 min to aid the settling of coarse material and were then kept at −80 °C before use.
Blood samples were taken from ocular veins of the rats after 12 h fasting at weeks 0, 1, 3, 5, and 8 after the treatment. The serum samples were obtained by centrifugation (12000 rpm, 10 min, 4 °C), and stored at −80 °C before analysis.
Histopathology
At the end of the experiment, the rats were fasted overnight and then anaesthetized by an intraperitoneal injection (i.p.) of 3.5% chloralhydrate (350 mg per kg body weight). The kidney and liver were quickly removed, then rinsed with cold PBS, and immersed in 10% neutral-buffered formaldehyde for 24 h, embedded in paraffin, and sliced into 5 μm thicknesses. The sliced sections were stained with hematoxylin and eosin (H&E), and examined by light microscopy (200× and 400×).Biochemistry and kidney index
To assess renal function, the concentrations of BUN and SCR in serum, and UUN, UCR, UP, NAG, and RBP in urine were determined, and the kidney index (kidney weight/body weight) was calculated.Sample preparation for NMR recording
Frozen kidney tissues (500–600 mg) were homogenized in a mixture of volumetric equivalent acetonitrile and water (5 mL g−1 tissue) in an ice/water bath and centrifuged at 12000 rpm for 10 min at 4 °C. The supernatant was collected and concentrated under a stream of nitrogen and lyophilized. Dried kidney extracts were reconstituted in 600 μL D2O (0.2 M Na2HPO4 and 0.2 M NaH2PO4, pH 7.0, containing 0.05% TSP). To 400 μL urine or 300 μL serum samples, 200 or 300 μL D2O were added, respectively, in order to minimize any NMR shift variation due to the pH discrepancy.
The suspension was vortexed, and then centrifuged at 12000 rpm for 10 min to remove any precipitates. Aliquots of the resulting supernatant (450 μL) were pipetted into 5 mm NMR tubes. TSP was used as the chemical shift reference (δH = 0.00), and D2O provided the field frequency lock signal.
1H NMR spectrometry
1H NMR spectra of the urine and kidney samples were acquired at 298 K on a Bruker Avance 500 MHz spectrometer with a Bruker 5 mm probe, using a modified nuclear Overhauser enhancement spectroscopy (NOESY) pulse sequence to suppress the residual water signal. Free induction delays (FIDs) were collected with 1024 transients into 32768 data points using a spectral width of 10000 Hz with a relaxation delay of 2 s, an acquisition time of 4 s, and a mixing time of 100 ms. All the spectra were zero-filled to 64k data points, and a line-broadening of 0.5 Hz was applied.
The 1H NMR spectra of the serum were recorded on a Bruker Avance 500 MHz spectrometer using the Carr–Purcell–Meiboom–Gill (CPMG) pulse sequence to attenuate the NMR signals of any residual proteins, with a water presaturation applied for a recycle delay of 2 s and a mixing time of 100 ms. Typically, 128 free induction decays (FIDs) were collected into 64k points using a spectral width of 10000 Hz, an acquisition time of 1.36 s, and a relaxation delay of 1.5 s. The Fourier transformed NMR spectra were manually phased and automatically baseline corrected.
Data analysis
The 1H NMR spectra were converted to ASCII files using MestReNova (Version 8.0.1, Mestrelab Research SL), and aligned based on a least square minimization with the shift corrected by the TSP signal. The spectral ranges of δ 0.40–4.16 and δ 5.7–8.5 for urine, δ 0.60–4.25 for serum, and δ: 0.60–4.60 and δ 5.10–9.50 for the kidney were binned into integrated segments of equal width of 0.005 ppm using the R software (http://cran.r-project.org/). The region of 4.16–5.7 ppm in the urine spectra and 4.60–5.10 ppm in the kidney spectra were excluded to remove those regions affected by residual water. All the spectra were normalized by probabilistic quotient normalization. The data were centered and Pareto-scaled before the multivariate analysis. A non-supervised principal components analysis (PCA) revealed no obvious clustering of groups (data not shown). A supervised OSC-PLS-DA method was then carried out to remove any systematic variations unrelated to the interested status through an orthogonal filter. Repeated twofold cross-validation (20 times) was applied in the OSC-PLS-DA model; the validity of the models against overfitting was assessed by the parameter R2, and the predictive ability was described by Q2. The classification performance was evaluated by analyzing the receiver operating characteristic (ROC) plots generated using the R-package ROCR (http://rocr.bioinf.mpi-sb.mpg.de). For each classification, the average prediction accuracy given as the arithmetic mean ± SD of the individual results and the area under the ROC curve (AUROC) was given. The integration areas of the detected metabolites with potential differentiating ability were first tested for their normality of distribution. If the distribution followed the normality assumption, a parametric Student's t-test was applied; otherwise, a nonparametric Mann–Whitney test was performed to detect statistically significant metabolites that were increased or decreased between groups over time. Data were expressed as mean ± SD, and P < 0.05 was considered as statistically significant.Results
Identification of ricinine by 1H NMR and LC-MS
Ricinine was indentified by 1H NMR and LC-MS. 1H NMR (500 MHz, DMSO): δ 8.10 (1H, d, J = 7.8 Hz, 6-H), 6.43 (1H, d, J = 7.8 Hz, 5-H), 3.98 (3H, s, OCH3), 3.43 (3H, s, CH3). The results are consistent with those reported in the literature.6 According to the precursor ion [M + H]+ of 165.0659, corresponding to an elemental composition of C8H8N2O2, the signal of response at an acquisition time of 9.818 min was identified as ricinine (Fig. S1†).Histopathology
The livers and kidneys of the rats exposed to crude ricinine were examined for histopathology. The kidney section of the NC rat showed an apparently normal structure in renal glomerulus and tubule (Fig. 1A and D). The kidney of the HD rats showed significant tubular epithelial cell degeneration (edema) and diaphanous tubular cast (Fig. 1C and F); the kidney of the LD rats showed a moderate degeneration (Fig. 1B and E). No significant pathological changes were observed in the liver tissues of the dosed rats.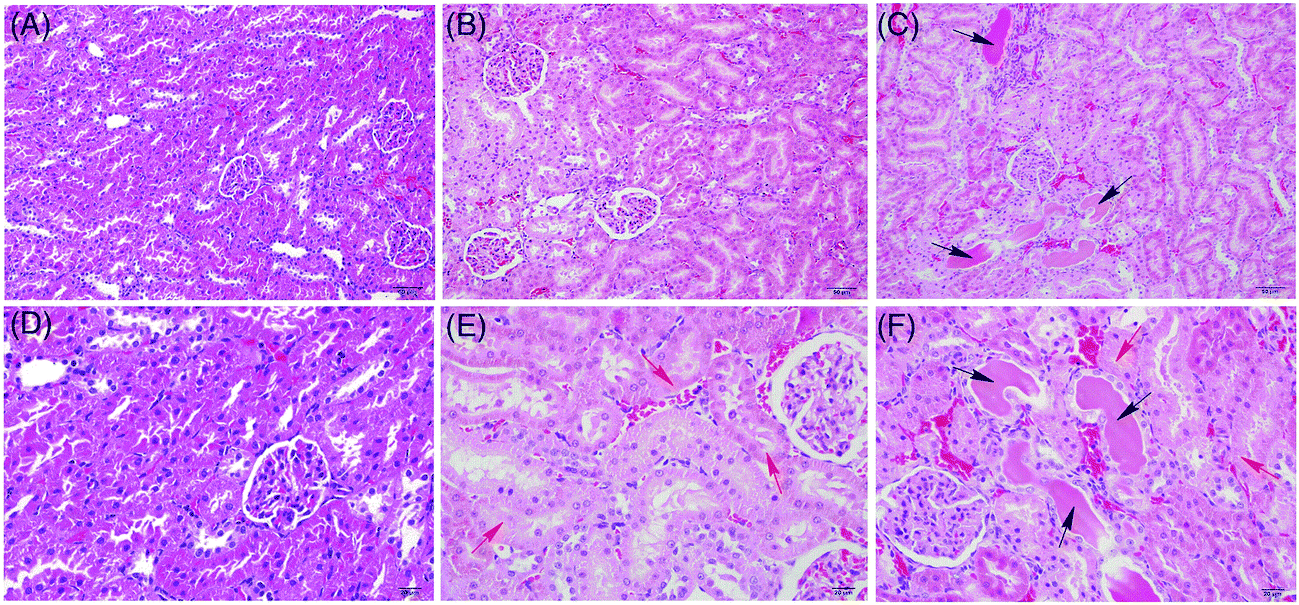 | ||
| Fig. 1 Histopathological photomicrographs of rat kidney (A, B and C ×200; D, E and F ×400) manifested from hematoxylin–eosin (HE) staining. The kidney of the HD rats (C and F) showed severe tubular epithelial cell edema (red arrow) and diaphanous tubular cast (black arrow) as compared with those of the NC (A and D), and the kidney of the LD rats (B and E) showed moderate edema (red arrow). | ||
Biochemistry
The levels of BUN, UUN, SCR, UCR, UP, NAG, and RBP were measured, and the kidney indexes of all the groups of rats were calculated to assess the kidney function (Fig. 2). BUN in the dosed groups did not show any significant difference at week 1, but the difference increased gradually from week 3, reaching a maximum difference at week 8. SCR followed a similar but delayed trend compared with BUN: its level in the dosed groups showed no significant difference from the control group from week 1 to week 3, but then started to increase from week 5, reaching the maximum at week 8. The UUN and UCR of the dosed groups showed some fluctuations throughout the experiments: decreasing markedly at week 1 (early stage) and week 8 (late stage), but staying at nearly the same levels at other time periods. The UP, NAG, and RBP concentrations of the dosed groups showed a significant increase in all time periods, but alleviated from week 3 to week 5. However, the kidney index of the dosed groups only showed a slight decrease, without any significance change. | ||
| Fig. 2 Boxplots for values of BUN (A), UUN (B), SCR (C), UCR (D), UP (E), NAG (F), RBP (G) and the kidney index (H) in the control and dosed groups. At the bottom of each box, the line drawn in the box and at the top of the box represent the 1st, 2nd, and 3rd quartiles, respectively. The whiskers extend to ±1.5 times the interquartile range (from the 1st to 3rd quartile). Outliers are shown as an open circle. All the values are mean ± SD (n = 8). * P < 0.05 and ** P < 0.01 vs. NC. | ||
1H NMR spectra analyses
Typical 1H NMR spectra of urine, serum, and the kidney extract for the HD group and the NC group at week 8 are presented in Fig. 3, with the major metabolites labeled. Aided by the STOCSY technique, their assignments (Table 1) were made by referencing the reported data and searching publicly accessible metabolomic databases, such as HMDB (http://www.hmdb.ca), MMCD (http://mmcd.nmrfam.wisc.edu), and ECMDB (http://www.ecmdb.ca). | ||
| Fig. 3 Typical 500 MHz 1H NMR spectra of urine (A), serum (B), and kidney (C) at week 8 from the HD group (in blue) and NC group (in green). Metabolites in urine: 1, isoleucine/leucine; 2, valine; 3, lactate; 4, alanine; 5, acetate; 6, succinate; 7, 2-oxoglutarate; 8, citrate; 9, dimethylamine; 10, trimethylamine; 11, N,N-dimethylglycine; 12, creatinine; 13, choline; 14, phosphocholine; 15, taurine; 16, trimethylamine-N-oxide; 17, glycine; 18, urea; 19, 4-hydroxyphenylacetate; 20, tyrosine; 21, phenylalanine; 22, hippurate; 23, benzoate; 24, trigonelline; 25, formate. Metabolites in serum: 1, lipoprotein (LDL/VLDL); 2, leucine/isoleucine; 3, valine; 4, β-hydroxybutyrate; 5, lactate; 6, alanine; 7, arginine; 8, N-acetyl-glycoproteins; 9, O-acetyl-glycoproteins; 10, glutamate; 11, glutamine; 12, acetoacetate; 13, pyruvate; 14, citrate; 15, creatinine; 16, taurine; 17, trimethylamine-N-oxide; 18, glucose. Metabolites in kidney: 1, leucine/isoleucine; 2, valine; 3, β-hydroxybutyrate; 4, lactate; 5, alanine; 6, arginine; 7, acetate; 8, glutamate; 9, acetoacetate; 10, creatinine; 11, choline; 12, phosphocholine; 13, taurine; 14, trimethylamine-N-oxide; 15, scyllo-inositol; 16, myo-inositol; 17, adenosine; 18, uridine; 19, tyrosine; 20, phenylalanine; 21, xanthine; 22, hypoxanthine; 23, nicotinamide; 24, nicotinamide mononucleotide. | ||
| a Multiplicity: s, singlet; d, doublet; t, triplet; q, quartet; m, multiplet; br, broad singlet.b The superscript “1” and “2” mean fold change and P value, respectively (* P < 0.05, ** P < 0.01 and *** P < 0.001); the subscript “H” and “L” mean NC vs. HD and NC vs. LD, respectively.c Color coded according to log2(fold change) using the color bar labeled at the right-hand side. |
|---|
 |
OSC-PLS-DA score trajectory plot of all the groups at all time periods
In order to dynamically explore the chronic effect of crude ricinine on the metabolic pattern of the dosed rats, an OSC-PLS-DA model was constructed to analyze all the urine (R2 = 0.80, Q2 = 0.56, P < 0.005) and serum (R2 = 0.51, Q2 = 0.29, P < 0.005) data acquired from the control and treatment groups at weeks 1, 3, 5, and 8. The trajectory plot (Fig. 4) exhibited a good separation between the HD and NC group, with the LD group in between, showing an apparent dose-dependent toxic effect of crude ricinine. A radical alteration in the metabolomic profiles of the dosed groups happened at week 1, reflecting a prompt response of the body to the dosing. The metabolomic changes in the dosed rats attenuated from week 1 to week 3, and then this trend was terminated from week 3 onwards, decreasing to the minimum at week 5, but finally increasing to another maximum at week 8. The serum data showed a similar pattern, but with a delayed alleviation from week 5. This fluctuation of metabolic pattern change showed a complex response of the organism to counteract the toxicity of the dosing. | ||
| Fig. 4 Score trajectory plot of urine (A) and serum (B) from OSC-PLS-DA analysis of the HD, LD, and NC groups (1, 3, 5, 8 mean the time periods in weeks). | ||
1H NMR metabolomics profiles of the HD and NC groups at week 8
Considering the dose-dependent effects of crude ricinine, the NMR data from the HD and NC rats at week 8 were further analyzed by OSC-PLS-DA to denote the chronic toxic effects of crude ricinine on the rats in the long run. 2CV was used to validate the statistical significance of each model in order to avoid overfits. High values of R2 and Q2 of OSC-PLS-DA of urine, serum, and the kidney data (Fig. 5A, C and E) indicated satisfactory models with reliable predictive ability and minimal classification error. The value of AUROC of urine, serum, and the kidney data were 0.886, 0.999, and 0.994, respectively (Fig. 5B, D and F), showing the satisfactory classifier performance of the OSC-PLS-DA model. The score plots of PC1 versus PC2 (Fig. 6A, C and E), where each point represented one sample, revealed a clear separation of the HD group from the NC group along PC1. In order to identify the spectral bins that were responsible for the inter-class differences, the loadings of the S-plot for the first component were generated (Fig. 6B, D and F). The S-plot is a scatter plot that visualizes both the covariance (X axis) and correlation (Y axis) structure of the loading profiles, and thus would be helpful for filtering interesting metabolites in the projection, and for lowering the risk of false positives in the metabolite selection. The significant metabolites increased in the HD group were in the higher-right quadrant and the decreased ones were in the lower-left quadrant. A corresponding loadings plot color-coded with the correlation coefficients of the metabolites visualized the variables responsible for the separation between the HD and NC groups. The weight of a variable in the discrimination was given by the square of its correlation coefficient (r2), which was color coded from zero in blue to high values in red. The S-plot and color coded loadings plot revealed a large number of metabolites contributing to the clustering of the groups. Compared with the NC group, the following findings were observed in the HD group: elevated levels of lactate, alanine, acetate, phenylalanine, TMA (trimethylamine), 4-HPLA (4-hydroxyphenylacetate), formate in urine; elevated levels of leucine/isoleucine, valine, 3-HB (β-hydroxybutyrate), N-acetyl-glycoproteins, O-acetyl-glycoproteins, acetoacetate, pyruvate, glutamine, glutamate, citrate, creatinine in serum; elevated levels of leucine/isoleucine, valine, lactate, arginine, creatinine, tyrosine, phenylalanine, xanthine in the kidney; reduced levels of tyrosine, hippurate, succinate, 2-OG (2-oxoglutarate), citrate, taurine, TMAO (trimethylamine-N-oxide), TRG (trigonelline), urea, and creatinine in urine; reduced levels of LDL/VLDL, lactate, alanine, arginine, taurine, TMAO, glucose in serum; and reduced levels of choline, myo-inositol, adenosine, uridine, hypoxanthine in kidney. These important differential metabolites were selected based on the loadings plot and S plot of OSC-PLS-DA and were further tested for their between-group difference and found to be mostly significant, as visualized in the fold change plot (Fig. S2†).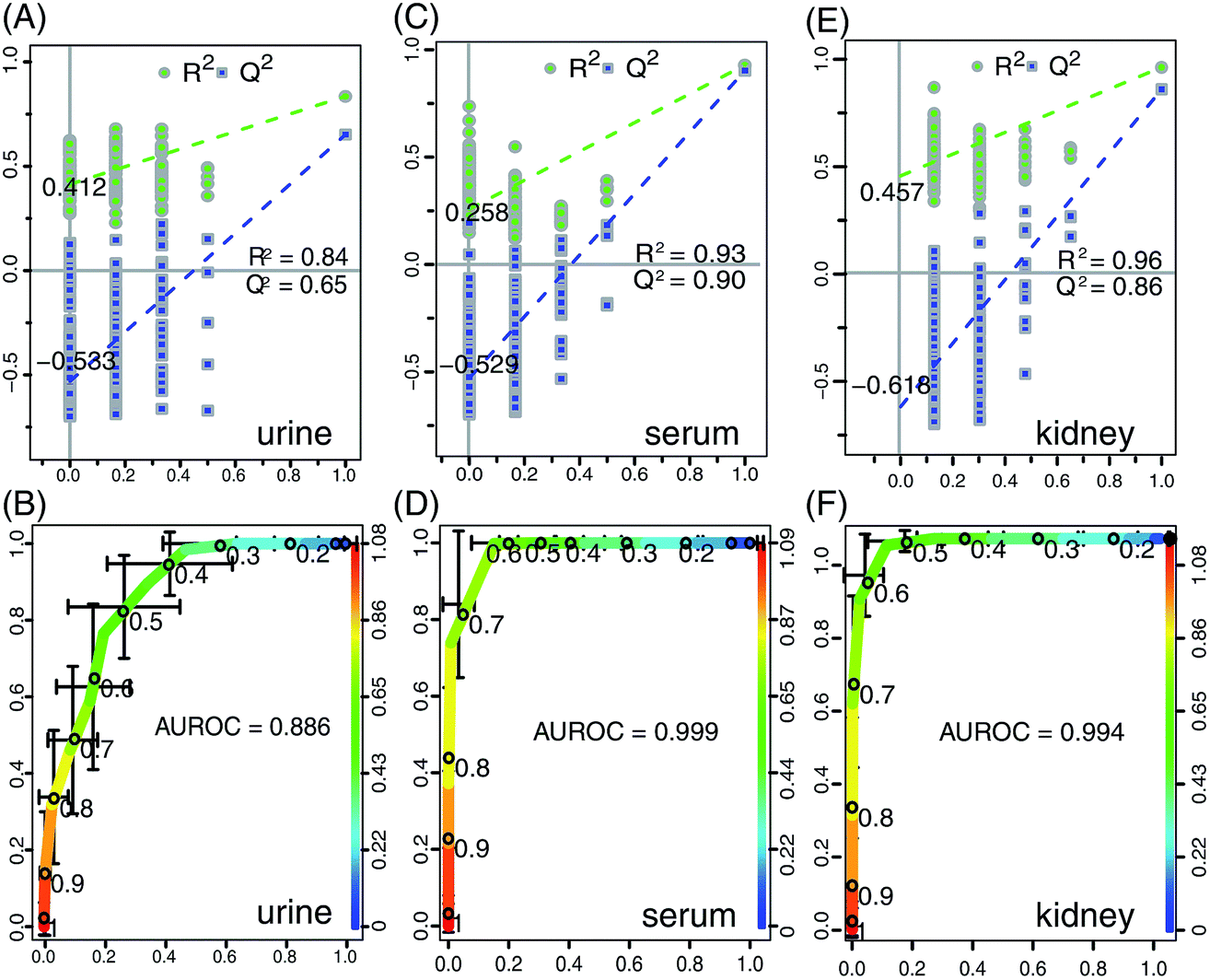 | ||
| Fig. 5 OSC-PLS-DA scatter plot from urine (A), serum (C), and the kidney (E) of the statistical validations obtained by 200 times permutation tests, with R2 and Q2 values in the vertical axis, the correlation coefficients (between the permuted and true class) in the horizontal axis, and OLS line representing the regression of R2 and Q2 on the correlation coefficients. Below are plots of the receiver operating characteristic (ROC) curves of the classifier performance of the OSC-PLS-DA model on the 1H NMR data of urine (B), serum (D) and kidney (F), with the area under the receiver operating characteristic curves (AUROC) labeled. The X-axis denotes the false positive rate, and the Y-axis, the true positive rate. After repeated twofold cross-validation 20 times, the AUROC was calculated. | ||
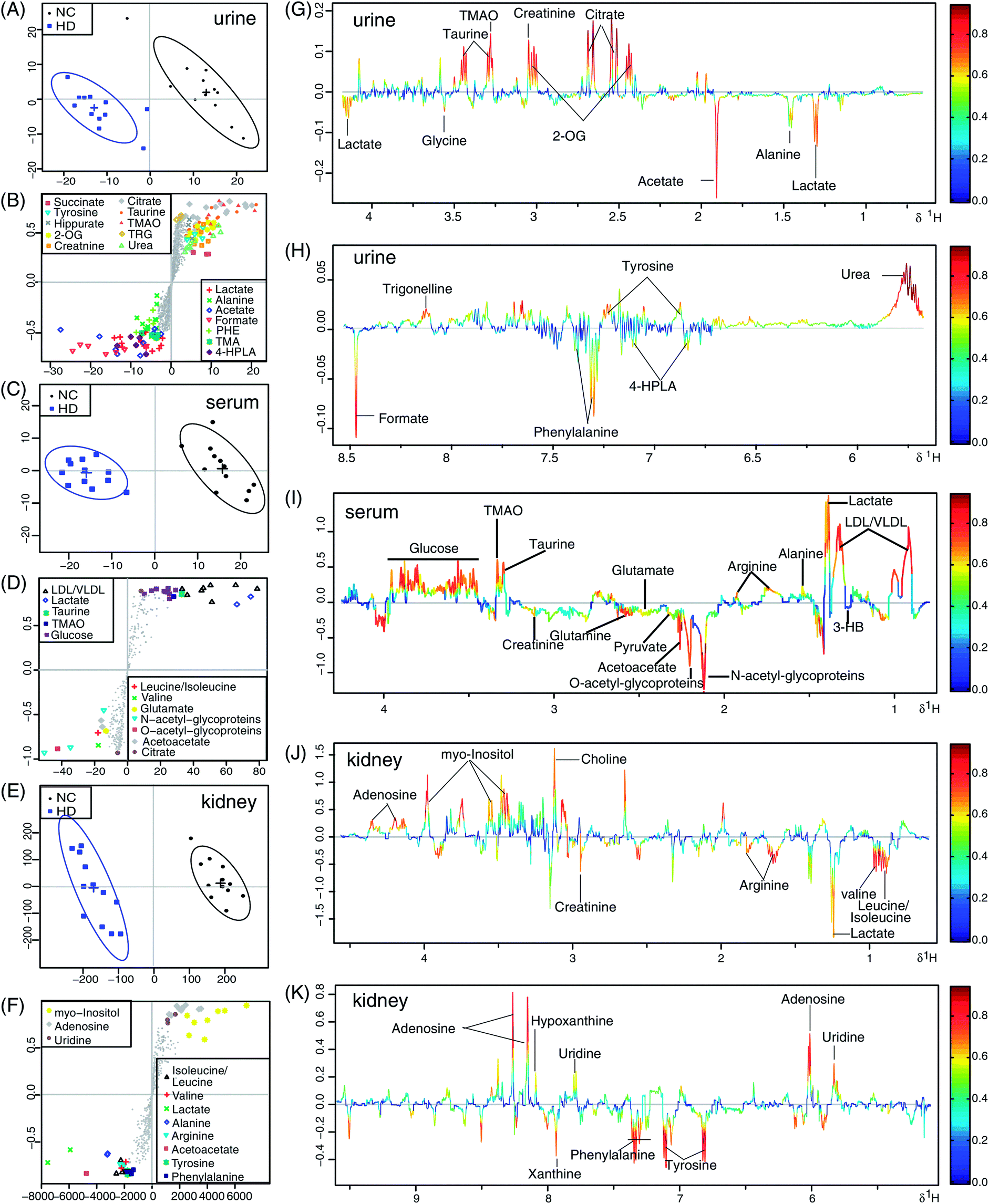 | ||
| Fig. 6 Scores plot (A, C and E), S-plot (B, D and F) and loadings plot with the metabolites labeled (G–K) corresponding to the OSC-PLS-DA analysis of urine (A, B, G and H), serum (C, D and I), and the kidney (E, F, J and K) from the HD and NC groups (n = 12 for each group) at week 8: the loadings plot is color-coded with the correlation coefficients of variables in the OSC-PLS-DA model, with blue the least important metabolic changes and red the most important. Positive peaks indicate a relatively decreased metabolite level in the dosed groups, whereas negative peaks indicate an increased metabolite level in the HD group. | ||
Metabolite pathway analysis
The potential biomarkers selected based on the OSC-PLS-DA loadings plot, S-plot and fold change plot were subjected to a pathway analysis using MetPA (http://www.metaboanalyst.ca) to identify the biologically meaningful metabolic patterns and the most relevant pathways. A hypergeometric test using over-representation analysis and pathway topology analysis (Tables S1–S3†) indicated that valine, leucine and isoleucine biosynthesis, phenylalanine, tyrosine and tryptophan biosynthesis, phenylalanine metabolism, the synthesis and degradation of ketone bodies and the TCA cycle were disturbed in the HD group rats (Fig. S3†).Discussion
The chronic toxicity of crude ricinine in the rats was firstly investigated by histopathological inspection and biochemical evaluation. The kidneys of the dosed rats showed marked tubular epithelial cell edema and diaphanous tubular cast. The urinary excretions of UP, NAG, and RBP were markers of tubular damage17 and alternative indicators of nephropathy,18 and the evaluation of BUN and SCR concentrations were indicators of renal dysfunction.19 Urinary RBP could be used for the early detection of renal tubular dysfunction, sensitively reflecting the damage extent of the renal proximal tubule; while, urinary NAG is a sensitive indicator of renal impairment, particularly on renal tubular ischemic and necrosis. The significantly increased urinary UP, NAG, and RBP after dosing indicated renal damage induced by ricinine. BUN and SCR in the dosed groups were significantly increased at week 8, while UUN and UCR in the dosed groups decreased remarkably at week 1 and 8, suggesting an obvious chronic dysfunction of the kidney.To investigate the variations of endogenous metabolites in rats administered with crude ricinine, a 1H NMR-based metabolomics approach on urine, serum, and the kidney samples was adopted to explore potential biomarkers and the affected metabolic pathways for the first time. OSC-PLS-DA analyses of the urine and serum NMR data of the three groups at all time periods were performed. The metabolic status of the rats was greatly changed by crude ricinine in a dose-dependent manner, peaking at the early (week 1) and late stages (week 8) of the experiments. The NMR data of urine, serum, and the kidney from the HD and NC groups at week 8 were then further analyzed, and revealed a series of metabolic pathway perturbations, including oxidative stress, energy metabolism perturbation, amino acid metabolism disorder, renal damage, and gut bacteria disruption.
Oxidative stress, a serious imbalance between the generation of reactive oxygen species (ROS) and antioxidant defenses, has been demonstrated to be a major mechanism involved in shock, inflammation, and ischemia/reperfusion injury,20 and in the toxicities of some toxins.21–25 Compared with NC rats, decreased levels of serum glutamate and glutamine were observed in the HD rats. As precursors of the major natural antioxidant glutathione (GSH), which combats oxidative injury, the increase of glutamate and glutamine might be a consequence of an inhibited GSH synthesis. The depletion of GSH leads to increased levels of ROS, causing damage to cellular lipids, proteins, or DNA, producing a dysfunction in the body.26 ROS led to the oxidation of membrane lipid, disrupting both the construction and function of membranes, eventually resulting in the rupture of cell and organelles.27 The elevated levels of amino acids (leucine, isoleucine, and valine) in serum and the kidney suggested protein degradation by ROS. Choline and myo-inositol are precursors of all membrane phospholipases;28 their decrease in the kidneys of intoxicated rats therefore suggested an accelerated utilization of them for the construction of damaged membranes, representing a self-repair mechanism. Acetyl-glycoproteins (both N-and O-acetyl glycoproteins) are acute phase proteins, acting as inflammatory mediators and could be a response to tissue damage,29 and thus, the increased concentrations of serum N-acetyl-glycoproteins and O-acetyl-glycoproteins are likely to reflect an inflammatory response.30 Taurine is one of the most abundant free amino acids present in mammalian tissues. It is reported that taurine treatment could decrease oxidative stress and hepatic prooxidant status; also, taurine supplementation may cause an enhancement in GSH levels by directing cysteine into the GSH synthesis pathway.31 Trigonelline (TRG) is an alkaloid containing a pyridine ring, showing activities such as anticancer,32 and improving cognitive function33 and auditory neuropathy.34 It has been reported that the feeding of TRG may suppress oxidative stress by inhibiting the formation of tumor necrosis factor alpha (TNF-α) and the end-products of advanced glycation, which are known to accelerate the production of ROS via NADPH oxidase; could slow fat accumulation, resulting in the suppression of ROS formation; and may downregulate the gene expressions involved with NADPH oxidase and electron transport chain, indicating that TRG may suppress the formation of ROS.35 The decreased levels of taurine in urine, serum, and the kidney and TRG in urine might be a consequence of their overconsumption to counteract ROS generated during intoxication. The decrease in renal uridine and adenosine in the rats treated with ricinine could be ascribed to the promoted purine and pyrimidine catabolism in the kidney by ROS, similar to the CCl4-induced elevations of purine catabolic product (uric acid) in rat serum,36,37 and pyrimidine catabolic product (β-alanine) in rat urine.38
Pyruvate is an important intermediate product of glycolysis, the first step in glucose metabolism where pyruvate is generated by the decomposition of glucose, generating a small amount of ATP. Pyruvate can be used to produce acetyl-CoA by pyruvate dehydrogenase complex. Acetyl-CoA can enter into the TCA cycle, and play a key role in glucose aerobic oxidation and energy production. As important intermediates of the TCA cycle, the decreased levels of 2-oxoglutarate (2-OG), succinate, and citrate might suggest an inhibition of the TCA cycle,39 the most efficient energy supply pattern. To replenish the insufficient energy supply, other means come to the rescue, such as fatty acid β-oxidation. Ketone bodies, comprising acetoacetate, acetone, and β-hydroxybutyrate (3-HB), are well-known metabolites of fatty acids in liver mitochondria. A decreased level of serum lipids (LDL/VLDL) and an increased level of serum ketone bodies indicated an enhanced lipid oxidation. The increased pyruvate and decreased glucose levels in the serum might indicate an enhanced glycolysis to produce energy.
Phenylalanine (PHE) is an essential amino acid and the precursor of tyrosine. Tyrosine is referred to as a semi-essential or conditionally indispensable amino acid because it can only be synthesized by the hydroxylation of PHE catalyzed by phenylalanine hydroxylase (PAH). The increase of PHE and decrease of tyrosine suggested an inhibition of PAH, which was also observed in a previous study on chronic kidney failure.40 4-Hydroxyphenylacetate (4-HPLA) is a tyrosine metabolite and can be converted into tyrosine. The increased level of 4-HPLA in the HD rats might be a result of tyrosine synthesis inhibition, which also suggests nephrotoxicity produced by crude ricinine, since tyrosine has been reported to be reduced substantially in chronic renal impairment.40,41
Alterations in urinary levels of the metabolites have at least two reasons, renal and extrarenal perturbations. For example, the increase of urinary lactate and alanine could be ascribed to toxicological impairment of the mitochondria in the liver or due to renal tubular injury, thus affecting renal reabsorption.42 Elevated urinary lactate and alanine, and decreased serum lactate and alanine confirmed the renal reabsorption impairment. Decreased levels of the TCA cycle intermediates in the urine have been observed in a series of studies on HgCl2-induced nephrotoxicity, and are due to toxin-induced effects on the key enzymes in the TCA cycle.43,44 The lesion of kidney is also indicated by the remarkable increase in urinary acetate.45 Thus, the decreased urinary levels of TCA cycle intermediates and serum lactate and alanine, and the increased urinary lactate, alanine, and acetate might demonstrate the nephrotoxicity induced by crude ricinine.
Urea is the principal end product of protein catabolism in the urea cycle, where L-aspartate and the amino groups donated by ammonia are converted to urea. Nitrogenous waste is produced mainly by protein catabolism and is removed in the form of urea from the body. Due to special anatomic-functional adaptations of the kidney, rodents normally have high urinary urea concentrations caused by an efficient urea concentration mechanism, which keeps the blood urea concentration low. Decreased levels of urinary urea and UUN and an increased level of BUN at week 8 in the HD rats indicated a dysfunction of the nitrogenous waste excretion, thus demonstrating renal damage induced by crude ricinine. Creatinine is a waste product formed by the slow spontaneous degradation of creatine-phosphate.46 A decrease in the urinary excretion of creatinine, accompanied with an increase of creatinine in serum, has also been observed in chronic renal failure, leading to filtration rate falls.47 This renal failure also happened in crude ricinine intoxicated rats, since the decreased creatinine in urine, increased the levels of creatinine in both serum and the kidney; increased SCR and decreased UCR in biochemistry were also observed in this study. Myo-inositol, one of the organic osmolytes, has been reported to be a renal marker of diabetic nephropathy, and to be one of the renal markers for the detection of renal tubular dysfunction induced by cadmium.48 Myo-inositol could be a sensitive indicator of impaired renal osmolyte activity caused by melamine and cyanuric acid and ochratoxin A induced toxicity.49,50 Notably, ricinine might inhibit two enzymes in the inositol pathway, namely, inositol poly-phosphate 1-phosphatase and inositol monophosphatase, leading to the depletion of inositol levels in tissues,51 which matches well with our results.
Decreased hippurate and increased TMA in urine and decreased TMAO in both urine and serum were observed in the HD rats. The decrease in hippurate has multifaceted reasons. Hippurate could be synthesized from benzoic acid in the kidney or liver. Its significant decrease in this study may thus be indicative of a metabolic alteration and, even more importantly, of an impairment in its secretion at the level of the proximal tubule.52 However, the level of hippurate has also been related to the microbial activity and micro floral composition of the colon.53,54 It could be produced from the degradation of shikimic acid (quinic acid) by intestinal microorganisms, and could also be synthesized from benzoic acid and phenyl acetic acid, two metabolites produced by bacterial metabolism.55,56 Also, the urinary level of hippurate is altered in animals exposed to drugs or foods with antimicrobial activity.57–59 Therefore, the decreased urinary level of hippurate in the crude-ricinine-treated rats may also mirror a disturbance of the gut microbiota, which is also supported by the significant decrease in TMAO and increase in TMA in the HD rats. TMAO is an aliphatic amine and the oxidation product of TMA through the action of gut microbes. It was reported that the concentration of urinary TMAO gradually increased over the time when germ-free rats were introduced into a normal environment, gradually establishing a stable gut microbiota.60
Conclusion
A 1H NMR based metabolomics approach complemented with histopathological inspection and biochemical assay has been developed to study the chronic toxic effects of crude ricinine in rats. Crude ricinine exhibited obvious nephrotoxicity and produced severe metabolic alterations, which were related with oxidative stress, energy metabolism, amino acid metabolism, renal function, and the gut bacteria system. This work provides a molecular basis for the chronic toxicity of crude ricinine and shows the power of a 1H NMR-based metabolomic approach to study the toxicity of drugs dynamically and systematically.Acknowledgements
This research work was financially supported by the Key Project of National Natural Science Foundation of China (81430092), NSFC grant 81173526, the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD), and the Program for Changjiang Scholars and Innovative Research Team in University (PCSIRT-IRT1193).References
- X. K. Xie, J. Kirby and J. D. Keasling, Phytochemistry, 2012, 78, 20–28 CrossRef CAS PubMed.
- G. R. Waller, E. K. Nowacki and K. Edmund, Alkaloid biology and metabolism in plants, Springer, 1978 Search PubMed.
- V. Bullangpoti, N. Khumrungsee, W. Pluempanupat, Y. Kainoh and U. Saguanpong, J. Pestic. Sci., 2011, 36, 260–263 CrossRef CAS PubMed.
- M. F. Bigi, V. L. Torkomian, S. T. De Groote, M. J. A. Hebling, O. C. Bueno, F. C. Pagnocca, J. B. Fernandes, P. C. Vieira and M. F. G. Da Silva, Pest Manage. Sci., 2004, 60, 933–938 CrossRef CAS PubMed.
- M. A. Ramos-López, S. Pérez, G. C. Rodríguez-Hernández, P. Guevara-Fefer and M. A. Zavala-Sánchez, Afr. J. Biotechnol., 2010, 9, 1359–1365 Search PubMed.
- J. Peng, S. Cai, L. Wang, N. Zhao, T. J. Zhang, Z. X. Chen and F. H. Meng, PLoS One, 2014, 9, e90416 Search PubMed.
- A. W. Nicholls, E. Holmes, J. C. Lindon, J. P. Shockcor, R. D. Farrant, J. N. Haselden, S. J. Damment, C. J. Waterfield and J. K. Nicholson, Chem. Res. Toxicol., 2001, 14, 975–987 CrossRef CAS PubMed.
- D. D. Wei, J. S. Wang, P. R. Wang, M. H. Li, M. H. Yang and L. Y. Kong, J. Pharm. Biomed. Anal., 2014, 98, 334–338 CrossRef CAS PubMed.
- D. D. Wei, G. Dong, P. P. Guo, J. S. Wang, M. H. Li, M. H. Yang and L. Y. Kong, Mol. BioSyst., 2014, 10, 2923–2934 RSC.
- J. K. Nicholson and I. D. Wilson, Nat. Rev. Drug Discovery, 2003, 2, 668–676 CrossRef CAS PubMed.
- J. R. Sheedy, Metabolomics Tools for Natural Product Discovery, Springer, 2013, pp. 81–97 Search PubMed.
- A. Craig, J. Sidaway, E. Holmes, T. Orton, D. Jackson, R. Rowlinson, J. Nickson, R. Tonge, I. Wilson and J. Nicholson, J. Proteome Res., 2006, 5, 1586–1601 CrossRef CAS PubMed.
- P. Li, L. P. Yang and Y. W. Gong, J. Tradit. Chin. Med., 2009, 29, 153–157 CrossRef.
- J. K. Nicholson, Mol. Syst. Biol., 2006, 2, 1–6 CrossRef PubMed.
- B. W. Tang, J. J. Ding, F. H. Wu, L. Chen, Y. X. Yang and F. Y. Song, J. Ethnopharmacol., 2012, 141, 134–142 CrossRef CAS PubMed.
- P. P. Guo, J. S. Wang, G. Dong, D. D Wei, M. H. Li, M. H. Yang and L. Y. Kong, Mol. BioSyst., 2014, 10, 2426–2440 RSC.
- G. D. Amico and C. Bazzi, Curr. Opin. Nephrol. Hypertens., 2003, 12, 639–643 CrossRef PubMed.
- M. A. Salem, S. A. El-Habashy, O. M. Saeid, M. M. El-Tawil and P. H. Tawfik, Pediatr. Diabetes, 2002, 3, 37–41 CrossRef PubMed.
- D. Finco and J. Duncan, J. Am. Vet. Med. Assoc., 1976, 168, 593–601 CAS.
- S. Cuzzocrea, D. P. Riley, A. P. Caputi and D. Salvemini, Pharmacol. Rev., 2001, 53, 135–159 CAS.
- S. J. Stohs and D. Bagchi, Free Radical Biol. Med., 1995, 18, 321–336 CrossRef CAS.
- O. Kumar, K. Sugendran and R. Vijayaraghavan, Toxicon, 2003, 41, 333–338 CrossRef CAS.
- C. Behl, J. B. Davis, R. Lesley and D. Schubert, Cell, 1994, 77, 817–827 CrossRef CAS.
- M. Valko, H. Morris and M. T. D. Cronin, Curr. Med. Chem., 2005, 12, 1161–1208 CrossRef CAS.
- S. M. Hussain, K. L. Hess, J. M. Gearhart, K. T. Geiss and J. J. Schlager, Toxicol. In Vitro, 2005, 19, 975–983 CrossRef CAS PubMed.
- M. Valko, D. Leibfritz, J. Moncol, M. T. D. Cronin, M. Mazur and J. Telser, Int. J. Biochem. Cell Biol., 2007, 39, 44–84 CrossRef CAS PubMed.
- P.-R. Wang, J.-S. Wang, M.-H. Yang and L.-Y. Kong, J. Pharm. Biomed. Anal., 2014, 88, 106–116 CrossRef CAS PubMed.
- M. Yang, S. Wang, F. Hao, Y. Li, H. Tang and X. Shi, Talanta, 2012, 88, 136–144 CrossRef CAS PubMed.
- J. Saric, J. V. Li, J. R. Swann, J. Utzinger, G. Calvert, J. K. Nicholson, S. Dirnhofer, M. J. Dallman, M. Bictash and E. Holmes, J. Proteome Res., 2010, 9, 2255–2264 CrossRef CAS PubMed.
- M. Bhatia and S. Moochhala, J. Pathol., 2004, 202, 145–156 CrossRef CAS PubMed.
- E. B. Kalaz, J. Çoban, A. F. Aydın, I. Doğan-Ekici, S. Doğru-Abbasoğlu, S. Öztezcan and M. Uysal, J. Physiol. Biochem., 2013, 1–11 Search PubMed.
- N. Hirakawa, R. Okauchi, Y. Miura and K. Yagasaki, Biosci., Biotechnol., Biochem., 2005, 69, 653–658 CrossRef CAS.
- C. Tohda, T. Kuboyama and K. Komatsu, Neurosignals, 2005, 14, 34–45 CrossRef CAS PubMed.
- B. N. Hong, T. H. Yi, S. Y. Kim and T. H. Kang, Biol. Pharm. Bull., 2009, 32, 597–603 CAS.
- O. Yoshinari, A. Takenake and K. Igarashi, J. Med. Food, 2013, 16, 34–41 CrossRef CAS PubMed.
- B. Soni, N. P. Visavadiya and D. Madamwar, Toxicology, 2008, 248, 59–65 CrossRef CAS PubMed.
- Y. Ohta, M. Kongo-Nishimura, Y. Imai and T. Kishikawa, J. Clin. Biochem. Nutr., 2003, 33, 83–93 CrossRef CAS.
- Y. P. Lin, D. Y. Si, Z. P. Zhang and C. X. Liu, Toxicology, 2009, 256, 191–200 CrossRef CAS PubMed.
- M. I. Shariff, A. I. Gomaa, I. J. Cox, M. Patel, H. R. Williams, M. M. Crossey, A. V. Thillainayagam, H. C. Thomas, I. Waked, S. A. Khan and S. D. Taylor-Robinson, J. Proteome Res., 2011, 10, 1828–1836 CrossRef CAS PubMed.
- J. D. Kopple, J. Nutr., 2007, 137, 1586S–1590S CAS.
- R. E. Williams and E. A. Lock, Toxicology, 2004, 201, 231–238 CrossRef CAS PubMed.
- C. F. Lu, Y. W. Wang, Z. G. Sheng, G. Liu, Z. Fu, J. Zhao, J. Zhao, X. Z. Yan, B. Z. Zhu and S. Q. Peng, Toxicol. Appl. Pharmacol., 2010, 248, 178–184 CrossRef CAS PubMed.
- J. K. Nicholson, J. A. Timbrell and P. J. Sadler, Mol. Pharmacol., 1985, 27, 644–651 CAS.
- E. Holmes, F. W. Bonner, K. P. R. Gartland and J. K. Nicholson, J. Pharm. Biomed. Anal., 1990, 8, 959–962 CrossRef CAS.
- K. P. Gartland, F. W. Bonner, J. A. Timbrell and J. K. Nicholson, Arch. Toxicol., 1989, 63, 97–106 CrossRef CAS.
- M. Wyss and R. Kaddurah-Daouk, Physiol. Rev., 2000, 80, 1107–1213 CAS.
- R. Goldman, Exp. Biol. Med., 1954, 85, 446–448 CrossRef CAS PubMed.
- D. Suvagandha, M. Nishijo, W. Swaddiwudhipong, R. Honda, M. Ohse, T. Kuhara, H. Nakagawa and W. Ruangyuttikarn, Int. J. Environ. Res. Public Health, 2014, 11, 3661–3677 CrossRef CAS PubMed.
- T. H. Kim, M. Y. Ahn, H. J. Lim, Y. J. Lee, Y. J. Shin, U. De, J. Lee, B. M. Lee, S. Kim and H. S. Kim, Arch. Toxicol., 2012, 86, 1885–1897 CrossRef CAS PubMed.
- M. Sieber, S. Wagner, E. Rached, A. Amberg, A. Mally and W. Dekant, Chem. Res. Toxicol., 2009, 22, 1221–1231 CrossRef CAS PubMed.
- P. E. Brandish, M. Su, D. J. Holder, P. Hodor, J. Szumiloski, R. R. Kleinhanz, J. E. Forbes, M. E. McWhorter, S. J. Duenwald and M. L. Parrish, Neuron, 2005, 45, 861–872 CrossRef CAS PubMed.
- C. Zuppi, I. Messana, F. Forni, C. Rossi, L. Pennacchietti, F. Ferrari and B. Giardina, Clin. Chim. Acta, 1997, 265, 85–97 CrossRef CAS.
- A. W. Nicholls, R. J. Mortishire-Smith and J. K. Nicholson, Chem. Res. Toxicol., 2003, 16, 1395–1404 CrossRef CAS PubMed.
- R. E. Williams, H. W. Eyton-Jones, M. J. Farnworth, R. Gallagher and W. M. Provan, Xenobiotica, 2002, 32, 783–794 CrossRef CAS PubMed.
- A. N. Phipps, J. Stewart, B. Wright and I. D. Wilson, Xenobiotica, 1998, 28, 527–537 CrossRef CAS PubMed.
- J. Delaney, W. A. Neville, A. Swain, A. Miles, M. S. Leonard and C. J. Waterfield, Biomarkers, 2004, 9, 271–290 CrossRef CAS PubMed.
- Y. L. Wang, H. R. Tang, J. K. Nicholson, P. J. Hylands, J. Sampson and E. Holmes, J. Agric. Food Chem., 2005, 53, 191–196 CrossRef CAS PubMed.
- C. A. Daykin, J. P. M. Van Duynhoven, A. Groenewegen, M. Dachtler, J. M. M. Van Amelsvoort and T. P. J. Mulder, J. Agric. Food Chem., 2005, 53, 1428–1434 CrossRef CAS PubMed.
- N. J. Waters, C. J. Waterfield, R. D. Farrant, E. Holmes and J. K. Nicholson, J. Proteome Res., 2006, 5, 1448–1459 CrossRef CAS PubMed.
- J. K. Nicholson and I. D. Wilson, Nat. Rev. Drug Discovery, 2003, 2, 668–676 CrossRef CAS PubMed.
Footnotes |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/c4ra14660c |
| ‡ These authors contributed equally to this work. |
| This journal is © The Royal Society of Chemistry 2015 |