
Synthesis of ethyl (R)-4-cyano-3-hydroxybutyrate in high concentration using a novel halohydrin dehalogenase HHDH-PL from Parvibaculum lavamentivorans DS-1†
Abstract
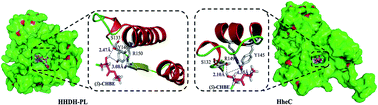
We identified and characterized a novel halohydrin dehalogenase HHDH-PL from Parvibaculum lavamentivorans DS-1. Study of substrate specificity indicated that HHDH-PL possessed a high activity toward ethyl (S)-4-chloro-3-hydroxybutanoate ((S)-CHBE). After optimizations of the pH and temperature, whole cell catalysis of HHDH-PL was applied to the synthesis of ethyl (R)-4-cyano-3-hydroxybutyrate (HN) at 200 g L−1 of (S)-CHBE, which gave 95% conversion and 85% yield in 14 h.
 Please wait while we load your content...
Please wait while we load your content...

