
Potential of grape seed-derived polyphenols extract for protection against testosterone-induced benign prostatic hyperplasia in castrated rats
Abstract
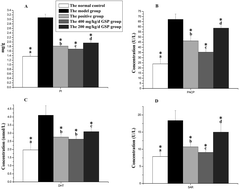
This study was to investigate the potential of grape seed-derived polyphenols extract (GSP) to protect against testosterone-induced benign prostatic hyperplasia (BPH) in castrated rats. After a 5 week experimental period, the prostatic levels of proinflammatory cytokines and plasma androgen level were measured by enzyme linked immunosorbent assay. Prostatic oxidative stress was evaluated by detecting the activities of antioxidant enzymes. Additionally, the prostatic levels of extracellular signal-regulated kinases (ERK), p38, protein kinase B (PKB/AKT), nuclear factor (NF)κB and intercellular cell adhesion molecule (ICAM) were determined using western blot analysis. It was found that GSP ameliorated the testosterone-induced high androgen level, over-expressions of NFκB and ICAM, and the high phosphorylation levels of ERK, p38 and AKT, as well as normalized antioxidant enzyme activities and regulated the proinflammatory cytokines. These results suggested that GSP had prostatic protective nature via regulating the androgen-MAPK/AKT-ICAM pathway and eventually alleviating the prostatic inflammatory responses and oxidative stress.
 Please wait while we load your content...
Please wait while we load your content...

