Proteins, cells, and tissues in patterned environments
Ulrich S.
Schwarz
a,
Celeste M.
Nelson
b and
Pascal
Silberzan
c
aHeidelberg University, Institute for Theoretical Physics and BioQuant, Philosophenweg 19, D-69120 Heidelberg, Germany. E-mail: Ulrich.Schwarz@bioquant.uni-heidelberg.de
bDepartment of Chemical and Biological Engineering and Department of Molecular Biology, Princeton University, Princeton, NJ 08544, USA. E-mail: celesten@princeton.edu
cLaboratoire Physico-Chimie Curie-UMR 168, Institut Curie, Centre de Recherche, CNRS, UPMC, F-75248 Paris, France. E-mail: pascal.silberzan@curie.fr
 Ulrich S. Schwarz | Ulrich S. Schwarz, Heidelberg University, Germany |
 Celeste M. Nelson | Celeste M. Nelson, Princeton University, USA |
 Pascal Silberzan | Pascal Silberzan, Institut Curie, France |
Introduction
During the last two decades, researchers have increasingly focused on the role of spatial coordination in biological molecules, cells and tissues. Using microscale tools from materials science, such as printing of adhesive patterns or fabrication of reaction chambers, it has been revealed that the amazing capacity of biological systems to organize themselves strongly depends on the way the environment is spatially structured.1 These insights not only unveil formerly unknown mechanisms underlying biological functions, but also permit a more quantitative understanding of biological systems, including e.g. the ability to control cell behaviour by designing appropriate environments.2The line of research discussed in this themed issue was sparked by the development of microcontact printing, a versatile technique to anchor organic molecules to well-defined regions on a planar substratum using microfabricated stamps inked with a solution of the molecule to be printed.3 This technique then paved the way to study how cells respond to spatial distributions of ligand molecules for cell adhesion. A standard choice has been to print fibronectin onto a culture dish in a desired spatial arrangement to activate the integrin adhesion receptors of the cell. In a pioneering study, it was found that cell survival requires a sufficient degree of spatial extension, without which the cells would undergo programmed cell death.4 Since this early work, a rapidly growing body of studies using micropatterned substrates for cell culture has demonstrated that many cellular functions depend on the spatial presentation of ligands, for example cell division5 and stem cell differentation.6 In addition to the spatial distribution of ligands, the mechanical properties of the environment have also emerged over the last decade as a critical determinant of cell behaviour. For example, it has been shown that stem cell differentiation depends crucially on substratum stiffness.7 These insights make it necessary to measure cellular forces, which again can be achieved using microfabrication tools. For example, microfabrication has been used to generate fiducial markers on flat elastic substrata8 and to build arrays of pillars that function as local force sensors.9 As illustrated by several contributions to this themed issue, micropatterning and microfabrication have become standard techniques to study a large range of interesting questions regarding how single cells interact with their environment. Due to the quantitative nature of such an approach, these studies naturally lend themselves to image processing and mathematical modelling.
Spatial control of biological processes is not restricted to the cellular level, but also occurs at the molecular and pluricellular scales. During tissue development and regeneration, two essential aspects are cell migration and cell growth. In a pioneering study using microcontact printing, it was shown that the spatial patterns of proliferating cells within a tissue strongly depend on the tissue's overall shape.10 Using microfabricated barriers to control the release of cell collectives from confinement and image processing to track their movement, quantitative analysis of collective cell migration became possible.11 For example, by combining this approach with soft elastic substrata for force measurements, it was recently shown that mechanical waves occur in expanding cell monolayers.12
On the molecular level, a line of research of increasing importance for our understanding of biological systems is the reconstitution of biological processes in biomimetic assays, in which a minimal number of molecular components recapitulate some essential aspect of the biological system. For example, it has recently been shown using microcontact printing of activators of actin polymerization that the contraction of the actomyosin cytoskeleton can be reconstituted on a flat substrate.13 Another pioneering study revealed that the spatiotemporal oscillations of the bacterial Min-proteins can be reconstituted with only two proteins interacting with lipid bilayers on a substrate.14 Later it was shown that these processes can be reconstituted in patterned environments, both for patches of membrane deposited on gold patterns on a glass substrate15 and in microfabricated chambers.16
In summary, the spatial control of molecules, cells and tissue is a rapidly developing, exciting and very promising field of research at the interface between materials science, biophysics and biology. In the following, we will discuss how the different contributions of this themed issue can be placed in this context. We structure our discussion by the different scales provided by molecules, cells and tissue.
Molecules
Because of the versatility and flexibility offered by photolithography, substrates can now be prepared with several different moieties grafted onto the surface. These molecules can be mixed on the same pattern or used as pure components in adjacent patterns. This has been achieved by a combination of local UV-photodestruction and excitation of photocleavable molecules by Biswas et al. (DOI: 10.1039/C3SM53000K). In the future, such a technology may allow the patterning of several proteic synergic components at well-defined positions to more easily dissect molecular mechanisms. This approach also demonstrates the substantial progress made in the past few years in the area of locally controlled surface treatments.In cells, by definition a confined environment, the precise spatio-temporal localization of proteins is crucial to understand their function. A prime example is the system of the bacterial Min-proteins interacting with lipid membranes, which gives rise to oscillations that strongly depend on the membrane geometry.15,16 The formation of these dynamic patterns has now been modelled by Hoffmann and Schwarz (DOI: 10.1039/C3SM52251B) for micropatterned environments using particle-based stochastic computer simulations. The authors showed that two fundamentally different oscillation patterns can emerge and stochastically switch stability if the micropattern is designed appropriately (Fig. 1 (left)).
Another example for the reconstitution of the confinement typical for cellular systems is the study of high-density actin filaments in microfabricated chambers conducted by Alvarado et al. (DOI: 10.1039/C3SM52421C). In this experimental model, the behavior of the microfilaments can be examined by carefully tuning various parameters such as the filament density, the presence of crosslinkers or the geometry of the confinement itself. A nematic order naturally emerges under these conditions at high density and, interestingly, the aspect ratio of the boxes is a control parameter of this nematic order. Therefore, wall-induced alignment competes with deformation in the bulk of these nematic phases. Such assays, both regarding the geometries and their mode of analysis, are also well adapted to investigate the interactions of other components of the cytoskeleton, such as molecular motors, with the microfilaments.
Cells
Several contributions examine how varying the geometry of adhesive micropatterns affects cell behaviour. Müller et al. (DOI: 10.1039/C3SM52424H) studied the cell shape and organization of the actin cytoskeleton on adhesive stripes of decreasing width. As is typical for such cell culture, they observed a system of thick actin bundles (stress fibers) using confocal fluorescence microscopy. In cells on wide strips, several stress fibers were found to run parallel to the direction of the stripe, while in cells on stripes of widths below a critical value of around 15 μm, the organization switched to two dominant stress fibers running along the edges of the stripe. A quantitative analysis revealed a window of bistability between 5 and 20 μm. Strikingly, a similar bistable response was observed in a mathematical model for cell shape based on a minimization of the surface energy. This suggests that the transition in stress fiber organization corresponds to a change in cell shape from a spread to a bulged morphology as the stripe width decreases.Micropatterns for single cell studies have not only become a standard tool in biophysics and cell biology, but they are also increasingly used in areas such as biotechnology and systems biology, which often require high throughput. In order to optimize the occupancy level of micropatterns for cell arrays, Röttgermann et al. (DOI: 10.1039/C3SM52419A) conducted a systematic study of how cells explore arrays of adhesive square islands. They found that if the cells are allowed to search the substratum for adhesive islands for 3–5 hours, they can reach occupancy levels of 40–60%. These results can be explained if the cells undergo a random walk search process. In many cases of practical interest, for example when dealing with rare cells like stem or tumour cells, this self-organization approach is an attractive alternative to the standard procedure of using a surplus of cells and then washing most of them away. This strategy also avoids harsh treatment that might damage cells or change their behaviour.
This themed issue contains two articles that use micropatterning techniques to investigate cellular systems with specific biological functions, namely platelets or neurons. Platelets are specialized blood cells that, upon activation after injury, contract the blood clot. Using both topographically and biochemically structured surfaces, Sandmann et al. (DOI: 10.1039/C3SM52636D) mimicked the essential ingredients of the heterogeneous situation in a wounded blood vessel. They show that platelet spreading is very dynamic and strongly adapts to the environmental cues, but that the cells always manage to achieve their maximal spreading area of around 30 μm2. This result is surprising because in their non-activated state, platelets have a diameter of only 2–5 μm. This study shows that micropatterning can be used to study highly relevant physiological and medical situations in quantitative detail.
Tomba et al. (DOI: 10.1039/C3SM52342J) used stripes with variable widths and lengths and with a central disk to study the kinetics by which neuronal extensions (neurites) become an axon. The authors found that neurites on stripes of smaller widths performed better because they had a larger extension velocity and thus were the first to reach the critical length of approximately 40 μm that seems to be required to become an axon. The authors developed a mathematical model for neurite extension that is able to fit all experimental data with the same parameter set and that now can be used for the design of e.g. artificial neuronal circuits.
Three contributions to this themed issue address the role of substratum stiffness and forces in cellular systems. In their theoretical study, Banerjee et al. (DOI: 10.1039/C3SM52647J) use a continuum model to predict cell shape and forces for adhesive patterns on an elastic foundation. The model combines contractility in the bulk of the cell (modeled as a negative pressure in the elastic equations) with peripheral contractility and bending rigidity. The model is solved with an innovative particle-based method and predicts transitions between convex and concave cell shapes that might be induced experimentally by changing the stiffness of the peripheral region.
In another theoretical study, Nisenholz et al. (DOI: 10.1039/C3SM52524D) examined how cell polarity depends on cell shape and the stiffness of the surrounding matrix using the formalism of polarizable force dipoles in an elastic continuum. Experimentally, this would correspond to changes in the magnitude and direction of traction forces due to a dynamic response of the actomyosin system to external cues. Their model predicts that different cell shapes would exert maximal force above and below a critical value in matrix stiffness.
The study by Moussus et al. (DOI: 10.1039/C3SM52318G) addresses a long-standing issue regarding cellular forces, namely if the internal stress state of a cell or a cluster of cells can be inferred from its traction pattern. Similar to the work of Banjeree et al., adherent cells are modeled as thin elastic films adhering to a planar substratum. Assuming equal displacements on both sides of the cell–substratum interface, the stress tensor inside the cell layer can be directly calculated from the local displacement field of the substratum without the need to explicitly reconstruct the traction forces. By confining the cells or clusters to increasingly smaller islands on the elastic substratum, the authors show that mechanical stress increases with confinement.
The environment also has a dramatic effect in the bacterial system of Pseudomonas aeruginosa investigated by Deforet et al. (DOI: 10.1039/C3SM53127A). Mutants of this bacterium, identified in a previous study17 as “hyperswarmers”, form colonies that expend very rapidly at the surface of gels. In the present study it is shown that these mutants actually do not individually swim faster than the wild type strain. However, the details of their displacements and in particular their ability to modulate their changes of direction both in frequency and in amplitude are very different. In the context of swarming, this then leads to a faster extension of the colony.
Tissue
As with molecules and cells, surface patterning and confinement are also of large relevance for tissues. In embryonic development in particular, cells are often constrained by their environment in the form of populations comprising a relatively small number of cells. One particularly important aspect of these systems are mechanical forces, as they arise naturally in confined systems and have been shown to regulate many essential cellular functions including growth, division and migration. Therefore it is important to measure traction forces for well-defined cell clusters, a goal which can be achieved by the patterning of soft elastomeric substrata. Most studies in this field use polyacrylamide substrata. Hampe et al. (DOI: 10.1039/C3SM53123F) have developed a novel lift-off technology to achieve micropatterning of silicone rubber substrata, which have a long shelf life and different mechanical and optical properties. They use their method to show that cardiac myocytes exert larger forces when working together on appropriately designed patterns (Fig. 1 (middle)).Patterning is also important to understand the organization of a large number of cells. When Duclos et al. (DOI: 10.1039/C3SM52323C) cultured elongated 3T3 fibroblasts in non-bounded conditions, they found that the cells formed juxtaposed clusters having a well-defined nematic order (similar to the analysis by Alvarado et al. for actin filaments, but on a very different scale). However, the presence of defects trapped by the progressive jamming of this cell monolayer18 kept them from attaining a perfect macroscopic order. This limitation is avoided when cells are grown in well-defined bands whose width is smaller than the orientation correlation length. The versatility of the patterning techniques allows making more complex structures, such as crosses where alignment in the center results from a competition between different arms (Fig. 1 (right)).
In vivo, tissue patterning is often three-dimensional and therefore a large effort is underway to extend two-dimensional studies into the third dimension. One standard model for tissues in three dimensions is the growth of spheroids in collagen gels. Rather than patterning the surrounding collagen gel, MacKay et al. (DOI: 10.1039/C3SM52265B) have introduced a novel approach to pattern the spheroid by internal cell activity. The authors genetically engineered promotors for the cytoskeletal regulator Rac, such that its activity can be tuned externally by adding small molecules. In this way, they are able to control the extent and timing of cell migration in the spheroid. When dealing with two populations under the control of two distinct small molecule inducers, the 3D architecture of these mixed populations can be controlled by their successive additions and withdraws, thereby allowing the spatial organization of multiple cell populations in three dimensions.
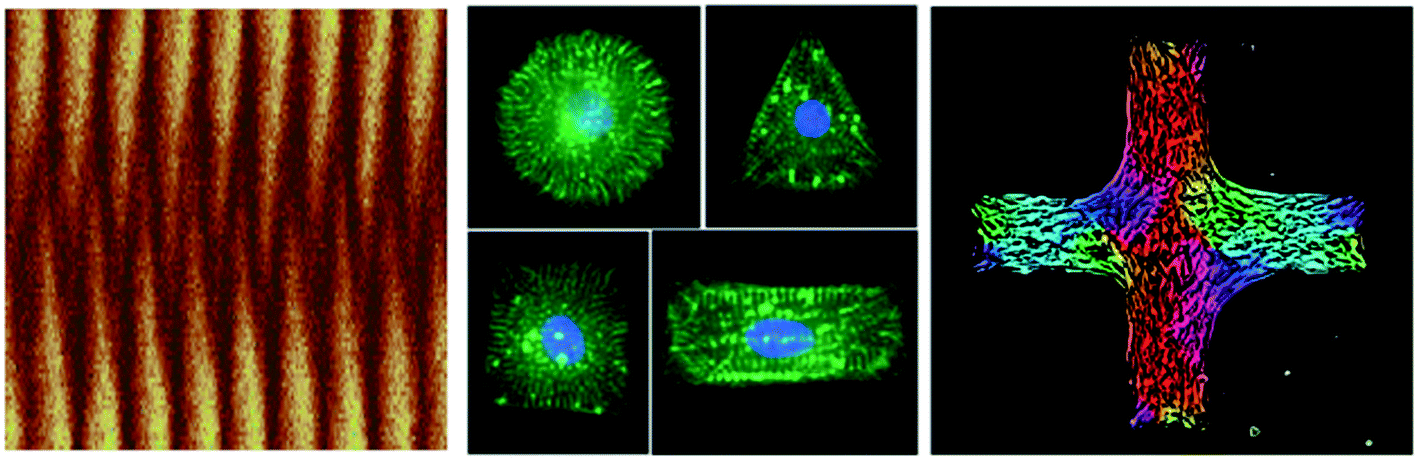 | ||
| Fig. 1 Images reproduced by permission of The Royal Society of Chemistry (RSC). Left: kymograph from a computer simulation of the pole-to-pole oscillations of the protein MinD in a confined geometry from Hoffmann and Schwarz (DOI: 10.1039/C3SM52251B). Middle: cardiomyocytes on differently shaped adhesive islands on an elastic substratum from Hampe et al. (DOI: 10.1039/C3SM53123F). Right: domains of different orientations for a layer of fibroblast cells on a cross-shaped adhesive island from Duclos et al. (DOI: 10.1039/C3SM52323C). | ||
Acknowledgements
USS and PS thank the Kavli Institute for Theoretical Physics (KITP) at Santa Barbara for its hospitality during the Workshop on Active Matter. This research was supported in part by the National Science Foundation under Grant No. NSF PHY11-25915 for the workshop program at KITP. USS is a member of the cluster of excellence CellNetworks at Heidelberg and acknowledges support by the EU-project MEHTRICS. CMN holds a Career Award at the Scientific Interface from the Burroughs Wellcome Fund. PS is a member of the Labex CelTisPhyBio.References
- T. Vignaud, L. Blanchoin and M. Théry, Directed cytoskeleton self-organization, Trends Cell Biol., 2012, 22, 671–682 CrossRef CAS PubMed.
- B. Geiger, J. P. Spatz and A. D. Bershadsky, Environmental sensing through focal adhesions, Nat. Rev. Mol. Cell Biol., 2009, 10, 21–33 CrossRef CAS PubMed.
- K. L. Prime and G. M. Whitesides, Self-assembled organic monolayers: model systems for studying adsorption of proteins at surfaces, Science, 1991, 252, 1164–1167 CrossRef CAS.
- C. S. Chen, M. Mrksich, S. Huang, G. M. Whitesides and D. E. Ingber, Geometric Control of Cell Life and Death, Science, 1997, 276, 1425–1428 CrossRef CAS.
- J. Fink, et al. External forces control mitotic spindle positioning, Nat. Cell Biol., 2011, 13, 771–778 CrossRef CAS PubMed.
- K. Kilian, B. Bugarija, B. Lahn and M. Mrksich, Geometric cues for directing the differentiation of mesenchymal stem cells, Proc. Natl. Acad. Sci. U. S. A., 2010, 107, 4872–4877 CrossRef CAS PubMed.
- A. Engler, S. Sen, H. Sweeney and D. Discher, Matrix Elasticity Directs Stem Cell Lineage Specification, Cell, 2006, 126, 677–689 CrossRef CAS PubMed.
- N. Q. Balaban, et al. Force and focal adhesion assembly: a close relationship studied using elastic micropatterned substrates, Nat. Cell Biol., 2001, 3, 466–472 CrossRef CAS PubMed.
- J. L. Tan, et al. Cells lying on a bed of microneedles: an approach to isolate mechanical force, Proc. Natl. Acad. Sci. U. S. A., 2003, 100, 1484–1489 CrossRef CAS PubMed.
- C. Nelson, et al. Emergent patterns of growth controlled by multicellular form and mechanics, Proc. Natl. Acad. Sci. U. S. A., 2005, 102, 11594–11599 CrossRef CAS PubMed.
- M. Poujade, et al. Collective migration of an epithelial monolayer in response to a model wound, Proc. Natl. Acad. Sci. U. S. A., 2007, 104, 15988–15993 CrossRef CAS PubMed.
- X. Serra-Picamal, et al. Mechanical waves during tissue expansion, Nat. Phys., 2012, 8, 628–634 CrossRef CAS.
- A.-C. Reymann, et al. Actin Network Architecture Can Determine Myosin Motor Activity, Science, 2012, 336, 1310–1314 CrossRef CAS PubMed.
- M. Loose, E. Fischer-Friedrich, J. Ries, K. Kruse and P. Schwille, Spatial Regulators for Bacterial Cell Division Self-Organize into Surface Waves In Vitro, Science, 2008, 320, 789–792 CrossRef CAS PubMed.
- J. Schweizer, et al. Geometry sensing by self-organized protein patterns, Proc. Natl. Acad. Sci. U. S. A., 2012, 109, 15283–15288 CrossRef CAS PubMed.
- K. Zieske and P. Schwille, Reconstitution of Pole-to-Pole Oscillations of Min Proteins in Microengineered Polydimethylsiloxane Compartments, Angew. Chem., Int. Ed., 2013, 52, 459–462 CrossRef CAS PubMed.
- D. Van Ditmarsch, et al. Convergent Evolution of Hyperswarming Leads to Impaired Biofilm Formation in Pathogenic Bacteria, Cell Rep., 2013, 4, 697–708 CrossRef CAS PubMed.
- T. E. Angelini, et al. Glass-like dynamics of collective cell migration, Proc. Natl. Acad. Sci. U. S. A., 2011, 108, 4714–4719 CrossRef CAS PubMed.
| This journal is © The Royal Society of Chemistry 2014 |