
Anti-inflammatory and hypoglycemic efficacy of Poria cocos and Dioscorea opposita in prediabetes mellitus rats
 ab
ab
Abstract
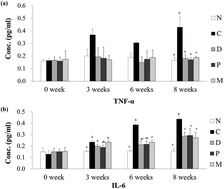
Poria cocos (Fu Ling) and Dioscorea opposita (Chinese Yam) were suggested to have potential benefits in blood sugar control. However, little is known about the underlying mechanisms. In this study, we investigated the anti-inflammatory and hypoglycemic effects of Poria cocos and Dioscorea opposita extracts in prediabetic rats. Fifty streptozotocin-injected rats with a mimic prediabetic status were gavaged with a single dose of either D. opposita (0.35 g kg−1 per day) or P. cocos (0.14 g kg−1 per day), a combination dose comprising single doses of the 2 herbal extracts, or vehicle for a 6-week treatment. Each group contained 10 rats. Blood and selected organ samples were collected during the study. The results indicated that administering the extracts singly or in combination for 6 week, significantly reduced the fasting blood-glucose level. The levels of interleukin-6 in plasma and selected organs significantly decreased during the treatment. The concentration of plasma free fatty acids (FFA) and the ratio of n − 6/n − 3 polyunsaturated fatty acids (PUFAs) were also significantly lowered after the 6 week intervention. The results indicate that administering P. cocos and D. opposita extracts produced anti-inflammatory effects in prediabetic rats by reducing the levels of interleukin-6 and the FFA ratio of n − 6/n − 3 PUFAs.
 Please wait while we load your content...
Please wait while we load your content...

