DOI:
10.1039/C4RA06879C
(Paper)
RSC Adv., 2015,
5, 12338-12345
Assembly of plasmid DNA with pyrene-amines cationic amphiphiles into nanoparticles and their visible lysosome localization†
Received
9th July 2014
, Accepted 23rd December 2014
First published on 23rd December 2014
Abstract
In this study, we constructed a visible model for drug/gene dual delivery. Firstly, we prepared a series of pyrene fluorophore bearing cationic amphiphiles (Py-amines), which could be further employed as amphiphilic gene carriers and antitumor polyamine drug models. Then, the Py-amines fluorescent amphiphiles were utilized to bind/load plasmid DNA via electrostatic interactions to form nano-sized particles in aqueous solution. The average size, zeta potential and morphology of the self-assembled Py-amines/pDNA complexes were found to be largely dependent on the molecular structures of Py-amines amphiphiles. Moreover, the Py-amines and their pDNA complexes showed an evident cell proliferation inhibition capability in H1299 (human lung cancer) cells. Notably, the lysosomal localization of the Py-amines/pDNA complexes could be directly visualized using fluorescence microscopy in vitro. In summary, this current study could provide new and visible approach to design polyamine-based anti-tumor drug/plasmid DNA dual delivery systems, which could facilitate a greater understanding of the intracellular trafficking/localization of polyamine-based cationic gene/drug payloads.
Introduction
In recent years, nano-scaled functional delivery systems have been developed and employed as potential tools for the efficient transportation of drug/gene substances toward disease treatment.1,2 In these delivery systems, various amine-containing cationic building blocks were employed as positively charged moieties to bind/load negatively charged gene substances via electrostatic forces. Among them, polyamines (such as spermine2 and spermidine3) have often been employed as functional moieties in the construction of gene carriers, due to their relatively high positive charge density bringing about highly efficient DNA binding/loading affinities. Moreover, polyamine-bearing gene carriers could be easily protonated and lead to intracellular “lysosomal escape” effects,4,5 which is regarded as an efficient pathway for enhancing the gene transfection capability. To date, the development of new lipid-based polyamine conjugates with the merit of well-defined and controllable structures as gene delivery carriers has attracted increasing attention, e.g., Bhattacharya et al. disclosed that some cholesterol-conjugated low molecular weight polyethyleneimine (Cho-OEI)6 lipids and diammonium containing Gemini-lipids7–9 possessed remarkably high gene transfection efficiencies, even beyond the “gold standard” transfection agent bPEI-25K. Yang et al.10 developed 1,12-diaminododecane-polyamine cationic bolaamphiphiles as gene carriers and found these simple-structured bolaamphiphiles possessed highly efficient pDNA transfection capabilities. Yu et al.11 synthesized some cyclic polyamine-conjugated lipids, and revealed that they have high DNA binding affinities, and demonstrated proton buffering effects and efficient luciferase gene transfection behaviors. Although several polyamine-based cationic lipids have been developed as efficient gene carriers, their intracellular trafficking and distribution are still not well known due to the lack of optical labeling/imaging moieties. To date, the intracellular localization behaviors of polyamine-containing cationic gene carriers have only been observed on some fluorescent-labeled polyamine-containing cationic polymers.12,13 Therefore, to gain a better understanding of the intracellular fate of polyamine-conjugated functional lipid gene carriers, the development of fluorophore-bearing polyamine cationic lipids for “visible intracellular imaging” is highly desired.
On the other hand, in addition to the contribution of the polyamines in the construction of cationic gene carriers, recently, it has been revealed that some polyamine containing amphiphiles or lipids participate in cell metabolism and tumor proliferation,14,15 in which the polyamine moiety plays a vital role as a “pharmacophore”. For example, some polyamine–anthracene conjugates and/or their analogs16–18 were shown to possess evident anti-tumor activities through the inhibition of the polyamine transporter (PAT), resulting in tumor cell apoptosis. Mojzis et al.19 found that polyamine–naphthalimides conjugates have an evident anti-proliferative effect in MCF-7 and A549 cells through a possible non-specific DNA binding mechanism. Wang et al.20 revealed that some triamine-modified anthraquinones could target lysosome and lead to a possible mitochondria-mediated cell apoptotic death, and the lysosome targeting behavior could be indirectly observed by Lysotracker-red and Hochest 33342 staining. These studies revealed that the anti-tumor mechanisms of the polyamine-bearing amphiphiles greatly rely on their molecular structures (such as the aromatic-ring, charge number, linker, stereo-configuration) and demonstrated that the polyamine-containing amphiphiles or lipids could be employed as latent anti-tumor agents for cancer treatment. Oupický et al.21 recently prepared several bisethylnorspermine-conjugates as cationic lipid prodrugs, which could be utilized for gene/drug dual delivery in synergistic breast cancer treatment, indicating that the polyamine-based gene/drug dual delivery system could be further developed as a potential approach for multi-channel cancer therapy. Moreover, for further investigation on the intracellular fate of polyamine-based gene/drug dual delivery systems, the development of a convenient, easily manipulated, efficient and in situ optical labeling/imaging techniques22 is required.
Herein, we designed and synthesized a series of pyrene fluorophore bearing cationic polyamine amphiphiles (Py-amines) as visible models for drug/gene dual delivery, in which Py-amines could bind with pDNA via positive–negative electrostatic interactions; moreover, the interaction of the Py-amines and pDNA were determined by EB displacement and DNA gel-retardation assays, and the average particle size, surface potential and shape/morphology of the Py-amines/pDNA complexes were analyzed by a dynamic laser scattering instrument (DLS) and atomic force microscope (AFM), respectively. Furthermore, MTT cytotoxicity assay and fluorescence microscopy imaging were employed to explore the possibility of drug/gene dual delivery using the Py-amines/pDNA complexes.
Experimental section
Materials
Pyrene (97.5%) and polyethyleneimine (Mw = 800 Da, PEI-800) were purchased from Sigma-Aldrich and were utilized as-received. Boc-protected amine-containing building blocks were prepared according to previously reported studies23. Dicyclohexylcarbodiimide (DCC, 99.0%), 4-dimethylaminopyridine (DMAP, 98.0%) and trifluoroacetic acid (TFA, 97.0%) were purchased from Shanghai Sinopharm Chemical Reagent Co. Ltd (Shanghai, China) and were utilized as-received. PCS plasmid DNA (pDNA, 1.0 mg mL−1) and the H1299 cell lines were generously gifted by Prof. Yuhong Xu from Shanghai Jiaotong University, and Lysotracker-red was gifted by Dr Hua Zhu from Peking University. 96-well microplates and 50 mL cell cultivation flasks were purchased from Corning Co. Ltd. 0.01 M phosphate buffer solution (PBS), Dulbecco's modified Eagle's medium (DMEM) and 10% FBS were supplied by Hangzhou Genom Co. Ltd. In this study, all the other reagents and organic solvents were of analytical grade and were utilized without further purification. Moreover, the chemical synthesis routes and structural characterization of the newly synthesized Py-amines cationic amphiphiles are described in detail in the ESI (S1†).
NMR and MS measurements
1H NMR spectra were recorded at room temperature on a Varian VXR 300 FT-NMR Fourier transform NMR spectrometer instrument, operating at 300.0 MHz for the 1H nuclei, whereas 13C NMR spectra were characterized under room temperature on a Bruker Avance 300 NMR spectrometer, operating at 75.0 MHz for the 13C nuclei with tetramethylsilane (TMS) as the internal chemical shift reference. The mass spectra (ESI-MS) were measured on a Varian SATURN 2000 spectrometer.
Ethidium-bromide (EB) dye displacement assay24
EB (5.0 mg) and pDNA (PCS plasmid DNA, 5.0 mg) were mixed in 1 mL of 1× PBS buffer solution (0.01 M, pH = 7.4). Then, the mixture solution was further incubated for 2 min at room temperature, and the newly prepared Py-amines solution under a calculated N/P charge ratio (N/P ratio of 3–30) was placed into the as-prepared pDNA/EB mixture, and continuously incubated at ambient temperature for 2 min. Then, the fluorescence spectra were measured on a Hitachi F-7000 fluorescence spectrometer under excitation at 510 nm and an emission at 590 nm. The relative fluorescence was measured and calculated according to our prior study.25
Agarose gel retardation assay
The pDNA binding affinity of the Py-amines amphiphiles was further analyzed by an agarose gel retardation assay.26 Before the assay, the Py-amines samples were preliminarily dissolved in pure water to prepare the solution with the concentration of 3.3 × 10−3 M, and then the Py-amines/pDNA complexes were prepared by mixing pDNA (1 μg) with a predetermined amount of Py-amines under a preset N/P charge ratio in 50 μL 1× PBS (0.01 M, pH = 7.4) buffer solution. After incubation at 37 °C for 30 min, the complex solution was loaded onto a 1% agarose gel containing 0.5 μg mL−1 ethidium bromide dye (EB). Then, the gel electrophoresis was conducted in 1× TAE running buffer under 100 mV for 40 minutes, and the retardant in pDNA migration was thus recorded on a UVP benchtop 2UV transilluminator system.
Particle size and surface charge of the Py-amines/pDNA complexes
The average particle size and surface charge of the Py-amines/pDNA complexes in aqueous solution were characterized at room temperature on a Malvern Zetasizer Nano ZS90 dynamic light scattering instrument with λ = 633 nm at a fixed scattering angle of 90° (UK).27 The Py-amines samples were preliminarily dissolved in pure water to prepare the solution with concentration of 3.3 × 10−3 M, and then each sample was mixed with pDNA (5.0 μg) in 1 mL water under a predetermined N/P charge ratio and then maintained under incubation at 37 °C for 20 min. After that, the average particle size and surface charge of the as-prepared Py-amines/pDNA complexes were analyzed on a Malvern Zetasizer Nano ZS90 dynamic light scattering instrument.
Morphology of the Py-amines/pDNA complexes by atomic force microscopy
The Py-amines/pDNA complex morphology measurements were conducted at room temperature on a Nanoscope IVa atomic force microscope (AFM, Veeco Instrument) in tapping mode with an Olympus AC160TS cantilever (frequency: 300 kHz, stiffness: 42 N m−1).28 The Py-amines/pDNA complexes solution were first prepared by mixing Py-amines (3.3 × 10−3 M) with pDNA (5.0 μg) under a N/P charge ratio of 20 in 1 mL of pure water, and then the complexes solution were preliminarily dropped onto fresh cleaved mica and air-dried at room temperature prior to the AFM measurements.
Proton buffering capacity measured by acid–base titration
The proton buffering capacity for the newly synthesized Py-amines in aqueous solution was measured by acid–base titration on a Schott Titroline Easy instrument at room temperature29 with NaCl (0.1 M, negative control) and PEI-800 (polyethyleneimine (800 Da) as a positive control) as references. The cationic Py-amines and PEI-800 were separately dissolved in distilled water to prepare the solution with total amine concentration of 0.01 M, and then 1.0 M NaOH was added to set the initial pH to 11.0–11.5, and then the solution was gradually titrated with 0.1 M HCl solution. The proton buffering curves were measured manually on a Schott Titroline Easy instrument.
MTT cytotoxicity assay of the Py-amines and Py-amines/pDNA complexes
MTT assays30 were conducted with H1299 cell lines in order to evaluate the cytotoxicities of the Py-amines as well as their pDNA lipoplexes. Firstly, the cells were seeded into 96-well microplates with 5 × 103 cells per well in 100 μL RPMI-1640 medium (10% FBS added) and cultivated under 37 °C and 5% CO2 for 24 h. Subsequently, the medium was replaced with fresh RPMI-1640 (10% FBS), and then the Py-amines and Py-amines/pDNA complex under various concentrations (N/P charge ratios) were individually added into the wells and further incubated for another 24 h. Then, 20 μL of MTT (5.0 mg mL−1) was added into each well and kept under incubation for 4 h. After removing the medium, DMSO (100 μL per well) was added to dissolve the formed MTT formazan. Finally, with a gentle shaking of the microplates for 10 min in order to dissolve the formazan, each sample with six replicates (n = 6) was analyzed on a microplate reader (BioTek, ELX800, USA) at λabs = 490 nm (λabs = 630 nm as reference wavelength).
Intracellular localization of the Py-amines/pDNA complexes
H1299 and COS-7 cells were seeded into 24-well microplates (4 × 105 cells per well in 1 mL DMEM medium with 10% FBS) and incubated at 37 °C under 5% CO2 for 24 h. Then, the Py-amines/pDNA complexes were added into each well at a N/P ratio of 30 and washed with 1× PBS three times in order to eliminate the fluorescence background after 4 h incubation. Subsequently, the cells were stained with Lysotracker-red (250 ng mL−1) for 15 min to localize the position of the lysosome and washed with 1× PBS three times. Finally, the fluorescent images were observed and recorded on a Nikon Ti–S invert fluorescence microscope.
Results and discussion
Synthesis of Py-amines amphiphiles
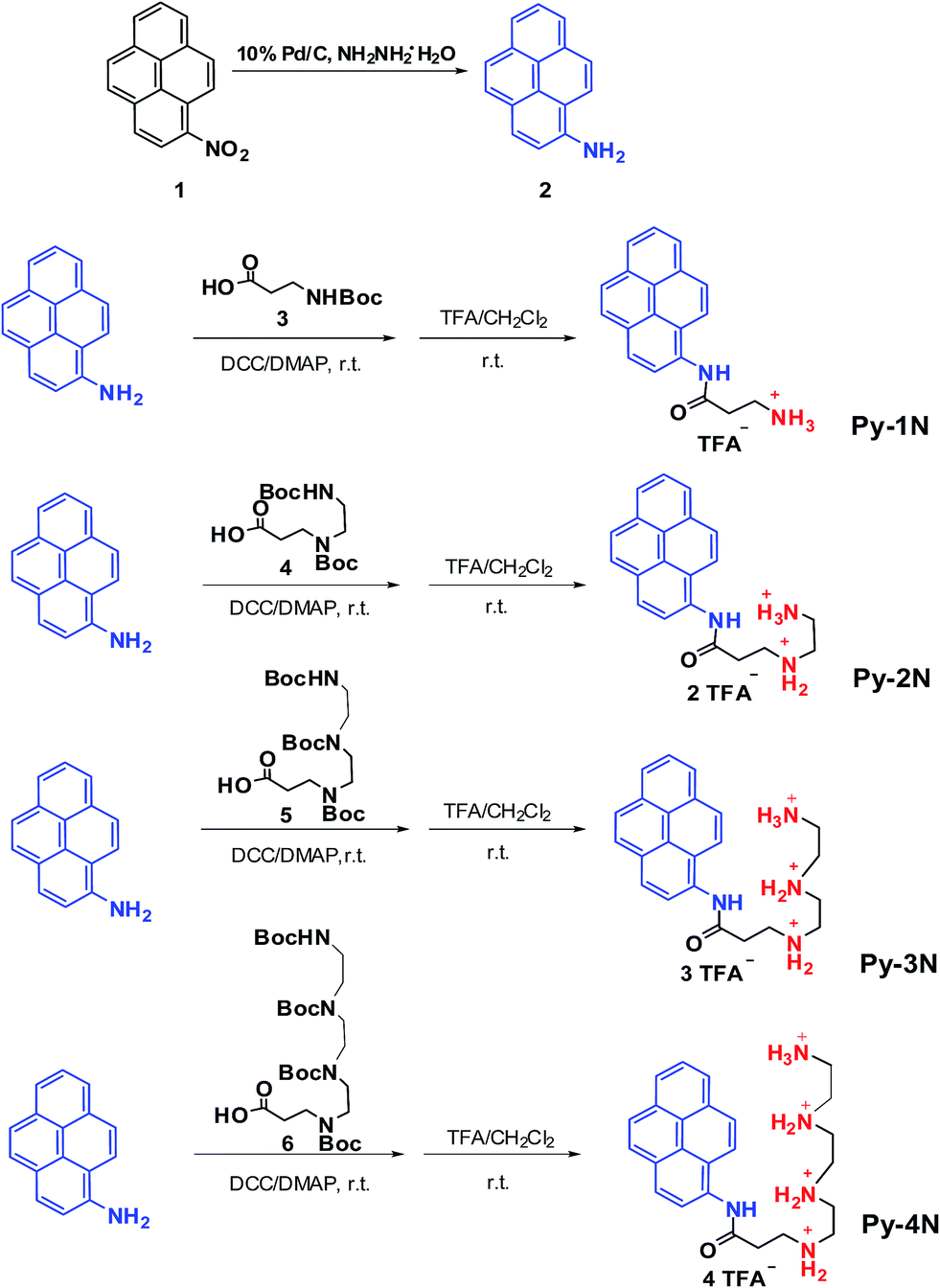
Facile, efficient and modular synthetic methods were acquired for the diversely oriented preparation of bioactive substances and pharmaceutics.31 In this study, a precursor compound of 1-amino-pyrene (2) was synthesized by catalyzed reduction (NH2NH2·H2O with 10% Pd/C as the catalyst) of 1-nitropyrene (1)25 and served as a fluorophore/hydrophobe dual functional moiety. Then, the as-prepared (2) was coupled with various Boc-protected amine/polyamine-containing building blocks (3, 4, 5, 6) under the catalysis of DCC/DMAP. Then, the Boc protection groups were removed in excess trifluoroacetic acid at room temperature to obtain a series of pyrene-derived cationic amphiphiles (Py-amines) bearing different numbers of amino groups as the final products, with the isolation yields of 50–65%. The amphiphiles were denoted as Py-1N (monoamine), Py-2N (diamines), Py-3N (triamines) and Py-4N (tetramines), respectively, and their synthetic routes are depicted in Fig. 1. The synthesized Py-amines cationic amphiphiles and important intermediates were characterized by NMR, MS and FTIR (see ESI, S1†). The results indicated that the Py-amines cationic amphiphiles were successfully prepared via facile, efficient and modular synthetic approaches.
 |
| | Fig. 1 Synthesis routes and chemical structures of the Py-amines cationic amphiphiles. | |
pDNA binding affinity of the Py-amines (EB and DNA-retardation assays)
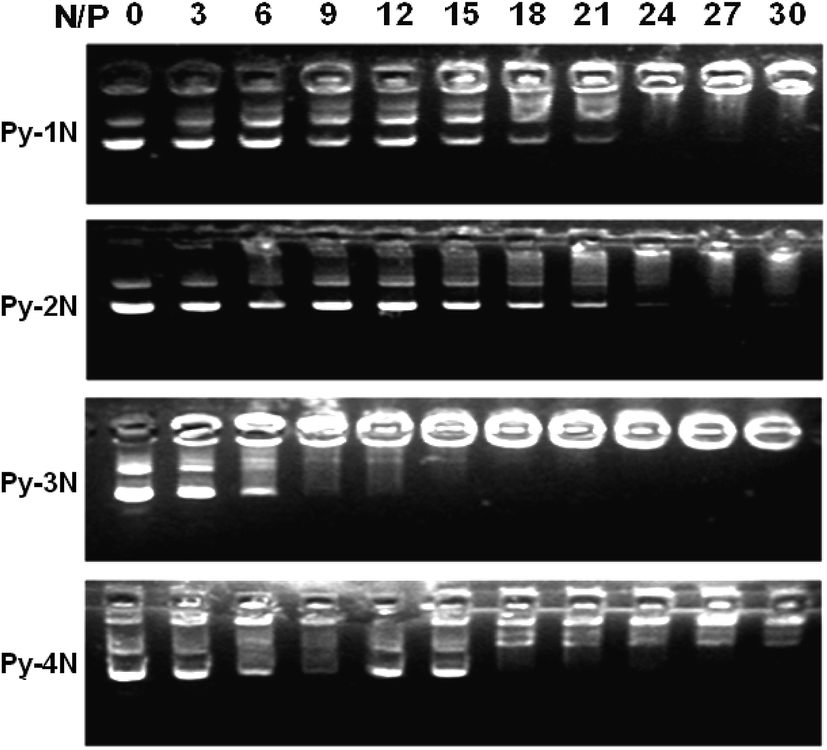
In order to examine the gene binding/loading capability of the Py-amines cationic amphiphiles, ethidium bromide (EB) displacement32 and DNA gel-retardation assays28 were employed. As shown in Fig. 2, it could be found that the relative fluorescence intensities of EB decreased gradually with the increasing amount of Py-amines (N/P from 0 to 30). Among them, Py-3N (BC50 = 3.3) showed a higher pDNA binding affinity than that of Py-1N (BC50 = 7.5) and Py-2N (BC50 = 5.1), indicating that the increasing positive charge number in cationic amphiphiles could enhance the pDNA binding affinity; however, lower binding affinity was observed on Py-4N (BC50 = 4.2), which might due to the longer polyamine chains decreasing the hydrophobicity of the cationic amphiphile.23 Furthermore, the pDNA binding capabilities of the Py-amines amphiphiles were examined using a pDNA retardation assay. As shown in Fig. 3, it could be seen that Py-1N and Py-2N inhibited the migration of pDNA at an N/P ratio of 21–24, Py-3N showed higher pDNA retardation capability at a N/P ratio of 12–15, and Py-4N showed lower pDNA retardation efficiency at a N/P ratio of 18–21). These results demonstrated that the number of amino groups in the Py-amines influenced the pDNA binding affinity. In addition, it could be estimated that the numbers of Py-amines molecules interacting/binding with per DNA unit were ca. 22 (Py-1N), 11 (Py-2N), 4 (Py-3N) and 5 (Py-4N), which might influence the condensation/decondensation of pDNA.8 Moreover, the trends of pDNA binding affinity for the Py-amines amphiphiles analyzed by DNA gel-retardation assay were in accordance with the EB displacement results. In other words, these results indicated that the pDNA binding affinity could be tuned by selecting cationic moieties/headgroups with proper charge numbers, which should provide a possible approach for controlled gene loading/release for drug/gene dual delivery systems.
 |
| | Fig. 2 Ethidium bromide (EB) displacement assay of the plasmid DNA binding affinity of the synthesized Py-amines amphiphiles in 0.01 M PBS buffer. | |
 |
| | Fig. 3 Agarose-gel pDNA retardation assay for determining the pDNA binding affinity of the synthesized Py-amines cationic amphiphiles under various N/P charge ratios. | |
Particle size, surface potential and morphology of the Py-amines/pDNA complexes
The size and shape, surface potential, as well as the particle distribution of nano-scaled particles are already known to be essential factors for determining the related endocytosis pathway, intracellular trafficking and localization behaviors.33 Herein, dynamic laser scattering (DLS) method was utilized to characterize the average particle size and surface potential of the Py-amines/pDNA aggregates in distilled water with various N/P charge ratios. As depicted in Fig. 4a, it can be seen that the Py-amines and pDNA formed nano-scaled aggregates (diameter ca. 176–296 nm) at a low N/P charge ratio of 5, while when the N/P charge ratio was increased to 10–15, the particle sizes of the Py-amines/pDNA aggregates decreased to ca. 135–226 nm, indicating that the Py-amines amphiphiles possessed pDNA condensation abilities34 in water. However, with the continuous increase in the N/P charge ratio from 15 to 30, the average particle sizes of Py-amines/pDNA aggregates were drastically raised (about 81–196%). Notably, the pDNA complexes formed by Py-3N and Py-4N showed much larger particle size than that of the Py-1N and Py-2N counterparts at a higher N/P ratio (see Fig. 4a and ESI, S2†), which might be attributed to their higher hydration capabilities (see ESI, S3†). Moreover, for the Py-amines/pDNA complexes, the surface zeta potential converted from negative to positive with increase in the N/P charge ratio from 0 to 5 (see Fig. 4b), and then gradually increased with increase in the N/P ratio from 10 to 30. In addition, it could be found that the surface potentials were in a trend of Py-1N/pDNA (from +20.4 to +27.0 mV) > Py-2N/pDNA ≈ Py-3N/pDNA (from +7.9 to +20.4 mV) > Py-4N (from +5.5 to +11.8 mV). The results might be attributed to the comparatively high amino group numbers on Py-3N and Py-4N, which increased the hydrophilicity of the amphiphiles and further resulted in decrease in the surface potential and stability of the pDNA payloads.35
 |
| | Fig. 4 The particle sizes (a) and surface potentials (b) of the Py-amines/pDNA complexes under various N/P charge ratios in pure water, determined using a dynamic light scattering instrument (DLS, Malvern Nano ZS290, UK) with a laser wavelength of λ = 633 nm and a scattering angle of 90°. | |
Furthermore, we studied the morphology of the Py-amines/pDNA aggregates (N/P = 20) by means of atomic force microscopy (AFM). As shown in Fig. 5, it could be noticed that the Py-amines/pDNA complexes were spherical-shaped nanoparticles with an average particle size of 90–195 nm, indicating that the Py-amines amphiphiles could bind with pDNA via electrostatic interactions and form nano-scaled complexes in water. Moreover, the particle size observed by AFM was found to be smaller than that measured by DLS (around 130–490 nm). This could be explained as the Py-amines/pDNA aggregates measured by DLS were in their hydrated form, which made them observable as bigger-sized nanoparticles; moreover, the smaller-sized nanoparticles observed by AFM were due to the shrinkage of the Py-amines/pDNA complexes during the drying process in the AFM samples preparation.36
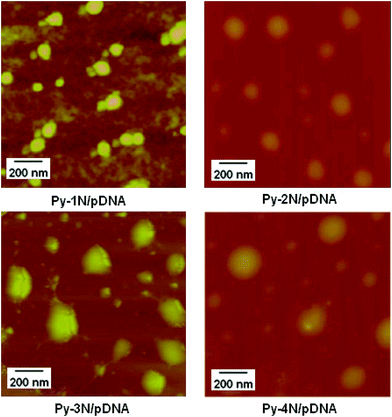 |
| | Fig. 5 AFM image of the morphology of Py-amines/pDNA complexes determined under an N/P ratio of 20 (inset: size distribution of Py-amines/pDNA measured by DLS at the same N/P charge ratio). | |
Proton buffering capacity of the Py-amines
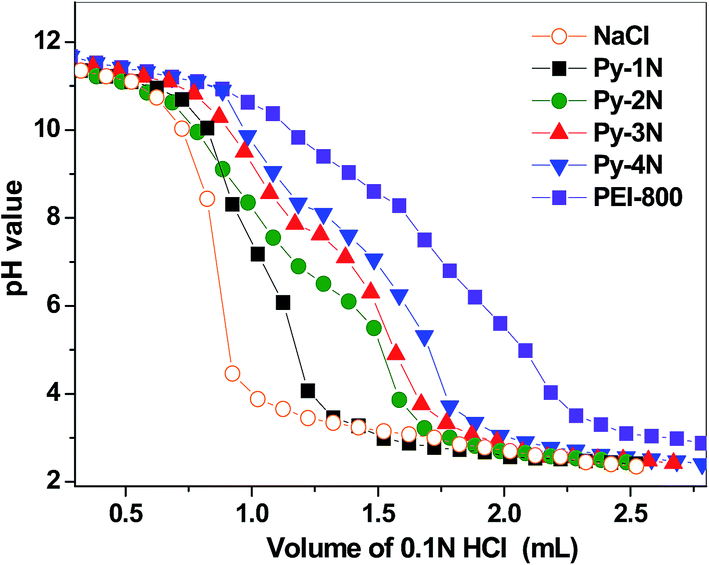
It had been revealed that the proton buffering capacity of multiamine-containing cationic gene carriers is regarded as an important factor because this influences the lysosome localization/release behaviors of their gene payloads through a “proton sponge” effect.37 Herein, the proton buffering capabilities of the Py-amines amphiphiles were examined by acid–base titration with NaCl (0.1 M, negative control) and PEI-800 (positive control) as the references, and the titration curves were obtained by the plot of pH values against the added HCl volume.38 As shown in Fig. 6, it could be seen that the negative control NaCl solution did not show any buffering effect. Notably, Py-2N, Py-3N and Py-4N showed higher buffering capacities than that of Py-1N, which might be attributed to their attached primary and secondary (1°, 2°) amine groups, which lead to higher protonation abilities in solution. Moreover, the positive control PEI-800 showed the highest buffering effect among the cationic amphiphiles, due to its three different types of amino groups (1°, 2° and 3°). It could be expected that the distinct proton buffering capacities of the Py-amines amphiphiles would lead to different ‘Lysosome escape’ effects, which might further cause different inter-lysosomal pDNA dissociation/releasing behaviors.
 |
| | Fig. 6 Acid–base titration curves of the Py-amines amphiphiles for determining the proton buffering capacities, with NaCl (0.1 M) as the negative control and PEI-800 as the positive control. | |
Cell proliferation inhibition of the Py-amines and Py-amines/pDNA by MTT assay
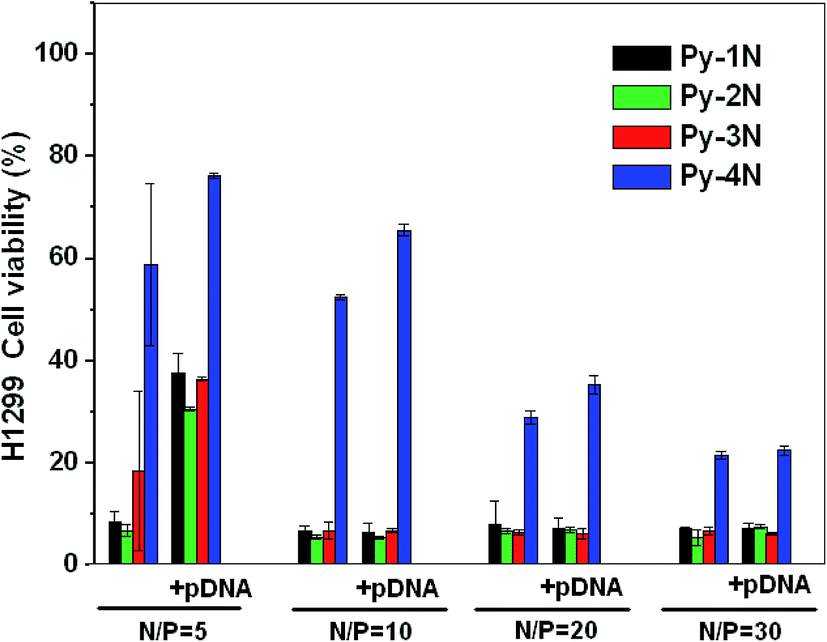
The cancer cell proliferation inhibition of Py-amines and Py-amines/pDNA complexes was examined by MTT assay in an H1299 (human lung cancer) cell line. As shown in Fig. 7, the cell viabilities of Py-1N (8.8%) and Py-2N (7.3%) were lower than that of Py-3N (18.6%) at the dose of N/P = 5 (pDNA 1 μg per well), whereas a much higher cell viability was observed with Py-4N at around 58%. After loading with pDNA (1 μg per well) at N/P = 5, it could be found that the cell viability greatly increased (Py-1N 38.5%, Py-2N 30.4%, Py-3N 36.8% and Py-4N 78.8%), possibly due to the binding of positively charged Py-amines with negatively charged pDNA diminishing their cell membrane permeation effect. Moreover, with the incubation of a higher dosage of Py-1N, Py-2N, Py-3N and their pDNA complexes (N/P > 10), very low H1299 viabilities (6.5–8.8%) could be observed, which indicated their H1299 tumor cell inhibition properties could not be interrupted by the loading of pDNA. However, the H1299 cells incubated with Py-4N (23.2–46.5%) and Py-4N/pDNA (26.2–67.5%) showed higher cell viabilities, indicating their lower tumor suppression efficiency. As far as the antitumor mechanism was concerned, prior studies revealed that some exogenous polyamines compounds could block the overexpressed polyamine (such as spermine and spermidine) transporter19 in the tumor cells. Thus, we supposed that the synthesized Py-amines might go through a similar pathway to block the polyamine transporter and then result in the inhibition of cancer cell proliferation and division. The results indicated that the cytotoxicity depended largely on the molecular structures of the Py-amines and that the Py-amines/pDNA complexes could be further optimized by changing the number of attached amino groups. Moreover, the Py-amines/pDNA complexes could be expected to serve as antitumor drug/pDNA dual delivery systems in practical applications.
 |
| | Fig. 7 The cytotoxicity of H1299 cells incubated with Py-amines/pDNA (+pDNA) complexes at various charge ratios of N/P (pDNA 1 μg per well), and with the Py-amines (−pDNA) at the same dosage as the control. | |
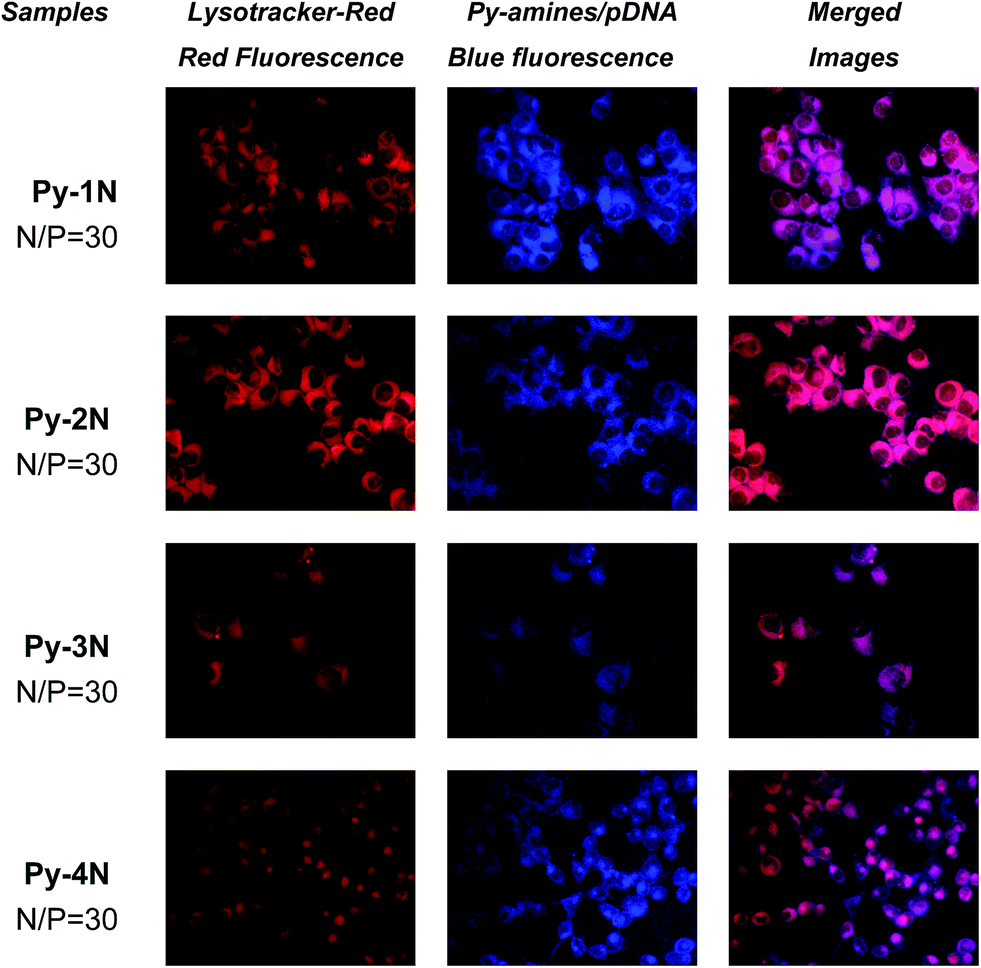
Lysosomal colocalization of the Py-amines/pDNA observed by fluorescent microscopy
The “Lysosomal escape” effect of gene carriers is regarded as a key factor for gene transfection, and the “endocytosis–endosomal/lysosomal escape-nuclear gene expression” pathway39 is widely regarded as the dominant intracellular trafficking route of cationic gene carriers because this facilitates the release of gene payloads trapped in lysosomes/endosomes via endosome swelling and breaking by buffering H+ and the subsequent accumulation of Cl−.40 In order to investigate the intracellular localization of Py-amines/pDNA complexes, the lysosome organelles of H1299 cells were stained with Lysotracker-red, a fluorescent agent for lysosomal-specific labeling. As shown in Fig. 8, after 4 h incubation of the Py-amines/pDNA complexes, an evident blue fluorescence emission located in the cytoplasm could be seen, and only weak blue fluorescence could be found around the plasma membrane. The blue fluorescence was attributed to the emission of pyrene fluorophore.41–43 Our previous study on pyrene-based cationic lipids also demonstrated that the strong blue-fluorescence emission of pyrene fluorophore could be observed even under high ionic strength conditions (in PBS buffer or cell culture medium).25 The intracellular blue fluorescence emission indicated that a large amount of the Py-amines/pDNA complexes could be efficiently taken up into the cytoplasm. Notably, a significant colocalization of red (Lysotracker-red) and blue (Py-amines/pDNA complexes) fluorescence were observed in the H1299 (Fig. 8, merged images) and COS-7 cells (ESI, S4†), suggesting that the Py-amines/pDNA complexes tended to localize inside the lysosome/endosome, which might due to the “proton sponge” effect of their attached amino groups.44 Moreover, it could be seen that the fluorescence of Py-3N/pDNA and Py-4N/pDNA partially overlapped with Lysotracker-red, indicating that they might possess a stronger “lysosomal escape” effect, due to their multiple (1° and 2°) amino groups for efficient H+ buffering, which was in accordance with that measured by acid–base titration experiments. The results illustrated that controllable ‘lysosome escape’ effects might be achieved by the incorporation of proper amine-containing moieties onto the pyrene fluorophore/hydrophobe.
 |
| | Fig. 8 Fluorescence images of Py-amines/pDNA (blue fluorescence) complexes in H1299 cell lines under an N/P ratio of 30, Lysotracker-red was used as the lysosomal specific labeling agent (red fluorescence). | |
On the other hand, as far as the nanoparticles were concerned, it was revealed that the surface features, such as charge, chemical component, and morphology, strongly affect the endocytosis pathway.40 Biju et al. recently revealed that clathrin-dependent endocytosis45 served as the dominant uptake pathway of some cationic insect neuropeptide (AST1) modified fluorescent quantum dots (QD).46 The endocytosis of some nanoparticles had been studied before; however, the endocytosis pathway of the most of nanoparticles (especially fluorescent nanoparticles), which act as gene/drug payloads still remained obscure. Thus, further investigation on the endocytosis/uptake pathway of the cationic Py-amines/pDNA complexes or other organic/polymeric fluorescent aggregates was necessary, and the related studies were thus carried out in our lab.
Conclusions
In summary, we designed and synthesized a series of pyrene-amine cationic amphiphiles (Py-amines) by the convenient chemical coupling of pyrene fluorophore/hydrophobe with various amine/polyamine derived cationic building blocks. The newly synthesized cationic Py-amines amphiphiles were able to bind with plasmid DNA and form self-assembled Py-amines/pDNA aggregates with nano-scaled particle size and positive surface potential, which greatly depends on the number of attached amino groups. In addition, the Py-amines amphiphiles showed different proton buffering capacity, which also relied on the structure of the attached amine/polyamine moieties. Moreover, the MTT results indicated the Py-amines and Py-amines/pDNA have structural-dependent anti-tumor activities in H1299 cells. Notably, the Py-amines/pDNA complexes possessed evident in vitro lysosome colocalization effects, which might be attributed to the proton buffering effects of the attached amine/polyamine moieties. In addition, the results might provide a new visible approach to design new anti-tumor drug/plasmid DNA dual delivery systems using synthesized amine/polyamine amphiphiles, and it deepens the understanding of the intracellular fate of amine/polyamine cationic amphiphiles and their gene payloads.
Acknowledgements
The authors are indebted to the financial supports partially from the national science foundation of China (21174160, 21002116 and 21372251).
Notes and references
- V. Biju, Chem. Soc. Rev., 2014, 43, 744–764 RSC.
- C. Fortier, Y. Durocher and G. De Crescenzo, Nanomedicine, 2014, 9, 135–151 CrossRef CAS PubMed.
- C. Wong-Baeza, I. Bustos, M. Serna, A. Tescucano, V. Alcantara-Farfan, M. Ibanez, C. Montanez, C. Wong and I. Baeza, Biochem. Biophys. Res. Commun., 2010, 396, 549–554 CrossRef CAS PubMed.
- T. Dewa, T. Asai, Y. Tsunoda, K. Kato, D. Baba, M. Uchida, A. Sumino, K. Niwata, T. Umemoto, K. Iida, N. Oku and M. Nango, Bioconjugate Chem., 2010, 21, 844–852 CrossRef CAS PubMed.
- P. Singh, C. Samori, F. M. Toma, C. Bussy, A. Nunes, K. T. Al-Jamal, C. Menard-Moyon, M. Prato, K. Kostarelos and A. Bianco, J. Mater. Chem., 2011, 21, 4850–4860 RSC.
- A. Bajaj, P. Kondaiah and S. Bhattacharya, Bioconjugate Chem., 2008, 19, 1640–1651 CrossRef CAS PubMed.
- A. Bajaj, P. Kondaiah and S. Bhattacharya, Bioconjugate Chem., 2007, 18, 1537–1546 CrossRef CAS PubMed.
- S. K. Misra, S. Naz, P. Kondaiah and S. Bhattacharya, Biomaterials, 2014, 35, 1334–1346 CrossRef CAS PubMed.
- A. Bajaj, P. Kondaiah and S. Bhattacharya, Biomacromolecules, 2008, 9, 991–999 CrossRef CAS PubMed.
- M. Khan, C. Y. Ang, N. Wiradharma, L. K. Yong, S. Liu, L. Liu, S. Gao and Y. Y. Yang, Biomaterials, 2012, 33, 4673–4680 CrossRef CAS PubMed.
- W. J. Yi, Q. F. Zhang, J. Zhang, Q. Liu, L. Ren, Q. M. Chen, L. Guo and X. Q. Yu, Acta Biomater., 2014, 10, 1412–1422 CrossRef CAS PubMed.
- S. M. Alex, M. R. Rekha and C. P. Sharma, Int. J. Pharm., 2011, 410, 125–137 CrossRef CAS PubMed.
- S. K. Cho and Y. J. Kwon, J. Controlled Release, 2011, 150, 287–297 CrossRef CAS PubMed.
- L. Djouhri-Bouktab, J. M. Rolain and J. M. Brunel, Anti-Infect. Agents, 2014, 12, 95–103 CrossRef CAS.
- L. Goyal, J. G. Supko, J. Berlin, L. S. Blaszkowsky, A. Carpenter, D. M. Heuman, S. L. Hilderbrand, K. E. Stuart, S. Cotler, N. N. Senzer, E. Chan, C. L. Berg, J. W. Clark, A. F. Hezel, D. P. Ryan and A. X. Zhu, Cancer Chemother. Pharmacol., 2013, 72, 1305–1314 CrossRef CAS PubMed.
- C. Wang, J. G. Delcros, J. Biggerstaff and O. Phanstiel, J. Med. Chem., 2003, 46, 2672–2682 CrossRef CAS PubMed.
- N. Kaur, J. G. Delcros, J. Archer, N. Z. Weagraff, B. Martin and O. Phanstiel, J. Med. Chem., 2008, 51, 2551–2560 CrossRef CAS PubMed.
- N. Kaur, J. G. Delcros, J. Imran, A. Khaled, M. Chehtane, N. Tschammer, B. Martin and O. Phanstiel, J. Med. Chem., 2008, 51, 1393–1401 CrossRef CAS PubMed.
- R. Seliga, M. Pilatova, M. Sarissky, V. Viglasky, M. Walko and J. Mojzis, Mol. Biol. Rep., 2013, 40, 4129–4137 CrossRef CAS PubMed.
- J. Wang, R. Gao, Q. Li, S. Xie, J. Zhao and C. Wang, Chem. Biol. Drug Des., 2012, 80, 909–917 CAS.
- Y. Dong, Y. Zhu, J. Li, Q.-H. Zhou, C. Wu and D. Oupický, Mol. Pharm., 2012, 9, 1654–1664 CrossRef CAS PubMed.
- M. Zhu, G. Nie, H. Meng, T. Xia, A. Nel and Y. Zhao, Acc. Chem. Res., 2012, 46, 622–631 CrossRef PubMed.
- Y. Li, Y. Zhu, K. Xia, R. Sheng, L. Jia, X. Hou, Y. Xu and A. Cao, Biomacromolecules, 2009, 10, 2284–2293 CrossRef CAS PubMed.
- S. Bhattacharya and S. S. Mandal, Biochim. Biophys. Acta, Biomembr., 1997, 1323, 29–44 CrossRef CAS.
- R. Sheng, T. Luo, Y. Zhu, H. Li and A. Cao, Macromol. Biosci., 2010, 10, 974–982 CrossRef CAS PubMed.
- M. Muñoz-Úbeda, S. K. Misra, A. L. Barrán-Berdón, S. Datta, C. Aicart-Ramos, P. Castro-Hartmann, P. Kondaiah, E. Junquera, S. Bhattacharya and E. Aicart, Biomacromolecules, 2012, 13, 3926–3937 Search PubMed.
- R. Sheng, T. Luo, H. Li, J. Sun, Z. Wang and A. Cao, Bioorg. Med. Chem., 2013, 21, 6366–6377 CrossRef CAS PubMed.
- R. Sheng, T. Luo, Y. Zhu, H. Li, J. Sun, S. Chen, W. Sun and A. Cao, Biomaterials, 2011, 32, 3507–3519 CrossRef CAS PubMed.
- T. I. Kim, T. Rothmund, T. Kissel and S. W. Kim, J. Controlled Release, 2011, 152, 110–119 CrossRef CAS PubMed.
- S. K. Misra, M. Muñoz-Úbeda, S. Datta, A. L. Barrán-Berdón, C. Aicart-Ramos, P. Castro-Hartmann, P. Kondaiah, E. Junquera, S. Bhattacharya and E. Aicart, Biomacromolecules, 2013, 14, 3951–3963 CrossRef CAS PubMed.
- D. M. Patterson, L. A. Nazarova and J. A. Prescher, ACS Chem. Biol., 2014, 9, 592–605 CrossRef CAS PubMed.
- R. Sheng, T. Luo, H. Li, J. Sun, Z. Wang and A. Cao, Colloids Surf., B, 2014, 116, 32–40 CrossRef CAS PubMed.
- Z. U. Rehman, I. S. Zuhorn and D. Hoekstra, J. Controlled Release, 2013, 166, 46–56 CrossRef PubMed.
- Q. F. Zhang, W. J. Yi, B. Wang, J. Zhang, L. Ren, Q. M. Chen, L. Guo and X. Q. Yu, Biomaterials, 2013, 34, 5391–5401 CrossRef CAS PubMed.
- B. Qin, Z. Chen, W. Jin and K. Cheng, J. Controlled Release, 2013, 172, 159–168 CrossRef CAS PubMed.
- R. Sheng, K. Xia, J. Chen, Y. Xu and A. Cao, J. Biomater. Sci., Polym. Ed., 2013, 24, 1935–1951 CrossRef CAS PubMed.
- I. Richard, M. Thibault, G. De Crescenzo, M. D. Buschmann and M. Lavertu, Biomacromolecules, 2013, 14, 1732–1740 CrossRef CAS PubMed.
- J. Shi, J. G. Schellinger, R. N. Johnson, J. L. Choi, B. Chou, E. L. Anghel and S. H. Pun, Biomacromolecules, 2013, 14, 1961–1970 CrossRef CAS PubMed.
- R. Duncan and S. C. W. Richardson, Mol. Pharm., 2012, 9, 2380–2402 CrossRef CAS PubMed.
- V. Biju, T. Itoh and M. Ishikawa, Chem. Soc. Rev., 2010, 39, 3031–3056 RSC.
- Y. Shiraishi, K. Ishizumi, G. Nishimura and T. Hirai, J. Phys. Chem. B, 2007, 111, 8812–8822 CrossRef CAS PubMed.
- Y. Shiraishi, Y. Tokitoh and T. Hirai, Org. Lett., 2006, 8, 3841–3844 CrossRef CAS PubMed.
- Y. Shiraishi, Y. Tokitoh, G. Nishimura and T. Hirai, J. Phys. Chem. B, 2007, 111, 5090–5100 CrossRef CAS PubMed.
- A. K. Varkouhi, M. Scholte, G. Storm and H. J. Haisma, J. Controlled Release, 2011, 151, 220–228 CrossRef CAS PubMed.
- A. Anas, T. Okuda, N. Kawashima, K. Nakayama, T. Itoh, M. Ishikawa and V. Biju, ACS Nano, 2009, 3, 2419–2429 CrossRef CAS PubMed.
- V. Biju, D. Muraleedharan, K. Nakayama, Y. Shinohara, T. Itoh, Y. Baba and M. Ishikawa, Langmuir, 2007, 23, 10254–10261 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/c4ra06879c |
|
| This journal is © The Royal Society of Chemistry 2015 |
Click here to see how this site uses Cookies. View our privacy policy here.