
Physicochemical and cytotoxicity analysis of glycerol monoolein-based nanoparticles
Abstract
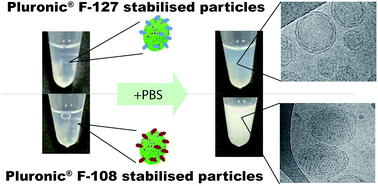
Lyotropic liquid crystalline dispersions, such as cubosomes, have been proposed as potential drug delivery vehicles. A recently described ‘salt induced’ method of cubosome production may be suitable for the encapsulation of macromolecular bioactive therapeutics, such as proteins, within the cubic phase. Here, we develop and characterise glycerol-monoolein (GMO)-based cubosomes using this novel method of cubosome production. Using the anionic biological lipid 1,2-dipalmitoyl phosphatidylserine (DPPS) to prevent GMO from forming its natural cubic-phase, we validate that addition of phosphate buffered saline (PBS) can be used to reverse the effects of DPPS. However, this transition is dependent on the type of Pluronic® block copolymer stabiliser used to prevent re-flocculation of the cubosome dispersions. Using small angle X-ray scattering (SAXS) and cryogenic transmission electron microscopy, we show that the ‘salt induced’ phase transition, from small unilamellar vesicles to cubosomes, is inhibited when using Pluronic® F127. In contrast, using the larger, more hydrophilic stabiliser Pluronic® F108, cubosomes can be formed, although further analysis using SAXS suggests these GMO-based cubosomes are less thermally stable than those comprising GMO alone. In addition, we find no significant difference in the in vitro cytotoxicity of cubosome dispersions formed using either of these stabilisers, or between those containing DPPS and those without. The ability to control cubic phase transitions may present an opportunity for the incorporation of therapeutically relevant proteins in these nanoparticles.
 Please wait while we load your content...
Please wait while we load your content...

