
Increased disulfide peptide sequence coverage via “cleavage ON/OFF” switch during nanoelectrospray†
Gongyu Li,
Yue Yin and
Guangming Huang*
Department of Chemistry, University of Science and Technology of China, Hefei, Anhui 230026, P.R. China. E-mail: gmhuang@ustc.edu.cn; Fax: +86 551 63600706; Tel: +86 551 63600706
First published on 4th November 2014
Abstract
Herein, we developed an in situ disulfide cleavage protocol. It is based on the selective reaction between disulfide and the hydroxyl radical, which could be rapidly and controllably generated via induced nanoelectrospray. In less than one minute, increased disulfide peptide sequence coverage could be obtained via switching the “cleavage ON/OFF” modes between conventional and induced nanoelectrospray.
Identification of posttranslational modifications1–4 is of increasing importance in functional proteomics studies, since posttranslational modifications reversibly or irreversibly alter the nature of proteins by chemical modification. Disulfide bond (DSB) formation, one of the most well known posttranslational modifications, is crucial for stabilizing tertiary structures of proteins5–9 and modulating activity of enzymes.10 As a promising tool for DSB-linked protein identification, mass spectrometry (MS) benefits widely from the striking capability of mass- and structure-accessibility. However, DSB would “bury” backbone sequence information in typical collision induced dissociation-mass spectrometry (CID-MS) since CID does not produce informative fragments if DSB remains intact. Thus, cleavage of DSB is normally mandatory prior to CID-MS identification since it would release “buried” peptide backbone.11 In situ DSB cleavage method should enable MS to obtain the actual implementation in the realm of top-down2,12–14 or bottom-up15–17 proteomics, taking de novo sequencing as an example.
The majority of classic DSB cleavage methods include chemical reduction using Tris(2-carboxyethyl) phosphine18–20 or dithiothreitol (DTT)21 and alternatively electrochemical reduction.22–25 To avoid time- and sample-consuming wet chemistry, MS-based DSB cleavages have been increasingly reported, such as electron capture dissociation (ECD),26,27 electron transfer dissociation (ETD),28,29 in-source decay,30 reduction with reducing matrix in MALDI31 and thermal dissociation.32 Previously, ultraviolet (UV) light photodissociation of DSB in gaseous proteins was reported,33 in which efficient DSB cleavage was performed as well as some backbone cleavages, and instrumentation modification is necessary to guild UV light into ion trap mass spectrometer. Interestingly, oxidative cleavage based on selective oxidation between hydroxyl radical (HO˙) and DSB was reported by Xia et al. for selective DSB cleavage free of backbone damage.34,35 The cleavage products, radical additive ions,34 reveal backbone sequence using low temperature plasma as additional HO˙ source during conventional nanoelectrospray ionization (nanoESI). Recently, reactive oxygen species (such as HO˙) was also reported to be produced during conventional ESI via corona discharge.36 Herein, we aimed at using induced nanoelectrospray ionization (InESI)37,38 source to provide HO˙ for instant and selective cleavage of DSB in peptides/proteins without extra plasma equipment.
In present work, this goal has been accomplished via in situ HO˙ generation by enhanced corona discharge39–45 during the InESI process (Fig. 1a). HO˙ could be controllably generated through regulating the amplitude and frequency of pulsed high voltage.
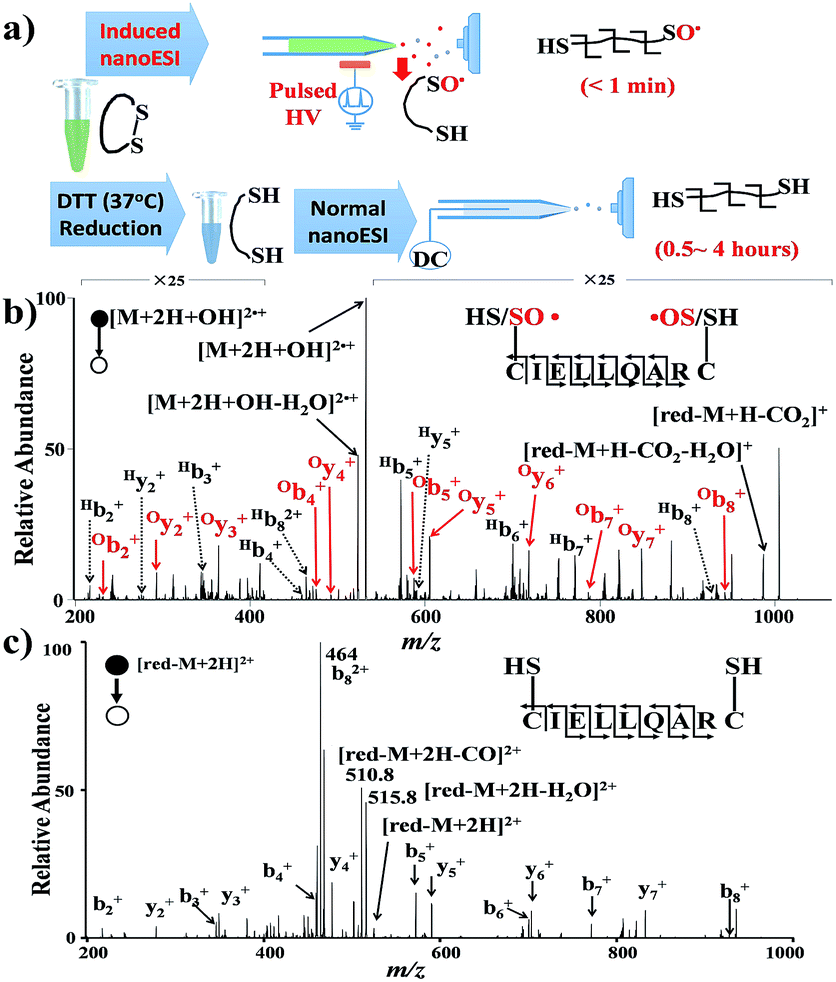 | ||
| Fig. 1 Experimental scheme and CID spectra of selectin binding peptide. (a) Comparison of InESI-MS (above) and classic DTT reduction nanoESI-MS (below) in total time required. CID spectra of selectin binding peptide via (b) radical additive ion derived from InESI (pulse, 1 kHz, 6 kV) and (c) protonated ion from DTT reduction nanoESI (DC, 1.5 kV). | ||
Selectin binding peptide (SCIELLQARCS), whose sequence is fully protected by DSB, was tested by InESI-CID-MS protocol. Water solution, containing 10 mM ammonium acetate (NH4OAc), was used as the spray solvent, and it required less than one minute for whole InESI-CID-MS experiment. Upon turning on the pulsed voltage, sulfinyl radicals were observed under typical operation condition (HSCIELLQARCSO˙/˙OSCIELLQARCSH) (the above path in Fig. 1a), as evidenced by the MS spectrum (Fig. S1†) and corresponding tandem MS spectrum of the radical ion ([M + 2H + OH]2˙+) (Fig. 1b). Sequence coverage of selectin binding peptide from the present method was similar to that from conventional nanoESI-MS method (Fig. 1c) following chemical reduction via DTT (the below path in Fig. 1a), which would be tedious (cost 0.5–4 hours) and inconvenient.22 Homolytic cleavage of the DSB was identified with the MS3 spectra of the oxygen additive fragment ions (Oy2+, Oy3+, Oy4+, Oy5+, Oy6+ and Oy7+). As shown in Fig. S2,† the 62 Da neutral loss, which is attributed to the loss of CH2SO from the oxidized cysteine residue, was observed as the major fragmentation pathway. Apart from selectin binding peptide, another peptide with an intra-disulfide chain (somatostatin, AGSCKNFFWKTFTSCS) (Table S1†) was observed to undergo similar DSB cleavage pathway and increased sequence coverage was also achieved with the present method.
Furthermore, we found that InESI-CID-MS protocol could rapidly discover backbone sequence of peptide both beyond and within DSB with slight modification of conventional nanoESI, as demonstrated by the “cleavage ON/OFF” experiment (Fig. 2a). Firstly, in “cleavage OFF” mode (Fig. 2a “OFF”), conventional nanoESI-CID-MS analysis of GSSG was conducted and peptide sequences containing intact DSB were obtained (Fig. 2b). Then, in “cleavage ON” mode (Fig. 2a “ON”), GSSG was reacted with HO˙ and peptide sequences from DSB dissociation was obtained (Fig. 2c). The “cleavage ON” and “cleavage OFF” modes could be swiftly and easily switched (Fig. 2a). Comparison of their tandem MS spectra enables identification of either intact or DSB-cleaved peptide, with the same solution and spray emitter. The above results implied this protocol would be an informative tool for DSB protein analysis, such as exploring the number and location of DSB as well as concomitant backbone sequence information within DSB.
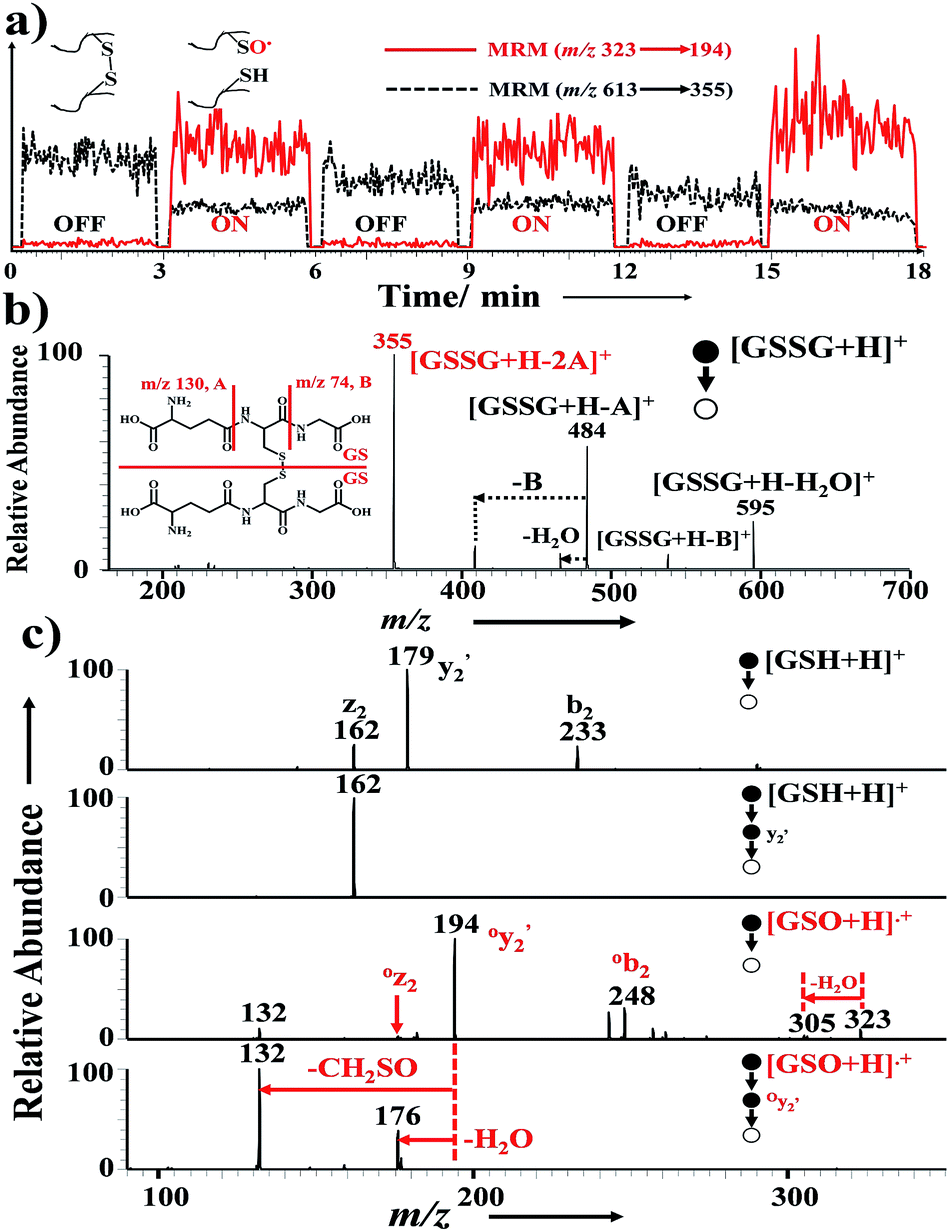 | ||
| Fig. 2 “Cleavage ON/OFF” experiment with (a) switch between InESI and nanoESI for GSSG analysis. (b) GSSG (m/z 613, CID m/z 355) and (c) GSO˙ (m/z 323, CID m/z 194) were monitored in multiple reaction monitor mode. The “cleavage ON” mode was operated as follows, pulsed high voltage (1 kHz, 7 kV) was applied on the ring electrode outside the tip for InESI operation and DC high voltage for conventional nanoESI was turned off. For “cleavage OFF” mode, DC high voltage (1 kV) was applied on the electrode inside the tip for nanoESI operation and pulsed high voltage for InESI was turned off. NH4OAc (10 mM) was added into the spray solution for all experiments. | ||
DSB cleavage in InESI-CID-MS is attributed to the selective reaction between DSB and HO˙, since tandem MS spectra of the product ions showed almost identical pathways to previous study.34 We also assume that the in situ generation of HO˙ correlates with the enhanced corona discharge originated from the regulation of pulsed high voltage in InESI. To validate these assumptions, our investigation was performed in three steps: (1) exploring the relationship between DSB cleavage and pulsed high voltage, (2) characterizing the relationship between pulsed high voltage and corona discharge and (3) proving the relationship between DSB cleavage and corona discharge.
In order to explore the relationship between DSB cleavage and pulsed high voltage, GSSG (including an interchain DSB) was chosen as a model compound. Various amplitudes of pulsed high voltage were used to analyze 8 μM GSSG in water solution. With increased amplitude of the pulsed high voltage, increased intensities of the cleavage products (GSO˙ and GSH) were observed (Fig. 3a). GSH and GSO˙ are the pair products from DSB dissociation, as shown in the possible cleavage pathway (Scheme S1†). Cascade oxidation products of GSH were also observed (Fig. 3a), as GSH could be easily oxidized. The ratio of GSO˙ to GSSG increased with elevation of the pulsed high voltage amplitude (Fig. 3b). Similar trend was observed with the ratio of GSH to GSSG (Fig. S3†). It should be noted that the increase amplitudes of GSH and GSO˙ showed distinct difference. Presumably, it is mainly because the short-lived GSO˙ is not easily observed in mass spectrometer. The effect of frequency on DSB cleavage showed similar trend with that of amplitude of the pulsed spray voltage (Fig. S4†). Similar DSB dissociations were also found for other DSB-containing peptides (selectin binding peptide and somatostatin). The above observations indicated that DSB of GSSG was increasingly dissociated with increased amplitude of the pulsed high voltage. Moreover, the presence of volatile electrolyte (NH4OAc or NH4HCO3) of low concentration (<10 mM) in spray solution would greatly facilitate the DSB dissociation (Fig. S5†).
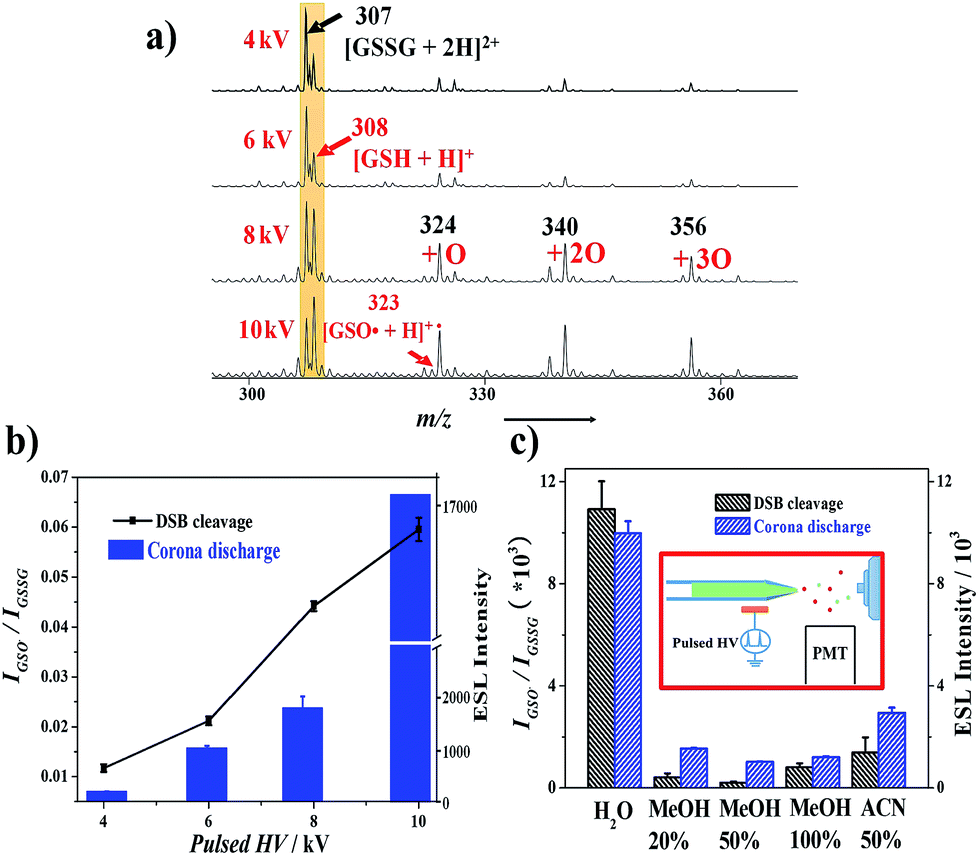 | ||
| Fig. 3 Dependence of oxidative cleavage reaction on corona discharge. (a) Effect of the amplitude of the pulsed high voltage (1 kHz) on disulfide cleavage of GSSG (8 μM). (b) Relationship between DSB cleavage and corona discharge. (c) Solvent-effect on DSB cleavage in InESI (pulse, 1 kHz, 6 kVp–p). Inset in (c) shows the characterization of corona discharge in InESI with a photomultiplier tube (PMT). In all these experiments, NH4OAc (5 mM) was added into the spray solution. For the identification of cleavage products, especially for [GSO˙ + H]˙+, CID MS/MS (Fig. 2c) was used and thus isotopic interfere from near peaks can be avoided. For all the characterization of cleavage degree, intact GSSG signal with minimal reaction (4 kVp–p) was used as the denominator to make a relative fair compare. | ||
Relationship between pulsed high voltage and corona discharge was also characterized. Although HO˙ from corona discharge is assumed to be responsible for DSB cleavage in this study, its presence in this system cannot be verified by MS detection. Thus we measured the electrospray-based light emission (ESL), a known sign of corona discharge,36 to evaluate the corona discharge during InESI. In a dark room, a PMT was placed under the spray tip to measure the ESL (inset in Fig. 3c), which is in accordance with previous investigation on corona discharge in low current (<0.2 μA) ESI.36 ESL signals increased in accordance with the elevation of the amplitude of pulsed high voltage, indicating corona discharge is enhanced gradually (Fig. 3b). Thus, the relationship between pulsed high voltage and corona discharge in InESI could be directly characterized by measuring the ESL with a PMT.
We then turned to investigate the relationship between DSB cleavage and corona discharge. In regard to solvent-effect, significant radical-induced DSB cleavage could be observed only in pure H2O solution, addition of organic solvents would result in dramatic decreasing of the cleavage product (Fig. 3c). That might be attributed to the organic solvents used in our system (methanol and acetonitrile) might also serve as electron scavengers and thus hinder enough corona discharge for HO˙ generation. Therefore, weak corona discharge would be found for organic solvents even with the presence of additional electrolyte but strong corona discharge for pure H2O solution. As a result, the radical induced cleavage of DSB could only be observed in H2O solution, which facilitates corona discharge generation. Except for solvent-effect, the effect of pulsed high voltage amplitude on DSB cleavage was also compared in the aspect of corona discharge. The increase trend for the dependence of DSB cleavage on the amplitude of pulsed high voltage (Fig. 3a/b) was well in accordance with that of corona discharge (Fig. 3b), indicating corona discharge is related with amplitude of the pulsed high voltage and DSB cleavage. Furthermore, to exclude the possibility of DSB cleavage prior to electrospray, sonic spray ionization (free from corona discharge due to the absence of high voltage) was introduced as a control experiment and no obvious cleavage product ions (GSH or GSO˙) were observed (Fig. S6†). In short, DSB cleavage via oxidative cleavage reaction in InESI is closely related to the existence of corona discharge.
Relationship of DSB cleavage with pulsed high voltage, pulsed high voltage with corona discharge, and DSB cleavage with corona discharge were proved successively. InESI-based DSB cleavage was thus demonstrated to be the result of oxidative cleavage reaction between DSB and HO˙, which is originated from the enhanced corona discharge.
To further investigate the capability to uncover backbone sequence of native DSB protein, we eventually tested human insulin with InESI-CID-MS protocol. It is challenging to obtain structural information from intact insulin precursor ions by conventional nanoESI-CID-MS because three DSBs (Fig. 4A insert) protect a large number of the peptide backbone (28 out of 51). Similar to the GSSG “cleavage ON/OFF” experiment, sequence information beyond DSBs could be obtained in “OFF” mode with conventional nanoESI (Fig. S7†). Then, in “ON” mode with typical pulsed high voltage turned on for InESI, through CID spectrum of HO˙ additive ion ([M + 5H + OH]5˙+) (Fig. 4A), sequence information “buried” within the two inter DSBs was revealed (Fig. 4B and C). Cleavage pathways similar with previous study46 were identified by fragment ions of OABb174+, OABb83+, OABy184+, OABb164+ and so on. The total sequence coverage obtained via the “ON/OFF” switch is shown in the insert of Fig. 4, 19 cleavages within the DSB were identified with InESI (marked red), which could not be provided by conventional nanoESI. In total, 30 out of 49 possible backbone cleavages (61%) are identified by InESI-CID-MS, exceeding that of conventional nanoESI-CID-MS (22%), which is comparable with that of ECD (49%)47 and ETD (64%).48 Thus, extensive sequence information of DSB-linked native protein could be revealed by InESI-CID-MS protocol.
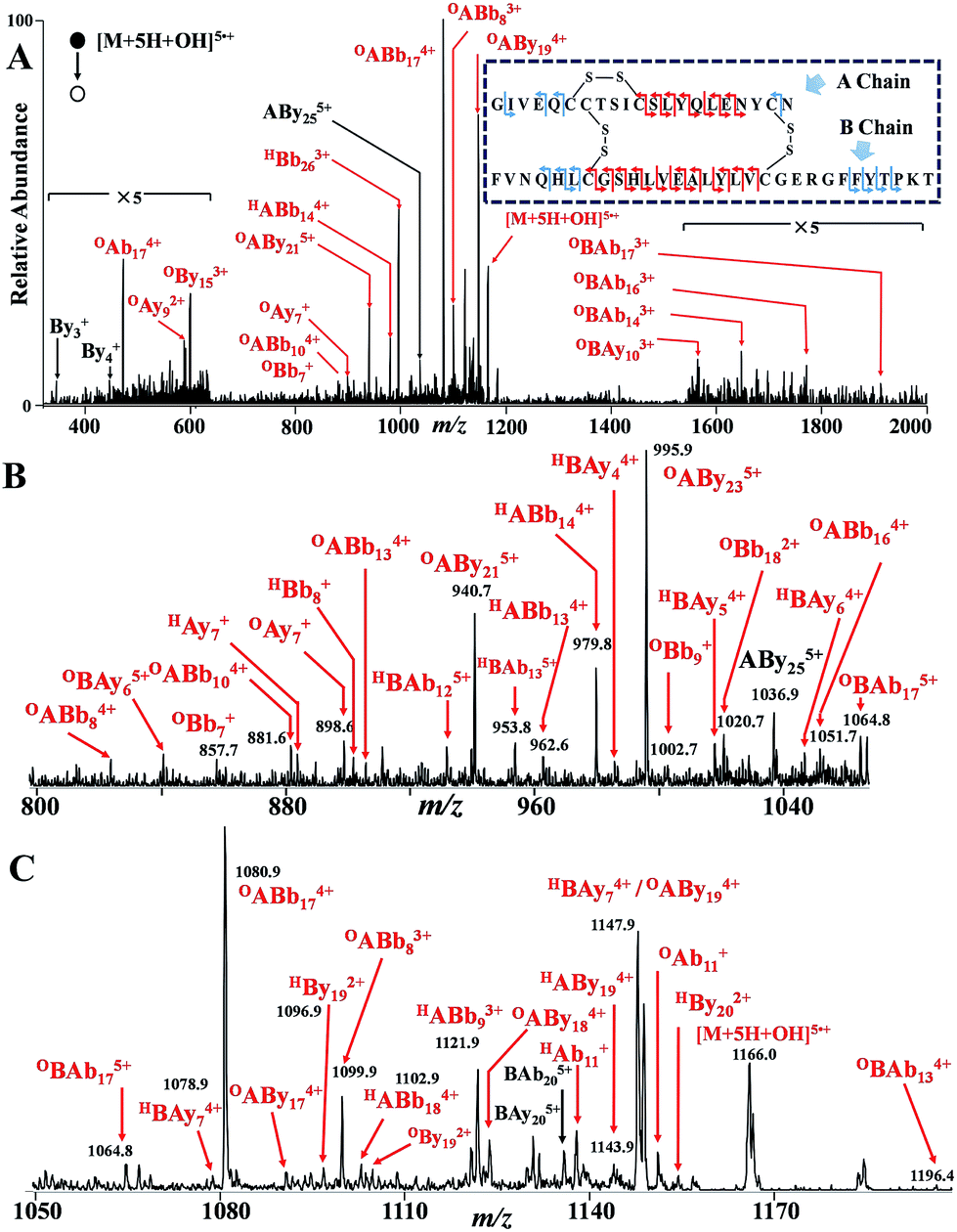 | ||
| Fig. 4 (A) Typical InESI-CID-MS spectrum of HO˙ additive precursor ion ([M + 5H + OH]5˙+, m/z 1166) of human insulin. (Insert: detailed sequence coverage) magnification of the above spectrum for mass range of (B) 800–1080 and (C) 1050–1200 to show the detailed fragment ions. Reaction MS1 spectrum and isotopic distribution seen in Fig. S8.† Fragment ions marked red are obtained from peptide backbone cleavages within DSB-protected region. | ||
In summary, InESI was utilized to generate HO˙ for selective and in situ cleavage of DSBs for peptides and native proteins. Selective reaction between HO˙ (generated from corona discharge) and DSB is the key process in InESI-CID-MS protocol. Peptide sequence of in-between and beyond DSB could be successively distinguished simply by regulating the amplitude of induced high voltage in InESI-CID-MS protocol or by rapid switch between InESI and nanoESI. Although the reaction yield and the HO˙-induced disulfide scrambling should be treated with caution, the representative characteristics of InESI-CID-MS protocol are: (i) free of additional tedious reducing steps (in situ DSB dissociation) or additional devices for HO˙ generation (low temperature plasma) that should avoid the time- and sample-consuming wet chemistry, (ii) native and rapid sequencing of DSB-bridged peptides/proteins with extensive amino acid sequence information that would facilitate the high throughput application, which are vital for potential applications in the realm of proteomics.
In typical InESI-CID-MS experiments, the size of spray emitter was kept ∼5 μm. All experiments were carried out with a LTQ-Velos Pro (Thermo Fisher Scientific, CA, USA) mass spectrometer. Insulin (recombinant human, CAS no. 11061-68-0) was purchased from J&K Scientific Ltd. (Beijing, China). DTT, GSSG, selectin binding peptide and somatostatin were supplied by Sangon Biotech Co., Ltd (Shanghai, China). Other chemicals were obtained from Sinopharm Chemical Reagent Co., Ltd (Shanghai, China). No further purifications were performed for all reagents. Chemical reduction was performed by incubating the native protein/peptide standards at 37 °C with 10 time's excess of DTT at pH 7 for 4 hours. All solvents used in this study were of HPLC grade. Purified water was obtained from Milli-Q® Reference System (Millipore Corp., USA) with conductivity of 18.2 MΩ cm. More detailed instrumental parameters seen in ESI.†
Acknowledgements
The authors acknowledge support for this work by National Natural Science Foundation of China (21205111) and Recruitment Program of Global Experts (Thousand Young Talent Program).Notes and references
- V. Zabrouskov, Y. Ge, J. Schwartz and J. W. Walker, Mol. Cell. Proteomics, 2008, 7, 1838–1849 CAS.
- Y. Ge, I. N. Rybakova, Q. Xu and R. L. Moss, Proc. Natl. Acad. Sci. U. S. A., 2009, 106, 12658–12663 CrossRef CAS PubMed.
- H. Zhang and Y. Ge, Circ.: Cardiovasc. Genet., 2011, 4, 711 CrossRef PubMed.
- Y.-C. Wang, S. E. Peterson and J. F. Loring, Cell Res., 2014, 24, 143–160 CrossRef CAS PubMed.
- J. M. Thornton, J. Mol. Biol., 1981, 151, 261–287 CrossRef CAS.
- T. E. Creighton, BioEssays, 1988, 8, 57–63 CrossRef CAS PubMed.
- C. N. Pace, G. R. Grimsley, J. A. Thomson and B. J. Barnett, J. Biol. Chem., 1988, 263, 11820–11825 CAS.
- M. Matsumura, G. Signor and B. W. Matthews, Nature, 1989, 342, 291–293 CrossRef CAS PubMed.
- W. J. Wedemeyer, E. Welker, M. Narayan and H. A. Scheraga, Biochemistry, 2000, 39, 4207–4216 CrossRef CAS PubMed.
- A. Bindoli and M. P. Rigobello, Antioxid. Redox Signaling, 2013, 18, 1557–1593 CrossRef CAS PubMed.
- J. A. Loo, C. G. Edmonds, H. R. Udseth and R. D. Smith, Anal. Chem., 1990, 62, 693–698 CrossRef CAS.
- X. Han, M. Jin, K. Breuker and F. W. McLafferty, Science, 2006, 314, 109–112 CrossRef CAS PubMed.
- G. Wang and I. A. Kaltashov, Anal. Chem., 2014, 86, 7293–7298 CrossRef CAS PubMed.
- M. Cammarata, K. Y. Lin, J. Pruet, H. W. Liu and J. Brodbelt, Anal. Chem., 2014, 86, 2534–2542 CrossRef CAS PubMed.
- L. Sun, G. Zhu, Y. Zhao, X. Yan, S. Mou and N. J. Dovichi, Angew. Chem., Int. Ed., 2013, 52, 13661–13664 CrossRef CAS PubMed.
- G. Zhu, L. Sun, X. Yan and N. J. Dovichi, Anal. Chem., 2014, 86, 6331–6336 CrossRef CAS PubMed.
- M. S. Lowenthal, Y. Liang, K. W. Phinney and S. E. Stein, Anal. Chem., 2014, 86, 551–558 CrossRef CAS PubMed.
- X. Zhang, E. Y. T. Chien, M. J. Chalmers, B. D. Pascal, J. Gatchalian, R. C. Stevens and P. R. Griffin, Anal. Chem., 2010, 82, 1100–1108 CrossRef CAS PubMed.
- X. Yan, H. Zhang, J. Watson, M. I. Schimerlik and M. L. Deinzer, Protein Sci., 2002, 11, 2113–2124 CrossRef CAS PubMed.
- J. A. Burns, J. C. Butler, J. Moran and G. M. Whitesides, J. Org. Chem., 1991, 56, 2648–2650 CrossRef CAS.
- J. L. Stephenson, B. J. Cargile and S. A. McLuckey, Rapid Commun. Mass Spectrom., 1999, 13, 2040–2048 CrossRef CAS.
- S. Mysling, R. Salbo, M. Ploug and T. J. D. Jørgensen, Anal. Chem., 2014, 86, 340–345 CrossRef CAS PubMed.
- H. Permentier and A. Bruins, J. Am. Soc. Mass Spectrom., 2004, 15, 1707–1716 CrossRef CAS PubMed.
- S. Nicolardi, A. M. Deelder, M. Palmblad and Y. E. M. van der Burgt, Anal. Chem., 2014, 86, 5376–5382 CrossRef CAS PubMed.
- Y. Zhang, H. D. Dewald and H. Chen, J. Proteome Res., 2011, 10, 1293–1304 CrossRef CAS PubMed.
- R. A. Zubarev, N. L. Kelleher and F. W. McLafferty, J. Am. Chem. Soc., 1998, 120, 3265–3266 CrossRef CAS.
- H. Oh, K. Breuker, S. K. Sze, Y. Ge, B. K. Carpenter and F. W. McLafferty, Proc. Natl. Acad. Sci. U. S. A., 2002, 99, 15863–15868 CrossRef CAS PubMed.
- J. E. P. Syka, J. J. Coon, M. J. Schroeder, J. Shabanowitz and D. F. Hunt, Proc. Natl. Acad. Sci. U. S. A., 2004, 101, 9528–9533 CrossRef CAS PubMed.
- D. F. Clark, E. P. Go and H. Desaire, Anal. Chem., 2012, 85, 1192–1199 CrossRef PubMed.
- S. D. Patterson and V. Katta, Anal. Chem., 1994, 66, 3727–3732 CrossRef CAS.
- Y. Fukuyama, S. Iwamoto and K. Tanaka, J. Mass Spectrom., 2006, 41, 191–201 CrossRef CAS PubMed.
- H. Chen, L. S. Eberlin and R. G. Cooks, J. Am. Chem. Soc., 2007, 129, 5880–5886 CrossRef CAS PubMed.
- Y. M. Fung, F. Kjeldsen, O. A. Silivra, T. W. Chan and R. A. Zubarev, Angew. Chem., Int. Ed., 2005, 44, 6399–6403 CrossRef CAS PubMed.
- Y. Xia and R. G. Cooks, Anal. Chem., 2010, 82, 2856–2864 CrossRef CAS PubMed.
- C. A. Stinson and Y. Xia, Analyst, 2013, 138, 2840–2846 RSC.
- I. L. Kanev, A. Y. Mikheev, Y. M. Shlyapnikov, E. A. Shlyapnikova, T. Y. Morozova and V. N. Morozov, Anal. Chem., 2014, 86, 1511–1517 CrossRef CAS PubMed.
- G. Huang, G. Li and R. G. Cooks, Angew. Chem., Int. Ed., 2011, 50, 9907–9910 CrossRef CAS PubMed.
- G. Li and G. Huang, J. Mass Spectrom., 2014, 49, 639–645 CrossRef CAS PubMed.
- W. N. English, Phys. Rev., 1948, 74, 179–189 CrossRef.
- G. A. Dawson, J. Geophys. Res., 1969, 74, 6859–6868 CrossRef.
- J. Lloyd and S. Hessb, J. Am. Soc. Mass Spectrom., 2010, 21, 2051–2061 CrossRef CAS PubMed.
- A. N. Vilkov, V. V. Laiko and V. M. Doroshenko, J. Mass Spectrom., 2009, 44, 477–484 CrossRef CAS PubMed.
- J. Lloyd and S. Hess, J. Am. Soc. Mass Spectrom., 2009, 20, 1988–1996 CrossRef CAS PubMed.
- M. Chen and K. D. Cook, Anal. Chem., 2007, 79, 2031–2036 CrossRef CAS PubMed.
- S. D. Maleknia, M. R. Chance and K. M. Downard, Rapid Commun. Mass Spectrom., 1999, 13, 2352–2358 CrossRef CAS.
- P. A. Chrisman and S. A. McLuckey, J. Proteome Res., 2002, 1, 549–557 CrossRef CAS.
- R. A. Zubarev, N. A. Kruger, E. K. Fridriksson, M. A. Lewis, D. M. Horn, B. K. Carpenter and F. W. McLafferty, J. Am. Chem. Soc., 1999, 121, 2857–2862 CrossRef CAS.
- J. Liu, H. P. Gunawardena, T.-Y. Huang and S. A. McLuckey, Int. J. Mass Spectrom., 2008, 276, 160–170 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/c4ra12386g |
| This journal is © The Royal Society of Chemistry 2014 |