
Calcium mitigates the stress caused by ZnSO4 as a sulphur fertilizer and enhances the sulforaphane formation of broccoli sprouts†
Runqiang Yanga,
Liping Guoab,
Yulin Zhoua,
Chang Shena and
Zhenxin Gu*a
aCollege of Food Science and Technology, Nanjing Agricultural University, Nanjing, 210095, People's Republic of China. E-mail: guzx@njau.edu.cn; Fax: +86-25-84396293; Tel: +86-25-84396293
bCollege of Food Science and Engineering, Qingdao Agricultural University, Qingdao, Shandong 266109, People's Republic of China
First published on 6th January 2015
Abstract
In order to improve the growth condition of broccoli sprouts under ZnSO4 application, exogenous CaCl2 was added in the cultural medium, and the growth profiles, key bioactive substances (glucosinolates, sulforaphane, ascorbic acid and phenolic compounds), antioxidant capacity, myrosinase activity and related genes expression of the broccoli sprouts were evaluated. Results showed that the stressful condition caused by ZnSO4 was effectively mitigated by CaCl2. Consequently, ascorbic acid and the total phenolics content as well as the antioxidant capacity of the broccoli sprouts decreased compared with that of the sole ZnSO4 treatment. However, sulforaphane formation increased because a higher glucoraphanin content, myrosinase activity and related gene expression was induced after the CaCl2 treatment. The glucoraphanin content and sulforaphane formation of the water-treated sprouts decreased steadily during germination. However, the sulforaphane formation of the sprouts treated with ZnSO4 plus CaCl2 increased after germinating for 2 days. These results suggest that CaCl2 could mitigate the stress caused by ZnSO4 and enhance the sulforaphane formation of broccoli sprouts.
1 Introduction
Compared with their mature counterparts, broccoli sprouts are more globally consumed as a promising food because they are rich in health-promoting compounds, such as folic acid, phenolics, ascorbic acid and glucosinolates.1,2 Regular consumption of broccoli sprouts can reduce the risk of developing chronic diseases and different types of cancer.3 These beneficial effects are mainly due to the presence of glucosinolates, especially glucoraphanin, which accounts for over 50% of the total glucosinolate in most broccoli cultivars.4,5 Upon the disruption of plant tissues, glucoraphanin is hydrolyzed by myrosinase into sulforaphane. Sulforaphane has been proven to be a natural inducer of the phase II detoxification enzyme to detoxify cancer-causing chemicals.3 In plants, sulforaphane formation depends on the content of glucoraphanin biosynthesized by flavin-containing monooxygenase (FMO). In addition to being hydrolyzed by myrosinase, glucoraphanin also serves as a precursor for the biosynthesis of alkenyl-glucosinolates by AOP2.6 Furthermore, sulforaphane formation also depends on myrosinase and epithiospecifier protein (ESP) activity. High ESP activity is beneficial for sulforaphane nitrile formation. Hence, it is necessary to investigate these enzymes gene expression and sulforaphane formation in broccoli sprouts.Glucosinolates are the one type of sulphur-containing compound, which may be considered as a source of sulphur for the growth of sprout under low-sulphur conditions.7 Simultaneously, the biosynthesis of glucosinolates also needs sulphur.7 Therefore, sulphur has been applied for the growth of broccoli sprouts in order to accumulate glucosinolates.8,9 However, the exceeded application of sulphur would significantly decrease the yield of broccoli sprouts.8 In our previous study, we compared 2 mmol L−1 ZnSO4, K2SO4 and methionine (Met) on the growth and glucosinolates content of broccoli sprouts. The results showed that ZnSO4 increased the glucosinolate content; however, it significantly inhibited the growth of the broccoli sprouts (Fig. S1, ESI†). Therefore, methods to mitigate the stress caused by ZnSO4 and increase the yield of the sprouts are very important.
Calcium is known to be involved in the signal transduction in plants by binding with Ca2+-sensors such as calmodulin to induce appropriate physiological cellular responses.10 Moreover, Ca2+ plays an important role in maintaining the functional and structural integrity of cell membranes.11 Epstein12 clearly demonstrated that the addition of supplemental Ca2+ in the growth medium could enhance the salt tolerance of a plant. Therefore, it is hypothesized whether Ca2+ could mitigate the stress caused by ZnSO4 during the growth of broccoli sprouts and further improve the content of its bioactive substances.
In the present study, we attempted to add Ca2+ in the growth medium of broccoli sprouts parallel with ZnSO4 application. Subsequently, the growth profiles, key bioactive substances (glucosinolates, sulforaphane, ascorbic acid and phenolic compounds), antioxidant capacity, myrosinase activity and related gene expression of broccoli sprouts were evaluated. The changing patterns of glucoraphanin content, sulforaphane formation and myrosinase activity of germinating broccoli seeds and sprouts were also investigated.
2 Materials and methods
2.1 Materials and reagents
Broccoli seeds (B. oleracea L. var. italica) were purchased from Nanjing Jinshengda Seed Co. Ltd. (Jiangsu, China). The standard samples of sulforaphane and glucotropaeolin were purchased from Sigma Chemical Co., (St. Louis, MO, USA). Other chemicals and reagents were of analytical grade and were purchased from the Shanghai Institute of Biochemistry (Shanghai, China).2.2 Seed germination
Dry seeds were immersed in 1.5% sodium hypochlorite for 15 min, then drained and washed with distilled water until they reached neutral pH. They were then placed in distilled water and soaked at 30 °C for 4 h. The soaked seeds were germinated on a filter paper in petri dishes (15 cm in diameter) filled with sterilized quartz sand and they were left to grow in the incubators at 30 °C in darkness for 4 days. Sprout samples were rapidly and gently collected from the surface of the filter paper. Then the fresh weight (FW) of the broccoli sprouts was weighed, after which they were immediately frozen in liquid nitrogen and kept in polyethylene bags at −70 °C for further experiments.2.3 ZnSO4 and CaCl2 treatment
The seeds in each petri dish were supplied with 60 mL distilled water on the first day of germination and then every 12 h with 20 mL of the following testing solutions: (i) distilled water as the control (water); (ii) 4 mmol L−1 ZnSO4 (Zn); (iii) 11 mmol L−1 CaCl2 (Ca); (iv) 4 mmol L−1 ZnSO4 plus additional solution of 11 mmol L−1 CaCl2 (Zn + Ca). The ZnSO4 concentration of 4 mmol L−1 was pre-selected with the highest sulforaphane formation, which significantly inhibited the growth of sprouts (Fig. S2, ESI†). As for the CaCl2 concentration, a series of concentrations, including 0, 3, 5, 7, 9, 11, 13 and 15 mmol L−1 was carried out. Then, 11 mmol L−1 was chosen because it gave the highest sulforaphane formation in the sprouts and also increased the sprouts length under the stress of 4 mmol L−1 ZnSO4 (Fig. S3, ESI†).2.4 Sprout length measurement
Twenty sprouts were taken as one sample group. Their length was directly measured using a vernier caliper (Shanghai Precision Instruments Co., Ltd., Shanghai, China).2.5 Malondialdehyde (MDA) level determination
MDA was determined using the 2-thiobarbituric acid reaction method. Fresh sprouts (0.5 g) were homogenized with 5 mL of 5% trichloroacetic acid. The mixture was centrifuged at 10![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 000 rpm for 17 min (4 °C). The supernatant was used to determine MDA content as described by Yang et al.,13 and the MDA content was expressed as nmol g−1 FW.
000 rpm for 17 min (4 °C). The supernatant was used to determine MDA content as described by Yang et al.,13 and the MDA content was expressed as nmol g−1 FW.
2.6 Ascorbic acid content determination
Ascorbic acid content was determined according to Volden et al.14 Fresh broccoli sprouts (0.3 g) were extracted with 5 mL of 1.0% (w/v) oxalic acid. After centrifugation at 10000 rpm for 15 min, the supernatant was filtered through a 0.45 μm millipore before it was injected into an HPLC (Agilent 1200, USA), which had an Agilent 1200 tunable absorbance detector and a reversed-phase C18 column (4.6 × 250 mm, 5 μm, ZORBAX Eclips) and detected at 254 nm. Ascorbic acid was calculated by an external standard curve and expressed as mg per 100 g FW.
2.7 Total phenolics content determination
The total phenolics content of the sprouts (0.2 g) was extracted with 5.0 mL of 50% methanol. The mixture was centrifuged at 10000 rpm for 15 min. One mL of the supernatant was mixed with 1 mL of 0.2 mmol L−1 Folin–Ciocalteu reagent and 2.0 mL of 2% Na2CO3. The mixture was reacted at room temperature in the dark for 2 h. Its absorbance was read at 765 nm. Gallic acid was used as the standard. The results were expressed as mg of gallic acid equivalent (GAE) per 100 g FW.
2.8 Antioxidant capacity test
The sample extract was prepared in the same way as that used for total phenolics. The antioxidant capacity was evaluated using the T-AOC Kit (Nanjing Jiancheng Biotech Inc., Jiangsu, China). One unit of antioxidant capacity is defined as an increase in absorbance of 0.01 at 520 nm per min per gram at 37 °C of the reaction system.2.9 Glucosinolates analysis
Glucosinolates were extracted and analyzed as previously reported by Guo et al.15 Briefly, samples (500 mg) were extracted with 2 mL of 75% methanol at 80 °C for 15 min in a temperature-controlled water bath, with vortexing at regular intervals. The supernatant was collected after centrifugation (5 min, 10000 rpm), and the residues were extracted once again by using 2 mL of 75% methanol and then centrifuged. One milliliter of the combined supernatant was applied to a 1 mL DEAE Sephadex A-25 column (acetic acid activated) and rinsed with 2 mL of 0.02 mol L−1 sodium acetate. After the addition of 200 μL of arylsulfatase solution and incubation for 16 h at 35 °C, the desulphoglucoraphanin was eluted with 4 mL of Milli-Q water and filtered through a 0.45 μm membrane filter. Separation and detection were performed on an Agilent 1200 HPLC system (Agilent Technologies Co. Ltd., Palo Alto, CA, USA) equipped with a G1314B UV detector and a G1311A Quat pump, using an Eclipse XDB-C18 column (5 μm particle size, 4.6 × 150 mm; Agilent Technologies Co. Ltd.) at 226 nm. Determination was conducted at a flow-rate of 1.0 mL min−1 in a linear gradient starting with 0% acetonitrile for 1 min, reaching 20% acetonitrile at 21 min, and 0% acetonitrile at 26 min. Sinigrin (2-propenyl glucosinolate) (Sigma, St. Louis, MO, USA) was added to each sample as an internal standard before the first extraction. The total glucosinolate and glucoraphanin content was calculated and expressed as μmol g−1 fresh weight of broccoli sprouts.
2.10 Sulforaphane formation determination
Sulforaphane formation was measured by the method of Gu et al.16 with some modifications. Sprouts (0.2 g) were homogenized with 4.0 mL of distilled water. The mixture was hydrolyzed at 37 °C for 3 h in a shaking water bath. After the hydrolysis, it was extracted three times with 10 mL of ethyl acetate. The ethyl acetate fraction was dried at 37 °C under vacuum on a rotary evaporator. The concentrate was then dissolved in 2.0 mL of 10% acetonitrile, and filtered through a 0.45 μm membrane filter before it was injected into the HPLC. An Agilent 1200 HPLC system (Agilent Technologies Co. Ltd., Palo Alto, CA, USA) equipped with an Eclipse XDB-C18 column (5 μm particle size, 4.6 × 150 mm; Agilent Technologies Co. Ltd.) was used.2.11 Myrosinase activity determination
Myrosinase activity determination was carried out as described previously by Guo et al.15 The protein content of the supernatant was determined according to Bradford (1976) using bovine serum albumin as the standard. One myrosinase activity unit corresponded to 1.0 nmol glucose formed per minute. The specific activity was expressed as units per milligram of protein.2.12 Gene expression assay (QRT-PCR, Quantitative Real-Time PCR)
Total RNA from the broccoli sprouts was isolated using an E.Z.N.A.™ Plant RNA Kit (OMEGA, R6827-01). Approximately 2 μg total RNA was used as a template for the first strand cDNA synthesis, which was performed with an RT-PCR Kit (TaKaRa: DR027S). First-strand cDNA was synthesized as described previously.17 The PCR amplification was performed using TaKaRa Ex Taq™ polymerase for the target genes and Actin. For QRT-PCR analysis, the sequence-specific primers used are shown in Table 1. Triplicate quantitative assays were performed on each cDNA using the SYBR®Premix Ex Taq™ (TAKARA: RR420A) with the ABI 7500 sequence detection system according to the manufacturer's protocol (Applied Biosystems, Foster City, CA, USA). The PCR cycling conditions used were as follows: 1 cycle of 95 °C for 30 s, followed by 40 cycles of 95 °C for 3 s, 60 °C for 30 s.| Gene | Primer name | Primer sequences (5′ → 3′) |
|---|---|---|
| FMOGS-OX | Sense | TCGTCTTGGTGTCGTCGGTC |
| Ant-sense | ATCCTCATAAGCCTCCGCAAT | |
| AOP2 | Sense | GAGTAACGGAAAGAAAGAAGACAAGG |
| Ant-sense | ATAAGCGTGAAGAGTAGAACGAGGT | |
| MYR | Sense | AAGGTCATCAGGGAGAAGGGTG |
| Ant-sense | TGTTTGGCAGGGTTCTTAGTGG | |
| ESP | Sense | ACATTTGGGACCAGGGACG |
| Ant-sense | TTTCCATACACGGTGGCAGTC | |
| Actin | Sense | CTGTTCCAATCTACGAGGGTTTCT |
| Ant-sense | GCTCGGCTGTGGTGGTGAA |
2.13 Statistical analysis
Experimental data were expressed as the mean ± standard deviation (SD) of three replicates (n = 3) and subjected to statistical analysis with SPSS 18.0 (SPSS Inc., Chicago, IL). Means were performed by the Duncan's multiple-range tests. Differences at p < 0.05 were considered to be significant.3 Results and analysis
3.1 Growth profiles of broccoli sprouts
Images of the growth profiles of the 4 day-old broccoli sprouts and sprout lengths are presented in Fig. 1. ZnSO4 application significantly (p < 0.05) inhibited the sprouts growth and the shortest length obtained was 17.3 mm, whereas that of the water-treated sprout was 38.9 mm. After spraying supplementary Ca2+, the ZnSO4-induced stress was significantly mitigated, and the sprouts elongation was accelerated. Interestingly, the application of Ca2+ solely also increased the sprout length by 32.4% compared with the control. | ||
| Fig. 1 The growth profiles of broccoli sprouts grown under different treatments. Values are the means of triplicate analyses. Error bars show the standard deviation. Those with different lower case letters are significantly different at p < 0.05. After 4 day germination, the sprouts were collected and washed, and the sprout lengths were measured directly using a vernier caliper. | ||
3.2 Fresh weight and MDA content of broccoli sprouts
The fresh weight of the sprouts grown under ZnSO4 was the least, which indicates that the growth of the sprouts was markedly inhibited (p < 0.05) (Fig. 2A). Spraying only CaCl2 favored sprout elongation, whose fresh weight increased by 27.9% compared with the control. CaCl2 addition alleviated the ZnSO4 stress, generating an increment of 47.0% in fresh weight in comparison with the ZnSO4-treated sprouts. | ||
| Fig. 2 The fresh weight (A) and MDA content (B) of broccoli sprouts grown under different treatments. Values are the means of triplicate analyses. Error bars show the standard deviation. Those with different lower case letters are significantly different at p < 0.05. After 4 day germination, the sprouts were collected, washed and sipped up. The fresh weight and MDA level were immediately measured. | ||
MDA content exhibited the opposite trend as that of the fresh weight (Fig. 2B). The application of ZnSO4 led to the highest accumulation of MDA in the sprouts compared with the control. After adding CaCl2 based on the ZnSO4 treatment, the sprouts' MDA content decreased by 26.9%. Water and CaCl2 alone did not cause stress condition, where the MDA content of sprouts kept at a very low level.
3.3 Ascorbic acid, total phenolics content and antioxidant capacity of broccoli sprouts
The sprouts treated with ZnSO4 had the highest contents of ascorbic acid and total phenolics, followed by that of ZnSO4 plus CaCl2, water and CaCl2 alone (Fig. 3A and B). The CaCl2 treatment had no significant effect (p > 0.05) on the ascorbic acid content; however, it decreased the total phenolics as compared with the control. The contents of ascorbic acid and total phenolics of the broccoli sprouts grown under ZnSO4 were 3.17- and 3.91-fold of the control, respectively. Whereas, the additional CaCl2 treatment decreased their contents by 36.47% and 44.23% compared with the ZnSO4 treatment. | ||
| Fig. 3 Ascorbic acid content (A), total phenolics content (B) and antioxidant capacity (C) of broccoli sprouts grown under different treatments. Values are the means of triplicate analyses. Error bars show the standard deviation. Those with different lower case letters are significantly different at p < 0.05. After 4 day germination, the sprouts were collected, washed and sipped up. The ascorbic acid content, total phenolics content and antioxidant capacity of the fresh samples were immediately measured. | ||
As shown in Fig. 3C, the antioxidant capacity of the ZnSO4 treated sprouts was the highest, which was 2.55-fold of the control. Adding CaCl2 on the basis of the ZnSO4 treatment decreased the antioxidant capacity. Compared with the control, spraying CaCl2 alone also decreased the sprouts antioxidant capacity by 19.1%.
3.4 Glucosinolate content, sulforaphane formation and myrosinase activity of broccoli sprouts
Compared with the control (water), the CaCl2, ZnSO4 and ZnSO4 plus CaCl2 treatments significantly increased the total glucosinolate content in the broccoli sprouts (Fig. 4A). ZnSO4 led to the highest total glucosinolate content, which was 2.13-fold of the control. However, the ZnSO4 plus CaCl2 treatment decreased the total glucosinolate content compared with the ZnSO4 treatment. In both the hypocotyl and cotyledon, the total glucosinolate content in the ZnSO4 plus CaCl2 treated sprouts increased significantly by 1.01- and 0.43-fold, respectively, compared with the control (Fig. 4B). Similar to the total glucosinolate content, the CaCl2, ZnSO4 and ZnSO4 plus CaCl2 treatments all increased the glucoraphanin content, but the treatment of ZnSO4 plus CaCl2 favored best the accumulation of glucoraphanin (Fig. 4C). In both the hypocotyl and cotyledon, the glucoraphanin content in the ZnSO4 plus CaCl2 treated sprouts increased significantly by 1.59- and 0.55-fold, respectively, compared with the control (Fig. 4D).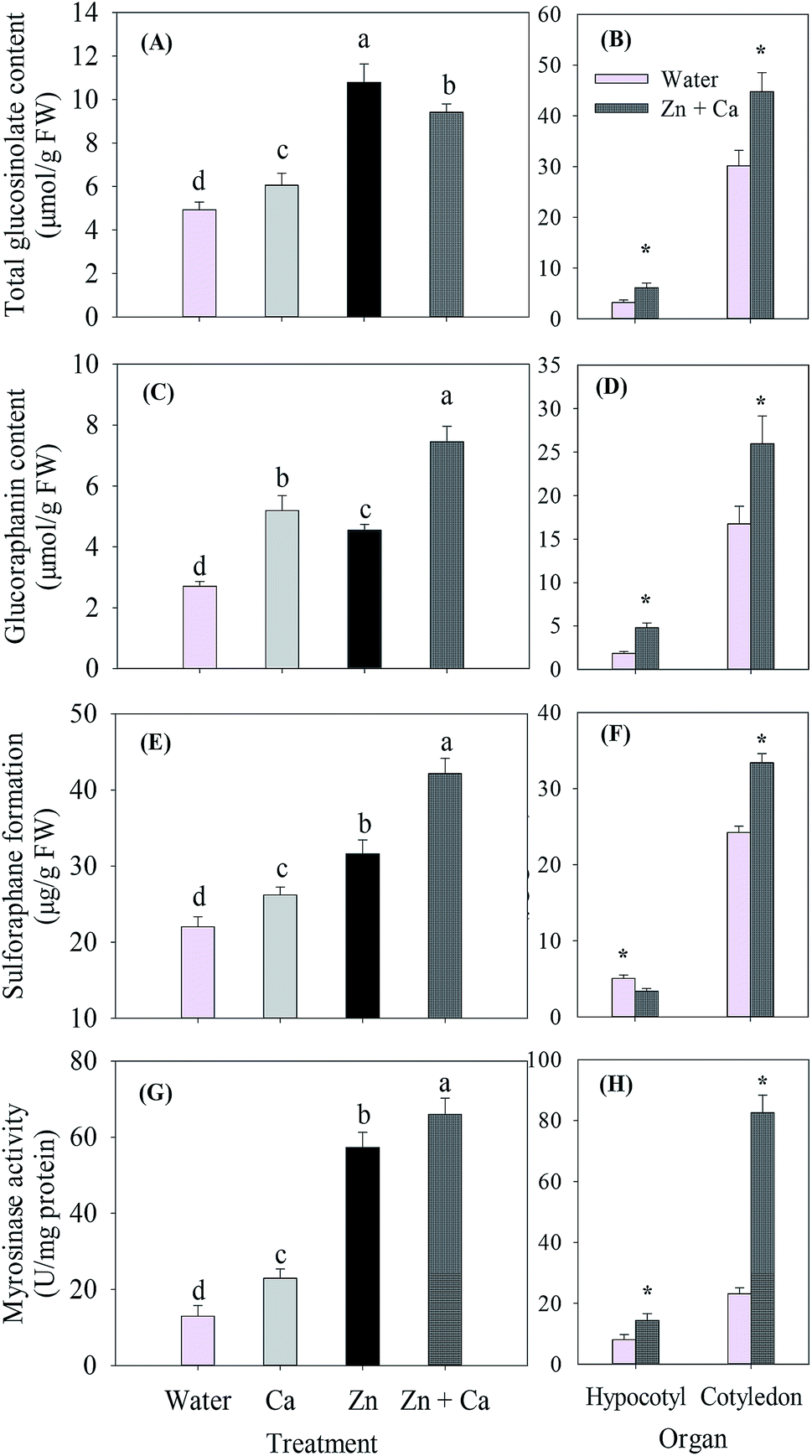 | ||
| Fig. 4 Total glucosinolate (A and B) and glucoraphanin (C and D) content, sulforaphane formation (E and F) and myrosinase activity (G and H) of broccoli sprouts under different treatments. Values are the means of triplicate analyses. Error bars show the standard deviation. Those with different lower case letters are significantly different at p < 0.05. * represents significant differences at p < 0.05. After 4 day germination, the sprouts were collected, washed and sipped up, then frozen in liquid nitrogen. The total glucosinolate and glucoraphanin content, sulforaphane formation and myrosinase activity were measured within 2 days. | ||
Compared with the control, treatments of CaCl2 alone, ZnSO4 and ZnSO4 plus CaCl2 were all beneficial for the formation of sulforaphane (Fig. 4E); moreover, they increased sulforaphane formation by 18.9%, 43.5% and 91.2%, respectively. However, ZnSO4 plus CaCl2 decreased sulforaphane formation in the hypocotyl by 33.62% and increased it in the cotyledon significantly compared with the control (Fig. 4F).
The myrosinase activity of the broccoli sprouts was enhanced by 3.40-fold under the ZnSO4 treatment in comparison with the control (Fig. 4G). Supplementary CaCl2 further stimulated the myrosinase activity by 15.3% compared with the ZnSO4 treatment. Spraying CaCl2 alone increased the activity by 75.9% compared with the control. Similar to the glucoraphanin content, myrosinase activity increased under the ZnSO4 plus CaCl2 treatment significantly and by 0.78 and 2.57-fold in the hypocotyl and cotyledon, respectively compared with the control (Fig. 4H).
3.5 Gene expression related to sulforaphane formation
The ZnSO4 and ZnSO4 plus CaCl2 treatments induced FMOGS-OX expressions, which were 2.48- and 2.67-fold of the control, respectively (Fig. 5A). However, they significantly decreased the AOP2 expression (Fig. 5B). CaCl2 addition significantly enhanced the expression of ESP (Fig. 5C). The ZnSO4 and ZnSO4 plus CaCl2 treatments all induced MYR expression, which was 1.97-, 4.12- and 9.45-fold of the control, respectively (Fig. 5D). | ||
| Fig. 5 The relative expression of FMOGS-OX (A), AOP2 (B), ESP (C) and MYR (D) of broccoli sprouts under different treatments. Each datum is the mean and standard deviation of three replicates per treatment. Values not sharing the same letter are significantly different at p < 0.05. | ||
After 4 day germination, the sprouts were collected, washed and sipped up and the total RNA immediately was extracted using a kit.
3.6 Changes in the glucoraphanin content, sulforaphane formation and myrosinase activity of broccoli sprouts during germination
Germination brought a significant (p < 0.05) decrease in the glucoraphanin content both in the control and the ZnSO4 plus CaCl2 treatment sprouts (Fig. 6A). The glucoraphanin content of the sprouts grown under water and ZnSO4 plus CaCl2 decreased by 96.5% and 90.4% within 4 days of germination, respectively. However, after germinating for 1 day, the glucoraphanin content of the ZnSO4 plus CaCl2 treated sprouts was significantly (p < 0.05) higher than that of the control at the same time point. | ||
| Fig. 6 Glucoraphanin content (A), sulforaphane formation (B) and myrosinase activity (C) of broccoli seeds and sprouts during different germination periods under the control and ZnSO4 plus CaCl2 treatments. Values are the means of triplicate analyses. Error bars show the standard deviation. * represents significant differences at p < 0.05. During germination, sprouts were sampled every day and frozen in liquid nitrogen. The glucoraphanin content, sulforaphane formation and myrosinase activity were measured within 2 days. | ||
Similar to the glucoraphanin content, the sulforaphane formation of the control sprouts decreased steadily with time elongation (Fig. 6B). Whereas, the ZnSO4 plus CaCl2 treatment led to a gradual increase in sulforaphane formation after 2 days of germination. No significant (p > 0.05) difference was observed between the control and ZnSO4 plus CaCl2 treatment in the first 2 days of germination. However, the sulforaphane formation of 3 day- and 4 day-old sprouts were significantly (p < 0.05) higher than that of the control.
The myrosinase activity of the sprouts sprayed with water increased on the 2nd day of germination and decreased afterwards (Fig. 6C). In contrast, the activity increased continuously upto the 3rd day of germination and remained stable when the sprouts were sprayed with ZnSO4 plus CaCl2. No significant (p > 0.05) change occurred on the first day of germination.
4 Discussion
The addition of CaCl2 to the seed germination medium under salinity stress have been reported to enhance the germination rate and seedling growth in wimmera ryegrass,18 cotton,19 tomato and cucumber plants.20 However, the concentration of CaCl2 should be clearly defined since lower or higher concentrations would have significantly different effects, which depend upon the salinity stress source and plant species or cultivars. In wimmera ryegrass, 5.0 mmol L−1 Ca2+ showed the best effect on NaCl stress, while 10.0 mmol L−1 Ca2+ was optimal for MgCl2 stress.18 Moreover, the author found that increasing the Ca2+ concentration upto 10.0 mmol L−1 under NaCl stress decreased the radicle length compared with that of 5.0 mmol L−1. In this study, it was also detected that a concentration higher than 11 mmol L−1 would decrease the broccoli sprout length (Fig. S2, ESI†). Apart from directly adding Ca2+ to the sprout growth medium, several studies chose to pre-treat seeds by soaking them in Ca2+ solutions.21,22 In both treatments they found that the salinity tolerance of the tested seeds was greatly enhanced. These results suggested that the mitigation effect of Ca2+ might also be achieved via soaking seeds prior to germination.A saline environment is usually created by NaCl treatment during sprouts growth.23,24 In broccoli sprouts cultivation, in order to enhance glucoraphanin biosynthesis, sulphur-containing chemicals are commonly supplied.7 However, since the sprouts are very salt sensitive,25 it is difficult to find the optimal sulphur concentration without influencing the sprouts growth. It has been reported that unsuitable concentrations of sodium thiosulphate8 and K2SO4 (ref. 25) inhibited sprouts growth and thus decreased the positive effects of sulphur fertilization. Herein, the stress was caused by spraying ZnSO4, which was originally expected to be a sulphur fertilizer. For one probable reason, the stress of ZnSO4 was due to the excessive level of Zn2+ because Zn is a micronutrient for sprout growth. The excessive Zn2+ was not utilized promptly and thus created an osmotic stress condition where the water absorption system was interrupted. Another reason might be that ZnSO4 presents a similar negative effect as NaCl does. Hence, towards different broccoli cultivars and growth media, the optimal sulphur type and concentration should be pre-selected depending on the sulphur uptake abilities of the sprouts and partitioning of sulphur into glucoraphanin.26
Ascorbic acid content and the activities of its regenerating enzymes in plants are highly related to environmental stresses such as light, temperature, salt, and drought.27 Nishikawa et al.28 investigated ascorbic acid metabolism in postharvest broccoli florets, and found that the ascorbate-regenerating system could be activated to protect plant organs from environmental stresses. In this study, the ascorbic acid content in broccoli sprouts was accelerated by ZnSO4 application, which could also be attributed to the activation of the regenerating enzymes under ZnSO4 stress.27 Phenolic compounds are secondary metabolites, which are produced when plants face environmental stresses.29 In the present study, the ZnSO4 application significantly promoted the biosynthesis of phenolic compounds as a result of possible osmotic stress (Fig. 3B). However, when the stress situation was mitigated likely by exogenous CaCl2, the total phenolics content of the broccoli sprouts decreased as compared with that of ZnSO4.
Ascorbic acid and phenolic compounds both have high antioxidant activities in plants,30,31 hence, the antioxidant capacity of plant tissues is closely associated with their contents.32 A significant positive correlation between ascorbic acid, phenolic compounds and antioxidant capacity was observed in radish33 and broccoli sprouts.5 In the present study, the antioxidant capacity of broccoli sprouts was tested with four different treatments had the same changing trend with the contents of ascorbic acid and phenolic compounds (Fig. 3). The highest antioxidant capacity was observed in the sprouts having the highest ascorbic acid and phenolic compounds content. Exogenous Ca2+ decreased the antioxidant capacity since the contents of ascorbic acid and phenolic compounds decreased.5 Barillari et al.34 reported that 4-methylthio-3-butenyl glucosinolate isolated from Raphanus sativus sprouts was related to its total antioxidant capacity. Guo et al.5 also suggested that glucosinolates in broccoli sprouts had some antioxidant capacity. However, in this study, no evident relations were observed between total antioxidant and total glucosinolates or glucoraphanin content (p > 0.05). The discrepancy could be attributed to the different cultivars of broccoli, in which the content of antioxidant components is different.
Falk et al.35 highlighted that sulphur fertilization could lead to an increase in glucosinolate content ranging from 0.25- to more than 50-fold, which relies on sulphur types and treating styles. Herein, as a sulphur-containing chemical, ZnSO4 increased the total glucosinolate content, but the ZnSO4 plus CaCl2 treatment decreased its content (Fig. 4A). Interestingly, the content of glucoraphanin was further enhanced by CaCl2 addition (Fig. 4C). Applying CaCl2 alone to sprouts increased the glucoraphanin content compared with that of the control. Moreover, it was noted that the glucoraphanin content of the sprouts under the sole CaCl2 treatment was higher than that of the sole ZnSO4 treatment. In addition, compared with the ZnSO4 treatment, the ZnSO4 plus CaCl2 treatment increased the gene expression related to glucoraphanin biosynthesis (Fig. 5A), but decreased AOP2 expression which is related to glucoraphanin degradation (Fig. 5B). These results indicated that CaCl2 could induce glucoraphanin biosynthesis in broccoli sprouts.
In both the ZnSO4 plus CaCl2 treated and the control sprouts, glucoraphanin content decreased steadily with germination time (Fig. 6A), which agreed with previous studies.2,36 The decrease in glucoraphanin content might result from the ‘dilution effects’ of water absorption during growth.37 Another possibility could be that glucoraphanin was degraded as the sulphur supply supports the sprouts tissue expansion and formation of other sulphur-containing compounds.7 Under the ZnSO4 plus CaCl2 treatment, the decrease of glucoraphanin was delayed to some extent, which was probably a result of the sulphur supply. Pérez-Balibrea et al.9 previously reported that the glucoraphanin content of broccoli sprouts treated with K2SO4 remained stable when they grew from 6 to 12 days.
Sulforaphane is hydrolyzed from glucoraphanin by myrosinase, whose formation largely depends on both glucoraphanin content and myrosinase activity.7 In this study, the sulforaphane formation of broccoli sprouts treated with ZnSO4 plus CaCl2 was the highest (Fig. 4D). This might be due to the higher glucoraphanin content (Fig. 4B), myrosinase activity (Fig. 4E) and its higher expression (Fig. 5D). Although the glucoraphanin content of the sprouts under the sole CaCl2 treatment was higher than that of sole ZnSO4, its sulforaphane formation was less because of the lower myrosinase activity (Fig. 4E) and its lower expression (Fig. 5D) as well as the higher expression of ESP (Fig. 5C). Liang et al.38 reported that Zn2+ could enhance myrosinase activity and thus increase sulforaphane formation. The findings in the present study are similar to theirs. It was clearly noticed that the glucoraphanin content of the broccoli sprouts under the ZnSO4 plus CaCl2 and the control treatments both decreased steadily during germination (Fig. 6A). However, the sulforaphane formation of the sprouts treated with ZnSO4 plus CaCl2 did not decrease steadily as expected (Fig. 6B). In contrast, its content increased after germinating for 2 days. This could be due to the fact that the myrosinase activity increased to a higher level after 1 day of germination. These results suggested that the glucoraphanin–myrosinase system is complicated and needs further investigation.
5 Conclusion
CaCl2 application effectively mitigated stressful condition caused by ZnSO4. Consequently, the antioxidant compounds, such as ascorbic acid and total phenolics content decreased. However, sulforaphane formation increased because higher glucoraphanin content, myrosinase activity and related genes expression were induced after the CaCl2 treatment.Conflicts of interest
The authors declare no conflict of interest.Acknowledgements
We are grateful for the financial support for this study from the Natural Science Foundation of China (Grant no. 31271912). The present study was also a project funded by the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD).References
- Q. Guo, L. Guo, Z. Wang, Y. Zhuang and Z. Gu, Food Chem., 2013, 141, 1580–1586 CrossRef CAS PubMed.
- S. Pérez-Balibrea, D. A. Moreno and C. García-Viguera, Food Chem., 2011, 125, 348–354 CrossRef PubMed.
- J. W. Fahey, Y. S. Zhang and P. Talalay, Proc. Natl. Acad. Sci. U. S. A., 1997, 94, 10367–10372 CrossRef CAS.
- L. Guo, R. Yang, Z. Wang, Q. Guo and Z. Gu, Int. J. Food Sci. Nutr., 2013, 65, 476–481 CrossRef PubMed.
- R. Guo, G. Yuan and Q. Wang, Sci. Hortic., 2011, 128, 159–165 CrossRef CAS PubMed.
- Y.-X. Zang, H. U. Kim, J. A. Kim, M.-H. Lim, M. Jin, S. C. Lee, S.-J. Kwon, S.-I. Lee, J. K. Hong and T.-H. Park, FEBS J., 2009, 276, 3559–3574 CrossRef CAS PubMed.
- Z. Gu, Q. Guo and Y. Gu, J. Integr. Agric., 2012, 11, 1804–1816 CrossRef CAS.
- R. M. Kestwal, J. C. Lin, D. Bagal-Kestwal and B. H. Chiang, Food Chem., 2011, 126, 1164–1171 CrossRef CAS PubMed.
- S. Pérez-Balibrea, D. A. Moreno and C. García-Viguera, J. Food Sci., 2010, 75, C673–C677 CrossRef PubMed.
- M. C. Kim, W. S. Chung, D.-J. Yun and M. J. Cho, Mol. Plant, 2009, 2, 13–21 CrossRef CAS PubMed.
- Z. Rengel, Plant, Cell Environ., 1992, 15, 625–632 CrossRef CAS PubMed.
- E. Epstein, Science, 1998, 280, 1906–1907 CrossRef CAS.
- Z. F. Yang, Y. H. Zheng and S. F. Cao, J. Agric. Food Chem., 2009, 57, 176–181 CrossRef CAS PubMed.
- J. Volden, G. B. Bengtsson and T. Wicklund, Food Chem., 2009, 112, 967–976 CrossRef CAS PubMed.
- L. Guo, R. Yang, Z. Wang, Q. Guo and Z. Gu, J. Funct. Foods, 2014, 9, 70–77 CrossRef CAS PubMed.
- Y. Gu, Q. Guo, L. Zhang, Z. Chen, Y. Han and Z. Gu, J. Agric. Food Chem., 2012, 60, 209–213 CrossRef CAS PubMed.
- B. Z. Li, W. J. Xin, S. B. Sun, Q. R. Shen and G. H. Xu, Plant Soil, 2006, 287, 145–159 CrossRef CAS.
- N. E. Marcar, Plant Soil, 1986, 93, 129–132 CrossRef CAS.
- L. Kent and A. Läuchli, Plant, Cell Environ., 1985, 8, 155–159 CrossRef CAS PubMed.
- A. R. Al-Harbi, J. Plant Nutr., 1995, 18, 1403–1416 CrossRef CAS.
- I. Chaudhuri and H. Wiebe, Plant Soil, 1968, 28, 208–216 CrossRef CAS.
- S. Rehman, P. Harris and W. Bourne, J. Plant Nutr., 1998, 21, 277–285 CrossRef CAS.
- F. V. A. Guimarães, C. F. de Lacerda, E. C. Marques, M. R. A. de Miranda, C. E. B. de Abreu, J. T. Prisco and E. Gomes-Filho, Plant Growth Regul., 2011, 65, 55–63 CrossRef PubMed.
- G. Kumar, R. S. Purty, M. P. Sharma, S. L. Singla-Pareek and A. Pareek, J. Plant Physiol., 2009, 166, 507–520 CrossRef CAS PubMed.
- A. Aires, E. Rosa and R. Carvalho, J. Sci. Food Agric., 2006, 86, 1512–1516 CrossRef CAS.
- N. Rangkadilok, M. E. Nicolas, R. N. Bennett, D. R. Eagling, R. R. Premier and P. W. J. Taylor, J. Agric. Food Chem., 2004, 52, 2632–2639 CrossRef CAS PubMed.
- M. W. Davey, M. Van Montagu, D. Inze, M. Sanmartin, A. Kanellis, N. Smirnoff, I. J. J. Benzie, J. J. Strain, D. Favell and J. Fletcher, J. Sci. Food Agric., 2000, 80, 825–860 CrossRef CAS.
- F. Nishikawa, M. Kato, H. Hyodo, Y. Ikoma, M. Sugiura and M. Yano, J. Exp. Bot., 2003, 54, 2439–2448 CrossRef CAS PubMed.
- C. Rice-Evans, N. Miller and G. Paganga, Trends Plant Sci., 1997, 2, 152–159 CrossRef.
- V. Valpuesta and M. A. Botella, Trends Plant Sci., 2004, 9, 573–577 CrossRef CAS PubMed.
- M. P. Kähkönen, A. I. Hopia, H. J. Vuorela, J.-P. Rauha, K. Pihlaja, T. S. Kujala and M. Heinonen, J. Agric. Food Chem., 1999, 47, 3954–3962 CrossRef PubMed.
- N. J. Miller and C. A. Rice-Evans, Food Chem., 1997, 60, 331–337 CrossRef CAS.
- H. J. Kim, F. Chen, X. Wang and J. H. Choi, J. Agric. Food Chem., 2006, 54, 7263–7269 CrossRef CAS PubMed.
- J. Barillari, R. Cervellati, M. Paolini, A. Tatibouët, P. Rollin and R. Iori, J. Agric. Food Chem., 2005, 53, 9890–9896 CrossRef CAS PubMed.
- K. L. Falk, J. G. Tokuhisa and J. Gershenzon, Plant Biol., 2007, 9, 573–581 CrossRef CAS PubMed.
- R.-f. Guo, G.-f. Yuan and Q.-m. Wang, J. Zhejiang Univ., Sci., B, 2013, 14, 124–131 CrossRef CAS PubMed.
- R. N. Bennett, E. A. Rosa, F. A. Mellon and P. A. Kroon, J. Agric. Food Chem., 2006, 54, 4005–4015 CrossRef CAS PubMed.
- H. Liang, Q. Yuan and Q. Xiao, J. Mol. Catal. B: Enzym., 2006, 43, 19–22 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/c4ra11371c |
| This journal is © The Royal Society of Chemistry 2015 |