
Elucidating the endocytosis, intracellular trafficking, and exocytosis of carbon dots in neural cells†
Nan Zhoua,
Shoujun Zhub,
Suraj Maharjana,
Zeyu Haoa,
Yubin Songb,
Xiaohuan Zhaob,
Yanfang Jiangc,
Bai Yang*b and
Laijin Lu*a
aDepartment of Hand Surgery, First Hospital of Jilin University, Changchun 130021, China. E-mail: lulaijin@hotmail.com; Fax: +86 431 84808391; Tel: +86 431 84808260
bState Key Laboratory of Supramolecular Structure and Materials, College of Chemistry, Jilin University, Changchun 130012, China. E-mail: byangchem@jlu.edu.cn
cDepartment of Central Laboratory, First Hospital of Jilin University second part, Changchun 130032, China
First published on 12th November 2014
Abstract
Carbon dots (CDs) due to their ultrasmall size, excellent optical properties, chemical stability, biocompatibility and most importantly possession of low toxicity have been widely used as a fluorescent probe. However, many studies on CDs are focused on their biomedical applications, the cellular behavior of CDs remains unrevealed and poorly understood. In this study, we used our recently synthesized highly photoluminescent CDs (ca. 1–5 nm) and studied their cellular uptake mechanism, intracellular distribution and exocytosis in neural cells with the help of various techniques such as confocal laser scanning microscopy, fluorescence activated cell sorter, atomic force microscopy, and transmission electron microscopy. Our results suggest that CDs exhibit low cytotoxicity and highly efficient internalization in neural cells. Cellular uptake of CDs is dose, time and partially energy-dependent along with the involvement of passive diffusion. CDs are endocytosed via caveolae-mediated and clathrin-mediated pathways. Internalized CDs are dispersed in cytoplasm and nucleus, and most of them accumulate in endo-lysosomal structures and Golgi apparatus. CDs were actively transported to the cell periphery and exocytosed with a half-life of 2 h.
1. Introduction
Carbon dots (CDs), a new class of fluorescent carbon-based nanomaterials, have increasingly drawn attention due to their excellent optical properties, good chemical stability, biocompatibility and low toxicity.1 Many synthesis methods of CDs have been developed and generally divided into top-down nano-cutting methods and bottom-up carbonization approaches.2 Top-down nano-cutting methods include cutting different carbon resources like graphite, carbon fiber, carbon nanotubes, and graphite electrode.3,4 Bottom-up carbonization approaches always contain carbonization of carbohydrate, natural substances and polymers.5–7 The fascinating physical and chemical properties promote a large number of applications in the field of bioimaging,8,9 phototherapy,10 biosensors,11,12 drug delivery,13,14 and gene delivery.15By contrast, biological behavior of CDs, especially with respect to their cellular uptake mechanism and intracellular distribution, which is the foundation of these biomedical applications, is still not clearly understood. Sun's group firstly demonstrated that CDs passivated with PPEI-EI labeled both the cell membrane and the cytoplasm of MCF-7 cells without reaching the nucleus. There was no significant internalization of CDs at 4 °C, suggesting that a temperature-dependent endocytosis was likely to exist.16 Whereas Ray and co-workers found that 2–6 nm CDs without any further surface passivation following acidic oxidation could widely disperse in both cytoplasm and nucleus after 30 minutes incubation.17 It implied that surface passivation of CDs might influence their cellular internalization. Subsequently, Goh et al.18 reported that positively charged CDs with amine groups were effectively uptaken to the cells and CDs conjugated to negatively charged hyaluronic acid were internalized by receptor-mediated endocytosis. Moreover, Liu et al.19 demonstrated that after 3 h transfection, most PEI-passivated CDs uptaken appeared in endosomal compartments; while some complexes had already successfully escaped from endosome in such short time, which showed a high efficient gene transfection of CDs. Evidences to date suggest that an endocytosis mechanism, which is an active transport occurring at the cell surface and involving internalization of extracellular materials, participates in cellular uptake of CDs. However, elucidating the exact mechanism of CDs uptake by cells still requires more investigations.20 The specific endocytic mechanisms of nanoparticles in non-phagocytic cells generally include clathrin-mediated, caveolae-mediated, clathrin/caveolae-independent endocytosis and macropinocytosis.21,22 The cellular internalization of nanoparticles not only depends on the size, surface charge, shape, surface modification of nanoparticles, but also on the cell type.23–27 After uptake of nanoparticles by cells, they are interacted with different subcellular components and organelles, leading to their delivery into different intracellular organelles, which is directly related to the cytotoxicity or medical functions of the internalized nanoparticles.28 Therefore, the exact endocytic mechanism and intracellular location of CDs is highly indispensable for assessing their biological properties and to improve further understanding in their biomedical applications.
In our previous study, we synthesized highly photoluminescent CDs and successfully used in cell labeling.29 Herein, to study the internalization mechanism of CDs in neural cells, we cultured two neural cell types, including differentiated rat adrenal pheochromocytoma cells (PC12), derived from a neuroendocrine tumor of the sympathetic nervous system, and rat Schwann cells (RSC96), a type of neuroglial cells in peripheral nervous system.30–32 Confocal laser scanning microscopy (CLSM), fluorescence activated cell sorter (FACS) and atomic force microscopy (AFM) were used to study the uptake kinetics and endocytic pathways. To further determine the intracellular trafficking and distribution, we assessed the colocalization of CDs with endocytic markers and organelles specific dyes by using CLSM and directly observed their existence in cellular ultrastructures by using transmission electron microscopy (TEM). The exocytosis of internalized CDs was studied using CLSM and FACS. The findings derived from this study not only provide detailed information about the cellular behavior of CDs in neural cells but also show their good biocompatibility and great potential in biomedical applications, particularly in bioimaging of the nervous system.
2. Results and discussion
2.1. Fabrication and characterization of CDs
Our group previously reported a facile and high-output strategy for the fabrication of polymer-like CDs with quantum yields as high as ca. 60–80% which is one of the highest values reported till date for fluorescent carbon-based materials, and we successfully utilized this kind of high photoluminescent CDs in multicolor patterning, biosensors, and cell imaging.29 As shown in Fig. 1a, The CDs were prepared by hydrothermal reaction of citric acid and ethylenediamine. The TEM image showed that the CDs had uniform dispersion without any aggregation and the diameter of particles is between 1–5 nm (Fig. 1b). The XRD patterns of the CDs (Fig. 1c) displayed a broad peak, which proved the amorphous aggregation nanoparticle of the CDs. In the FTIR analysis of CDs, stretching vibrations of C–OH at 3430 cm−1, C–H at 2923 cm−1 and 2850 cm−1, as well as bending vibrations of N–H at 1570 cm−1 and a vibrational absorption band of C![[double bond, length as m-dash]](https://www.rsc.org/images/entities/char_e001.gif) O at 1635 cm−1 were observed (Fig. 1d), which endowed the solubility of the CDs for bio-based applications. Furthermore, the CDs possessed optimal excitation and emission wavelengths at 360 nm and 443 nm, and showed blue color under a hand-hold UV lamp (Fig. 1e). The PL center was proved a kind of molecule state. The z-potential of the CDs was ca. +2.3 mV due to the excess amine-based groups. For the application of cell imaging, we detected the photostability of CDs in cells using CLSM, and found that PL intensities of CDs could remain constant within 2 min continuous scanning (Fig. S1, ESI†).
O at 1635 cm−1 were observed (Fig. 1d), which endowed the solubility of the CDs for bio-based applications. Furthermore, the CDs possessed optimal excitation and emission wavelengths at 360 nm and 443 nm, and showed blue color under a hand-hold UV lamp (Fig. 1e). The PL center was proved a kind of molecule state. The z-potential of the CDs was ca. +2.3 mV due to the excess amine-based groups. For the application of cell imaging, we detected the photostability of CDs in cells using CLSM, and found that PL intensities of CDs could remain constant within 2 min continuous scanning (Fig. S1, ESI†).
 | ||
| Fig. 1 The polymer-like CDs used for cellular uptake. (a) The scheme of the prepared CDs from citric acid and ethylenediamine. (b) The TEM image of CDs. (c) The XRD pattern of CDs. (d) The FTIR spectra of CDs. It contains the –OH, epoxy, CO, C–N (CN), NH and CH groups. (e) The optical properties of the CDs, the inset is a photograph of CDs in aqueous solution under UV light. | ||
2.2. Cellular uptake kinetics of CDs
Firstly, we evaluated the cytotoxicity of CDs in PC12 and RSC96 cells. After 24 h incubation with CDs (10–500 μg mL−1), the cell viability remained above 90% at concentration ranging from 10 to 200 μg mL−1 and it marginally decreased but still remained above 80% even at higher concentrations of 300 and 500 μg mL−1 (Fig. S2, ESI†). The low cytotoxicity is one of the most important characteristics of CDs, which was similar with Zhao's study showing that bare CDs were obviously noncytotoxic up to relatively high concentration of 400 μg mL−1.33 This fascinating advantage gives CDs a great potential in future clinical application when compared to QDs composed of toxic heavy metal elements such as cadmium.1 Subsequently, PC12 and RSC96 cells were incubated with different concentrations of CDs (10–500 μg mL−1) for 2 h, and then the intracellular fluorescent intensity of CDs was measured using FACS. It showed that the intracellular amount of CDs was linearly dependent on the concentration (r2 > 0.95) and no saturation of the uptake was observed at 10–500 μg mL−1 of CDs (Fig. 2). Previous study had reported that the cellular uptake of graphene quantum dots (GQD) by neural stem cells was dose-dependent, however, the uptake of GQD achieved saturation at 200 μg mL−1.34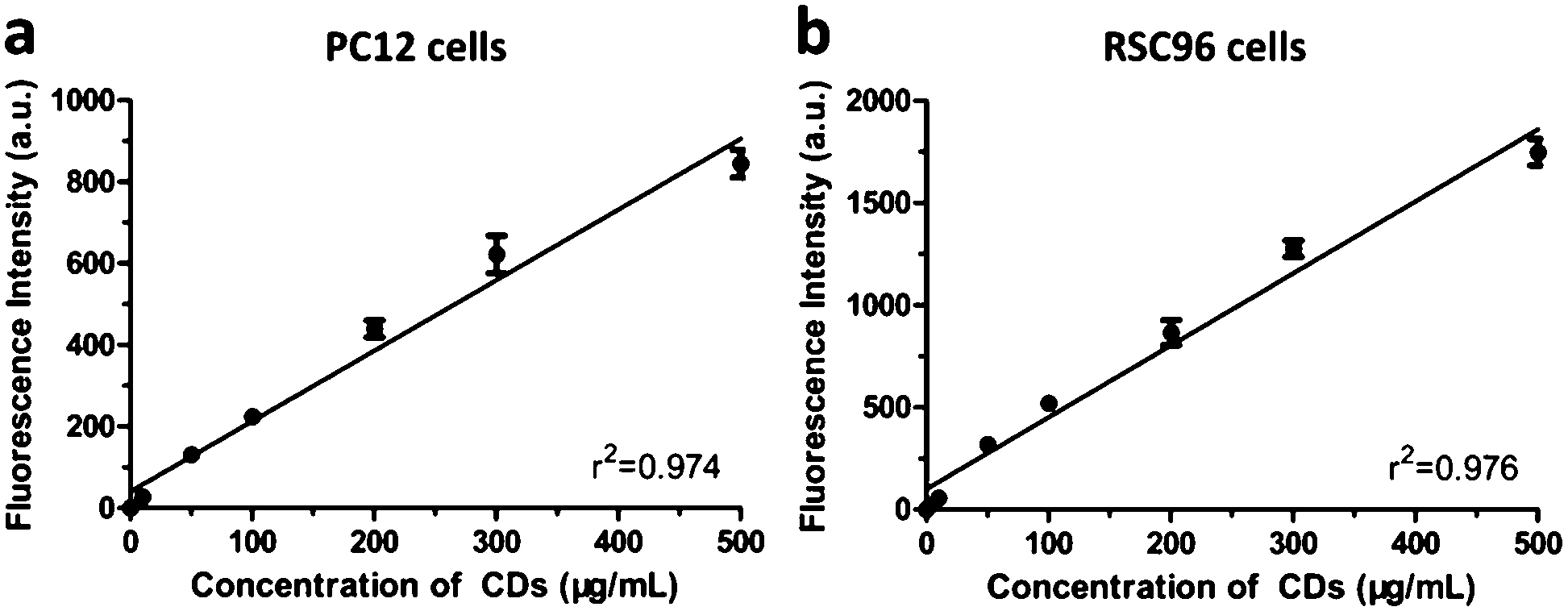 | ||
| Fig. 2 Dose dependent cellular uptake of CDs measured by FACS. The values are expressed as mean ± SD (n = 3). | ||
To study uptake kinetics, cells were incubated with 200 μg mL−1 CDs for different time periods (5–360 min) and amount of internalized CDs was detected using CLSM and FACS. As shown in Fig. 3a and b, after only 5 min incubation, CDs could distribute in cytoplasm without accumulating on the cell membrane and without entering nucleus. Over time, the fluorescence intensity of cells, including cytoplasm and nucleus, would enhance and we observed that CDs began to appear inside the nucleus in 15 min. After incubating with CDs for 360 min, it was clearly seen that a large amount of CDs located in the cytoplasm and a little amount inside the nucleus. In fact, similar phenomena were observed in previous studies, such as CDs (1–5 nm) synthesized via nitric acid oxidation of carbon soot,17 PEI-passivated CDs (4–12 nm),19 cationic quaternized CDs (few nm).35 Since the size exclusion of nuclear pore limit for the passive diffusion of macromolecules was about 9 nm and the size of CDs was 1–5 nm, we speculated that CDs entered the nucleus probably via passive diffusion through the nuclear pores.36 According to the densitometric analysis in Fig. 3c and d, the uptake rate of CDs within 30 min was extremely high, a significant deceleration occurred between 30 min and 120 min, then the uptake of CDs reached possible saturation after 120 min. Dynamic confocal imaging of endocytosis of CDs was also measured in RSC96 cells (Fig. S16†). Moreover, we used FACS to confirm above results and accidentally found that serum could inhibit cellular uptake of CDs. The time-dependent uptake of CDs was observed in both serum free media (SFM) and complete culture media (cMEM, DMEM supplemented with 10% FBS). But at each time point, the amount of internalized CDs in cMEM was less than that in SFM (Fig. 3e and f). We found that cell culture media caused the fluorescence quenching of CDs (Fig. S3, ESI†), probably due to the sabotage of molecule like PL center according to our previous study.37,38 But the inhibitory effects of SFM and cMEM on PL intensity of CDs were similar. We also found that the increase of CDs uptake in SFM could hardly affect the cell viability after 6 h or 24 h incubation (Fig. S4, ESI†). Several studies on inhibitory effect of serum on cellular uptake of nanoparticles, such as gold nanoparticles, silica nanoparticles and dihydrolipoic acid-coated quantum dots, suggested that a protein adsorption layer on nanoparticle surface played an essential role in their cellular uptake.39–41 The formation of protein corona on the surface of CDs was observed by SDS-PAGE (sodium dodecyl sulfate polyacrylamide) gel electrophoresis (Fig. S9 and S10†). Therefore, we suggested that stronger adhesion of CDs to cell membrane in SFM caused higher internalization and the different activations of growth factor receptors in SFM and cMEM could also affect the level of CDs endocytosis.
 | ||
| Fig. 3 Cellular uptake kinetics of CDs. (a and b) CLSM images of cells incubated with CDs for 0, 5, 15, 30, 60, 120, 360 min (the scale bar is 10 μm); (c and d) fluorescence intensity of internalized CDs in the whole cell and nucleus is quantified and used to calculate the amount of CDs endocytosed; (e and f) FACS analysis of CDs uptake kinetics in serum free media (SFM) and complete culture media supplemented with 10% serum (cMEM). The values are expressed as mean ± SD (n = 3). | ||
2.3. Cellular uptake pathways of CDs
Previous studies reported that the endocytosis, which was an energy-dependent active transport, participated in the cellular uptake of CDs.16–20 However, the distinct pathway involved in the process is still unknown till present. So we firstly used FACS to quantitatively analyze the cellular uptake of CDs at low temperature (4 °C) or in ATP-depleted environments with use of sodium azide (SA) and 2-deoxy-d-glucose (2-DG). As shown in Fig. 4, comparing with endocytosis of CDs at 37 °C, uptake of CDs by PC12 cells at 4 °C and ATP-depletion reduced to 56.4 ± 2.6% (P < 0.001) and 68.1 ± 7.6% (P < 0.001), respectively; and that of RSC96 cells reduced to 62.7 ± 2.3% (P < 0.001) and 59.1 ± 6.9% (P < 0.001), respectively. CLSM images also proved the inhibition of cellular uptake of CDs at 4 °C and ATP-depletion (Fig. S5, ESI†). As it is well known, at low temperature there is a decrease in various enzyme activities thereby causing a reduction in energy production in mitochondria and whereas SA and 2-DG create an ATP-depleted environment by blocking respiratory chain of mitochondria and interfering carbohydrate metabolism, respectively.42,43 Therefore, reduced uptake of CDs at 4 °C and ATP-depletion revealed an energy-dependent endocytosis of CDs. However, internalization of CDs still seen at 4 °C and ATP-depletion indicated the existence of energy-independent pathways as well, for example passive diffusion.66–69 Similar phenomena was also reported in several ultrasmall nanoparticles, such as gold nanoparticles (less than 10 nm),44 sub-5 nm gadolinium-based nanoparticles.45 It suggested that the cellular uptake of CDs (1–5 nm) was only partially energy-dependent and passive diffusion also participated in the process. | ||
| Fig. 4 Quantitative analysis of influence of uptake inhibitors for cellular uptake pathway. MβCD: methyl-β-cyclodextran; CytoD: cytochalasin D. The values are expressed as mean ± SD (n = 3). ***P < 0.001, compared with 37 °C group, using one-way ANOVA. | ||
Subsequently, we used different uptake inhibitors to determine which pathway was mainly involved in the endocytosis of CDs. Before that, we needed firstly evaluate the cytotoxicity of inhibitors to determine their appropriate concentration (Fig. S6, ESI†). The cell viabilities of PC12 and RSC96 cells incubated with different inhibitors (2 mM of methyl-β-cyclodextran, 50 μg mL−1 of genistein, 10 μg mL−1 of chlorpromazine, 50 μM of amiloride, and 5 μM of cytochalasin D) were more than 85%, suggesting that these inhibitors exhibited negligible cytotoxicity and it was able to eliminate the effect of non-specific cytotoxicity on cellular uptake. Next, we evaluated the effect of inhibitors on endocytic pathways of CDs by FACS (Fig. 4). Firstly, methyl-β-cyclodextran (MβCD), depleting cholesterol from the cell membranes, and genistein, a tyrosine kinase inhibitor blocking the phosphorylation of caveolin-1, were used to inhibit caveolae-mediated endocytosis.46 It revealed a significantly decreased uptake of CDs (P < 0.001) with approximately a 30% inhibition rate, suggesting that caveolae-mediated endocytosis was a primary uptake pathway of CDs. Secondly, chlorpromazine, which inhibited clathrin-coated pit formation by relocating clathrin and its adapter proteins from the plasma membrane to the endosomes, was used to inhibit clathrin-mediated endocytosis.46 For CDs, an inhibition rate of approximately 20% was observed in the chlorpromazine-treated cells, indicating that clathrin-mediated endocytosis was one of uptake pathways of CDs. Last, we used amiloride, interfering with membrane Na+/H+ ATPase, and cytochalasin D (CytoD), inhibiting F-actin polymerization,47 to suppress the macropinocytosis. No obvious inhibition of CDs uptake was observed, demonstrating macropinocytosis didn't play an important role in the uptake of CDs. Uptake pathway of CDs was also investigated in cMEM (Fig. S11†) and we found that CDs were endocytosed via caveolae, clathrin-mediated pathways in the presence and absence of serum.
We further used AFM to study the cell surface morphological change at the initial stage of uptake of CDs by RSC96 cells. Cells not treated with CDs were taken as a control and the smooth surfaces were observed in Fig. 5a. After exposure to CDs for 15 min, obvious invagination appeared on the cell surface (Fig. 5b). The AFM 3D structures (Fig. 5c and d) clearly displayed them. We further measured the diameter and depth of three representative invagination structures (Fig. 5e), and they exhibited ca. 250–300 nm in diameter and ca. 30–50 nm in depth. Similar result was also found in the uptake of fullerenic nanoparticles.48 The morphological changes of cell membrane also verified the characteristic features of the endocytosis process in the uptake of CDs.
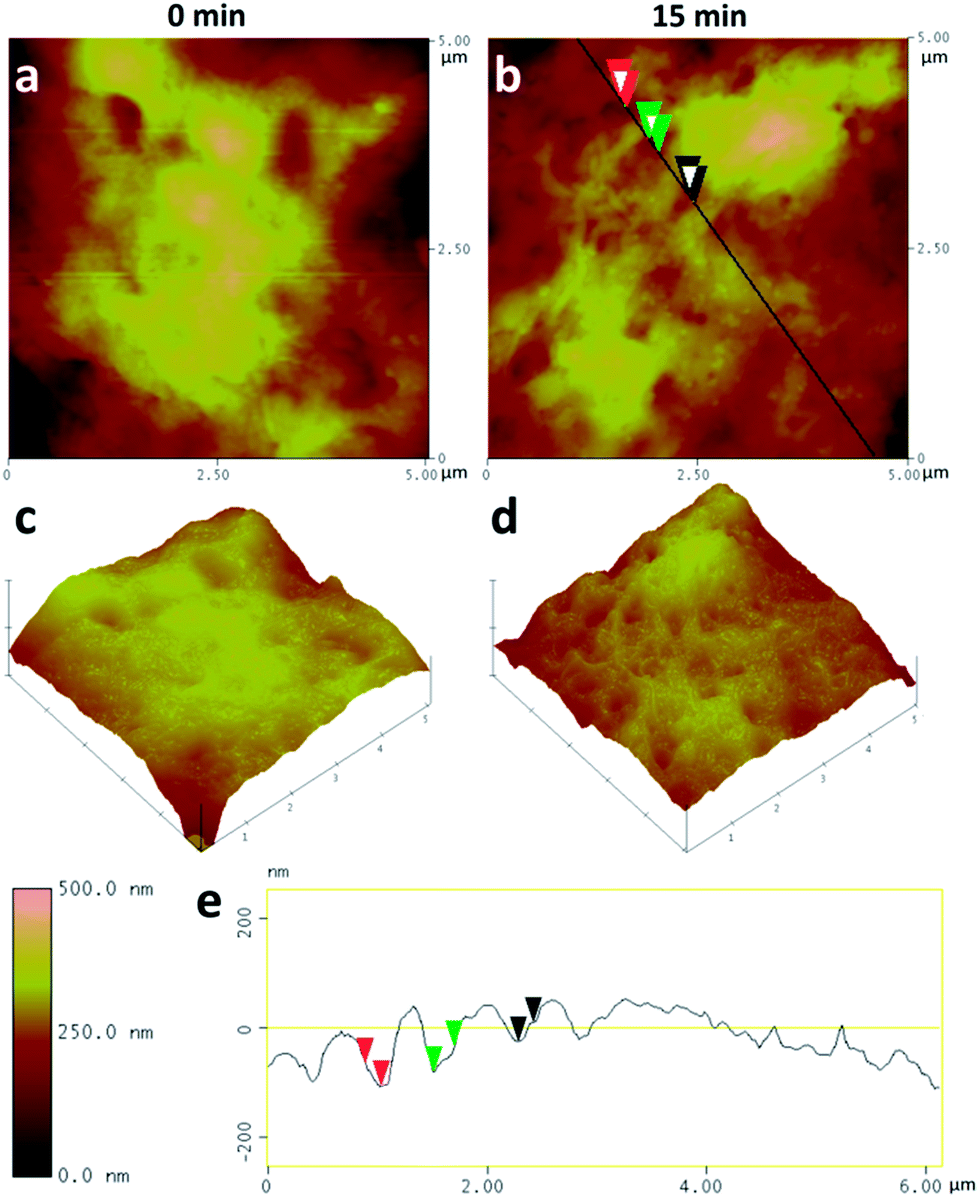 | ||
| Fig. 5 AFM scanning (tapping mode) of the morphological changes of RSC96 cell surface. Cell surface changes after incubation with CDs for 0 min (a) and 15 min (b). The images in the second row (c and d), corresponding to the upper row, exhibit 3D AFM images of cell surfaces. The diameter and depth of three representative invagination structures (e) are measured. | ||
Widely known, each endocytic pathway has its particular structural characteristics, such as the flask-shaped structures (60–80 nm) in caveolae, which can restrict internalization of large nanoparticles (>100 nm), clathrin-coated pits with an upper limit of about 200 nm diameter and macropinosomes which are larger (0.5–10 μm) than the other two pits and can internalize large particles with submicron.49–52 So the size of nanoparticles dominates their properties and also determine the processes of cellular uptake.28 Previous studies on the uptake of GQD (average lateral size is 20 nm) showed that it was both energy-independent and energy-dependent, and caveolae-mediated endocytosis might be the primary pathway.53 Another group reported that 50–100 nm SWNTs could be internalized through clathrin-coated vesicles as well as the caveolae pathway and short SWNTs (<50 nm) could partly enter into cells directly through an energy-independent pathway.54 Therefore, we suggested that the ultrasmall size of CDs determined their partially energy-dependent uptake and two major pathways referring to caveolae-mediated, clathrin-mediated pathways.
2.4. Intracellular trafficking and distribution of CDs
We firstly performed tomographic scanning by using CLSM, which clearly distinguished the intracellular distribution of CDs. As shown in Fig. S7,† blue fluorescence of CDs within cells was observed in each slice, suggesting that CDs were distributed inside the cytoplasm and nucleus instead of adhering only to the cell surfaces.Subsequently, we used endocytic markers, such as cholera toxin subunit B (CTB), transferrin and dextran, to assess colocalization of CDs with caveolae-mediated, clathrin-mediated, and macropinocytic pathways, respectively.55 As shown in Fig. 6, CDs significantly colocalized with CTB and transferrin in the cytoplasm, suggesting that CDs could be endocytosed via caveolae-mediated, clathrin-mediated pathways and internalized CDs resided within caveosomes and endosomes. However, we didn't observe any intracellular colocalization between CDs and dextran in both cell types, which indicated that CDs were not endocytosed via macropinocytosis. As it is well known, CTB can be delivered to the Golgi complex via caveosomes and transferrin is always a marker of endosomes and can reach the lysosomes in the end.47,56,57 So we also incubated cells with CDs and cellular organelles specific markers, including LysoTracker, Golgi-RFP, ER-Tracker and MitoTracker. Fig. 7a, b, e, and f showed that the internalized CDs were primarily transported to lysosomes and Golgi apparatus in both cell types. We observed only minor intracellular colocalization with endoplasmic reticulum and mitochondria (Fig. 7c, d, g and h). Moreover, the intracellular distribution of CDs in cMEM seemed to be similar with that in SFM (Fig S12†).
 | ||
| Fig. 6 Intracellular trafficking of internalized CDs. CDs are co-localized with AF647-CTB, AF647-Tf and AF647-Dex in PC12 (a–c) and RSC96 (d–f) cells. Blue and red spots show the intracellular distribution of CDs and caveosomes, endosomes and macropinosomes, and pink spots in the merged images indicate colocalization of CDs with caveosomes and endosomes. No colocalization between CDs and macropinosomes are observed. The scale bar is 5 μm and representative areas are magnified optically. | ||
 | ||
| Fig. 7 CDs co-localize with cellular organelles specific dyes in PC12 (a–d) and RSC96 (e–h) cells. LysoTracker red (a and e), Golgi-RFP (b and f), ER-Tracker red (c and g) and MitoTracker red (d and h) were used to label lysosomes, Golgi apparatus, endoplasmic reticulum and mitochondria, respectively. Pink spots in the merged images indicate colocalization of CDs and lysosomes along with Golgi apparatus. The scale bar is 5 μm and representative areas are magnified optically. | ||
Finally, TEM was used to further image the endocytosis and intracellular distribution after 1 h exposure to CDs. The invagination of plasma membrane formed a clathrin-coated pit (Fig. S8b†), which proved the involvement of clathrin-mediated endocytosis. And an endocytic vesicle in the cytoplasm (Fig. S8e†) was also observed. In these endocytic structures, we found some medium electron-dense contrast particles with the size of 3–8 nm which were similar with CDs (Fig. 1b), compared with the low contrast cell background. Lysosomes (Fig. S8b and e†) were also observed in the cytoplasm, however, since they had higher contrast than CDs, it was difficult to distinguish CDs inside of lysosomes. It was worth noting that metallic and metal oxide nanoparticles (e.g., Au, Ag, TiO2 and SiO2) could generate much greater contrast in the TEM compared with cell background and they seemed to be much easier to be observed in TEM than our medium contrast CDs.58
Different properties of nanoparticles would influence their intracellular location and translocation, which were directly related to the cytotoxicity or medical functions of the internalized nanoparticles.28 As far as we know, few studies have elucidated the intracellular fate of CDs in detail. We firstly reported that internalized CDs located mainly within endo-lysosomal structures, Golgi apparatus and partly entered nucleus, only a small amount of CDs dispersed in ER and mitochondria. The ultrasmall size (1–5 nm) and positive charge (ca. +2.3 mV) of CDs probably contributed in entering cells efficiently (within 5 min).59 CDs were able to enter nucleus, which was probably helpful in gene delivery.19 Moreover, CDs translocated into lysosomes, which were acidic cellular compartments, via endocytic pathways probably due to their positive charge, and this property was very important to lysosomotropic delivery drug delivery for nanomedicine.60 Only minor distribution of CDs in mitochondria should explain their low cytotoxicity due to the negligible disturbance to respiratory chain of mitochondria. CDs could translocate to Golgi apparatus, probably indicating a further transport via retrograde trafficking pathways,60 which was crucial to the application of nanoparticles in the nervous system, such as neuroimaging and neural tracing.61–64
2.5. Exocytosis of internalized CDs
To study exocytosis of internalized CDs, the cells were first pre-incubated with CDs for 3 h, then washed with PBS and incubated with fresh culture media for 0.5, 1, 2, 4, 24, 48 and 72 h at 37 °C. Fig. 8a and b showed the significant decrease of intracellular fluorescence intensity of CDs with increasing incubation time. There were two possible explanations: (1) chemical degradation of CDs in the cells, and (2) release of CDs out of the cells through exocytosis. Lysosomes could provide acidic local environment (pH 4.5–5) to cause the chemical degradation of molecules.65 However, we previously reported that the PL intensities of CDs could remain constant in a solution of pH 4–11.29 Moreover, we measured the fluorescence intensity of CDs exocytosed into medium from cells at different time points (Fig. S13†) and the increasing intensity of extracellular medium might also proved the exocytosis of CDs. Thus, we suggested that the attenuation of intracellular luminescent signal was caused by the exocytosis of CDs.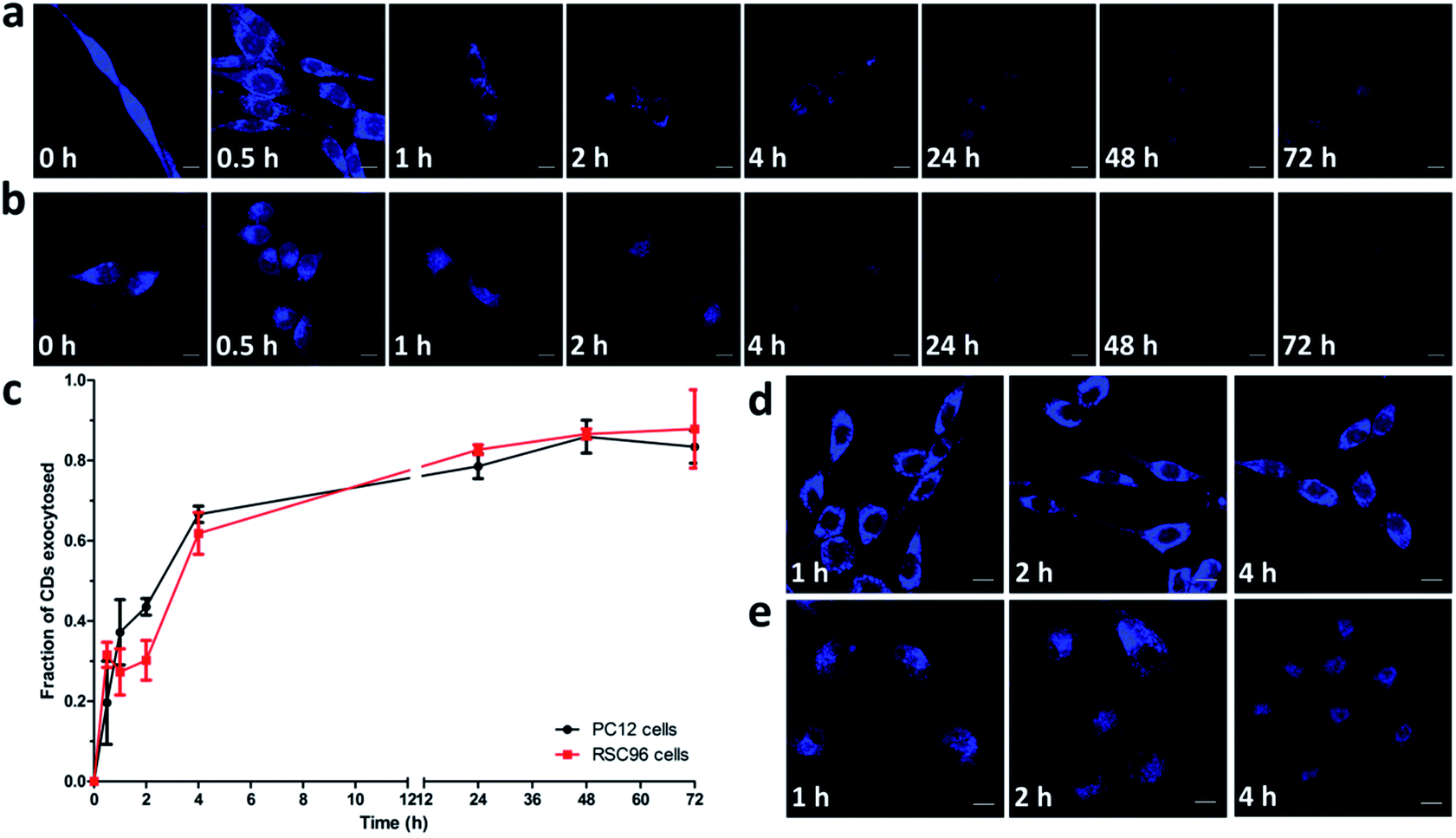 | ||
| Fig. 8 Exocytosis of internalized CDs. CLSM images of PC12 (a) and RSC96 (b) cells show the significant exocytosis of CDs after replacing extracellular CDs solution by fresh culture media at 37 °C. Kinetics of exocytosis of CDs is quantitatively measured by FACS (c) and the values are expressed as mean ± SD (n = 3). The exocytosis of CDs is inhibited at 4 °C in PC12 (d) and RSC96 (e) cells. | ||
Next, we used FACS to quantitatively measure the kinetics of exocytosis of CDs and we calculated the fraction of CDs exocytosed (Fexo) from the cells by using the following equation:
| Fexo = 1− It/I0 | (1) |
To further determine the energy-dependent exocytosis of CDs, the pre-treated cells were cultured with fresh media for 1, 2 and 4 h at 4 °C. Fig. 8d and e showed that the exocytosis of CDs was inhibited significantly at 4 °C compared with that at 37 °C, revealing an energy-dependent process, which was also proved by FACS analysis (Fig. S14†). Based on previous results, we speculated that CDs trapped in lysosomes and Golgi apparatus were actively transported to the cell periphery and exocytosed to the media. Finally, a schematic representation of endocytosis and exocytosis of CDs is depicted in Fig. 9.
 | ||
| Fig. 9 Schematic diagram illustrating the key steps involved in CDs uptake, intracellular trafficking and exocytosis. | ||
3. Experimental section
3.1. Materials
Fetal bovine serum (FBS), phosphate buffer solution (PBS), Dulbecco's Modified Eagle Medium (DMEM), penicillin–streptomycin, trypsin–EDTA, propidium iodide (PI), CellLight Golgi-RFP, MitoTracker Red, ER-Tracker Red, Lysosome-Tracker Red, Alexa Fluor 647-Transferrin (AF647-Tf), Alexa Fluor 647-Cholera Toxin Subunit B (AF647-CTB), and Alexa Fluor 647-Dextran 10 kDa (AF647-Dex) were obtained from Life Technologies. Sodium azide (SA), Methyl-β-cyclodextran (MβCD), genistein, amiloride, 2-deoxy-d-glucose (2-DG), chlorpromazine, tetrazolium salt (MTT) and dimethyl sulfoxide (DMSO) were purchased from Sigma-Aldrich. Cytochalasin D (CytoD) was obtained from Bioaustralis (Smithfield, NSW, Australia).3.2. Synthesis and characterization
CDs were prepared as follows: citric acid (1.0507 g) and ethylenediamine (335 μL) was dissolved in DI-water (10 mL). Then the solution was transferred to a poly (tetrafluoroethylene) (Teflon)-lined autoclave (30 mL) and heated at 200 °C for 5 h. After the reaction, the reactors were cooled to room temperature by water or naturally. The aqueous solution was evaporated by high temperature rotary evaporation to obtain the solid sample. The product, which was brown-black and transparent, was subjected to dialysis (out of the bag) in order to obtained the CDs. High-resolution transmission electron microscope (HTEM) was recorded on FEI Tecnai F20 (The decreased electro-beam intensity and increased exposure time can be beneficial to the obtained TEM images of CDs). Fluorescence spectroscopy was performed with a Shimadzu RF-5301 PC spectrophotometer. UV-vis absorption spectra were obtained using a Shimadzu 3100 UV-vis spectrophotometer. IR spectra were taken on a Nicolet AVATAR 360 FT-IR spectrophotometer. XRD investigation was carried out using a Siemens D5005 diffractometer. Zeta potential measurements were performed using a Zetasizer Nano-ZS (Malvern Instruments). Each sample was measured 5 times and the average data was presented. Photostability of CDs in cells was detected for 10 min using a FV1000 Olympus IX81 CLSM (Osaka, Japan) with a 40× objective lens to continuous scan at the same setting (Ex/Em: 405/430–470 nm, 18% laser intensity, HV 636, Gain 1.625, offset 27%).3.3. Cell culture
PC12 and RSC96 cells were purchased from the Cell Bank of the Chinese Academy of Sciences (Shanghai, China) and cultured in DMEM supplemented with 10% FBS, 100 U mL−1 penicillin, and 100 μg mL−1 streptomycin at 37 °C in a humidified atmosphere with 5% CO2.30–323.4. Cytotoxicity
PC12 and RSC96 cells were seeded in 96-well plates at 1 × 104 cells per well and incubated for 24 h at 37 °C and 5% CO2. Different concentrations of CDs (10, 20, 40, 60, 80, 100, 200, 300, 500 μg mL−1) was added and incubated for another 24 h. Then, the nanoparticle dispersion in each well was replaced by 180 μL serum free medium (SFM) and 20 μL MTT (5 mg mL−1). After 4 h incubation, the supernatant was removed by inverting the plates to decant the liquid; 200 μL per well DMSO was added to dissolve the crystals remaining at the bottom of plate. The absorbance was measured at 490 nm using a Synergy HT microplate reader (Bio-Tek, Winooski, VT). Cell viability was expressed as a percentage relative to untreated cells, which served as the control.3.5. Cellular uptake kinetics
PC12 and RSC96 cells were seeded and incubated overnight in 6-well plates (5 × 105 cells per well) for FACS analysis or in 35 mm glass-bottom dishes (5 × 104 cells per dish) for CLSM image. Incubation with different concentrations (10, 50, 100, 200, 300, 500 μg mL−1) of CDs for 2 h was prepared to examine the dose-dependent uptake using a FACSAria II flow cytometer from Becton Dickinson (New York, USA) with 405 nm laser. As for the FACS analysis, 10![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 000 events were collected for each sample. And incubation with CDs (0.2 mg mL−1) for different time (5, 15, 30, 60, 120, 360 min) was prepared to determine the time-dependent uptake using CLSM with a 40× objective lens (Ex/Em: 405/430–470 nm). Three different visual fields of each time point containing 30–40 cells were analyzed and triplicate experiments were done at three independent time points. The mean fluorescence intensity (MFI) of the whole cell and the nucleus were separately analyzed in all 9 images by Image J software. Time-dependent uptake of CDs was also performed by FACS analysis in the presence or absence of serum. Moreover, the cytotoxic effect of CDs in the presence or absence of serum was also evaluated using the MTT assay.
000 events were collected for each sample. And incubation with CDs (0.2 mg mL−1) for different time (5, 15, 30, 60, 120, 360 min) was prepared to determine the time-dependent uptake using CLSM with a 40× objective lens (Ex/Em: 405/430–470 nm). Three different visual fields of each time point containing 30–40 cells were analyzed and triplicate experiments were done at three independent time points. The mean fluorescence intensity (MFI) of the whole cell and the nucleus were separately analyzed in all 9 images by Image J software. Time-dependent uptake of CDs was also performed by FACS analysis in the presence or absence of serum. Moreover, the cytotoxic effect of CDs in the presence or absence of serum was also evaluated using the MTT assay.
3.6. Cellular uptake pathways
Firstly, the energy-dependent uptake was performed with low temperature incubation and ATP depletion treatments. PC12 and RSC96 cells were precooled at 4 °C or pretreated with 10 mM SA and 50 mM 2-DG at 37 °C for 45 min, then CDs were added and incubated for another 1 h. Intracellular fluorescence intensity of CDs was detected using FACS and CLSM. Secondly, different uptake inhibitors were used to determine the exact endocytosis mechanism. Before that, we used MTT assay to evaluate the cytotoxicity of inhibitors after 4 h incubation with MβCD (2 mM), genistein (50 μg mL−1), chlorpromazine (10 μg mL−1), amiloride (50 μM) and CytoD (5 μM). PC12 and RSC96 cells were pretreated with inhibitors in SFM for 45 min, CDs (0.2 mg mL−1) were then added and co-incubated for an additional 1 h prior to FACS analysis. The group treated neither with inhibitors nor CDs was used as a negative control and the group which was added CDs was used as a positive control. Finally, atomic force microscopy (AFM) was used to study the initial stage of CDs uptake. RSC96 cells were incubated overnight in chamber coverglass (5 × 104 cells) and were exposed to CDs for 15 min followed by 3 times washings with chilled PBS (pH = 7.4). Then the cells were fixed with freshly prepared 4% chilled paraformaldehyde in PBS for 20 min. After fixation, the cells were washed 3 times with chilled PBS and air-dried cells were scanned in tapping mode AFM with a Nanoscope IIIa scanning probe microscope from Digital Instruments.3.7. Intracellular trafficking and distribution
We firstly performed CLSM tomographic scanning of PC12 and RSC96 cells double labeled with CDs and PI. Cells were incubated with CDs for 2 h, then rinsed and fixed with 4% paraformaldehyde for 30 min at room temperature, protected from light. Next, cells were incubated with PI (2 μg mL−1) for 10 min followed by 3 times washing with PBS. Cells were examined using CLSM with a 60× oil immersion objective (Ex/Em: 405/430–470 nm for CDs; Ex/Em: 559/595–665 nm for PI). Z-series were collected at 0.5 μm intervals and 10 serial optical sections were obtained. Subsequently, we used endocytotic markers to colocalize with CDs to determine the intracellular trafficking of CDs. PC12 and RSC96 cells (5 × 104 cells per dish) were incubated with CDs for 1 h. After 3 times washings, cells were incubated with AF647-CTB (5 μg mL−1), AF647-Tf (10 μg mL−1), and AF647-Dex (50 μg mL−1) for additional 1 h. After washing 3 times, cells were examined using CLSM with a 60× oil immersion objective (Ex/Em: 405/430–470 nm for CDs; Ex/Em: 635/650–700 nm for AF647). Then we used cellular organelles specific fluorescent markers, including LysoTracker Red, Golgi-RFP, ER-Tracker Red and MitoTracker Red, to determine intracellular distribution of CDs by assessing colocalization of CDs with lysosomes, Golgi apparatus, endoplasmic reticulum, and mitochondria, respectively. The process was performed in accordance with the manufacturer's instructions. We used CLSM with a 60× oil immersion objective to observe and excitation/emission wavelengths of CDs, MitoTracker Red, ER-Tracker Red, LysoTracker Red and Golgi-RFP were 405/430–470 nm, 559/580–625 nm, 559/600–630 nm, 559/575–640 nm and 559/565–620 nm, respectively. Last, transmission electron microscopy (TEM) was used to observe the localization of CDs in the cellular ultrastructure. PC12 and RSC96 cells (1 × 105 cells per well) were seeded in 6-well plates and grew overnight, then cells were incubated with CDs (0.2 mg mL−1) for 1 h. Subsequently, cells were washed 3 times with cold PBS, trypsinized, harvested, centrifuged, the supernatant was discarded and the pallet of fresh cells samples were fixed overnight with 2% glutaraldehyde solution. This was followed by secondary fixation with 1% osmium tetroxide for 2 h. Then the samples were dehydrated in a graded series of ethanol ending with propylene oxide. Samples were then embedded in Epon resin. About 70 nm thin sections were prepared with a PowerTome-XL ultramicrotome (RMC, USA). The sections were observed using a JEM-2100F high-resolution transmission electron microscope (JEOL, Japan) operating at 120 kV.3.8. Exocytosis of internalized CDs
After 3 h pre-incubation with CDs, the cells were washed by PBS for three times and cultured in fresh culture media without CDs. After further incubation (0.5, 1, 2, 4, 24, 48, 72 h), the cells were collected for FACS and CLSM analysis to measure the intracellular MFI. For low temperature treatment, the cells were cultured in fresh culture media at 4 °C for 1, 2, and 4 h after 3 h pre-incubation with CDs and measured by CLSM prior to three times PBS washings.4. Conclusions
CDs exhibit low cytotoxicity and highly efficient internalization in neural cells. The cellular uptake of CDs is dose, time, and partially energy-dependent and passive diffusion is involved in this process. CDs are endocytosed via caveolae-mediated and clathrin-mediated pathways. Internalized CDs locate mainly within endo-lysosomal structures, Golgi apparatus and partly enter nucleus. CDs can be actively transported to the cell periphery and exocytosed with a half-life of 2 h. Based on their excellent optical properties, good biocompatibility and comprehensive understanding of cellular behavior, it facilitates our development of bioimaging applications of CDs in nervous system.Acknowledgements
This work was supported by National Natural Science Foundation of China (81171145, 81371363, 51373065), and the Specialized Research Fund for the Doctoral Program of Higher Education (no. 20130061130010). We will like to thank Dr Yugang Ma for assistance in the preparation of TEM samples; Rui Sun and Xu Shi for assistance in the use of flow cytometry and confocal microscopy, respectively.Notes and references
- C. Ding, A. Zhu and Y. Tian, Acc. Chem. Res., 2014, 47, 20 CrossRef CAS PubMed.
- H. T. Li, Z. H. Kang, Y. Liu and S. T. Lee, J. Mater. Chem., 2012, 22, 24230 RSC.
- Y. Li, Y. Hu, Y. Zhao, G. Shi, L. Deng, Y. Hou and L. Qu, Adv. Mater., 2011, 23, 776 CrossRef CAS PubMed.
- L. Cao, M. J. Meziani, S. Sahu and Y. P. Sun, Acc. Chem. Res., 2013, 46, 171 CrossRef CAS PubMed.
- Y. Fang, S. Guo, D. Li, C. Zhu, W. Ren, S. Dong and E. Wang, ACS Nano, 2012, 6, 400 CrossRef CAS PubMed.
- M. J. Krysmann, A. Kelarakis, P. Dallas and E. P. Giannelis, J. Am. Chem. Soc., 2012, 134, 747 CrossRef CAS PubMed.
- R. Liu, D. Wu, X. Feng and K. Mullen, J. Am. Chem. Soc., 2011, 133, 15221 CrossRef CAS PubMed.
- S. T. Yang, L. Cao, P. G. Luo, F. Lu, X. Wang, H. Wang, M. J. Meziani, Y. Liu, G. Qi and Y. P. Sun, J. Am. Chem. Soc., 2009, 131, 11308 CrossRef CAS PubMed.
- H. Tao, K. Yang, Z. Ma, J. Wan, Y. Zhang, Z. Kang and Z. Liu, Small, 2012, 8, 281 CrossRef CAS PubMed.
- P. Huang, J. Lin, X. Wang, Z. Wang, C. Zhang, M. He, K. Wang, F. Chen, Z. Li, G. Shen, D. Cui and X. Chen, Adv. Mater., 2012, 24, 5104 CrossRef CAS PubMed.
- W. Shi, X. Li and H. Ma, Angew. Chem., Int. Ed., 2012, 51, 6432 CrossRef CAS PubMed.
- A. Zhu, Q. Qu, X. Shao, B. Kong and Y. Tian, Angew. Chem., Int. Ed., 2012, 51, 7185 CrossRef CAS PubMed.
- S. Karthik, B. Saha, S. K. Ghosh and N. D. Pradeep Singh, Chem. Commun., 2013, 49, 10471 RSC.
- J. Tang, B. Kong, H. Wu, M. Xu, Y. Wang, Y. Wang, D. Zhao and G. Zheng, Adv. Mater., 2013, 25, 6569 CrossRef CAS PubMed.
- L. Wang, X. Wang, A. Bhirde, J. Cao, Y. Zeng, X. Huang, Y. Sun, G. Liu and X. Chen, Adv. Healthcare Mater., 2014, 3, 1203 CrossRef CAS PubMed.
- L. Cao, X. Wang, M. J. Meziani, F. Lu, H. Wang, P. G. Luo, Y. Lin, B. A. Harruff, L. M. Veca, D. Murray, S. Y. Xie and Y. P. Sun, J. Am. Chem. Soc., 2007, 129, 11318 CrossRef CAS PubMed.
- S. C. Ray, A. Saha, N. R. Jana and R. Sarkar, J. Phys. Chem. C, 2009, 113, 18546 CAS.
- E. J. Goh, K. S. Kim, Y. R. Kim, H. S. Jung, S. Beack, W. H. Kong, G. Scarcelli, S. H. Yun and S. K. Hahn, Biomacromolecules, 2012, 13, 2554 CrossRef CAS PubMed.
- C. Liu, P. Zhang, X. Zhai, F. Tian, W. Li, J. Yang, Y. Liu, H. Wang, W. Wang and W. Liu, Biomaterials, 2012, 33, 3604 CrossRef CAS PubMed.
- S. N. Baker and G. A. Baker, Angew. Chem., Int. Ed., 2010, 49, 6726 CrossRef CAS PubMed.
- A. Verma and F. Stellacci, Small, 2010, 6, 12 CrossRef CAS PubMed.
- J. Wang, J. D. Byrne, M. E. Napier and J. M. DeSimone, Small, 2011, 7, 1919 CrossRef CAS PubMed.
- W. Jiang, B. Y. Kim, J. T. Rutka and W. C. Chan, Nat. Nanotechnol., 2008, 3, 145 CrossRef CAS PubMed.
- R. Agarwal, V. Singh, P. Jurney, L. Shi, S. V. Sreenivasan and K. Roy, Proc. Natl. Acad. Sci. U. S. A., 2013, 110, 17247 CrossRef CAS PubMed.
- A. Asati, S. Santra, C. Kaittanis and J. M. Perez, ACS Nano, 2010, 4, 5321 CrossRef CAS PubMed.
- K. Y. Win and S. S. Feng, Biomaterials, 2005, 26, 2713 CrossRef CAS PubMed.
- T. Xia, M. Kovochich, M. Liong, J. I. Zink and A. E. Nel, ACS Nano, 2008, 2, 85 CrossRef CAS PubMed.
- F. Zhao, Y. Zhao, Y. Liu, X. Chang, C. Chen and Y. Zhao, Small, 2011, 7, 1322 CrossRef CAS PubMed.
- S. Zhu, Q. Meng, L. Wang, J. Zhang, Y. Zhang, H. Jin, K. Zhang, H. Sun, H. Wang and B. Yang, Angew. Chem., Int. Ed., 2013, 52, 3953 CrossRef CAS PubMed.
- H. C. Tian, J. Q. Liu, D. X. Wei, X. Y. Kang, C. Zhang, J. C. Du, B. Yang, X. Chen, H. Y. Zhu, Y. N. Nuli and C. S. Yang, Biomaterials, 2014, 35, 2120 CrossRef CAS PubMed.
- S. N. Masand, I. J. Perron, M. Schachner and D. I. Shreiber, Biomaterials, 2012, 33, 790 CrossRef CAS PubMed.
- H. M. Chang, M. K. Shyu, G. F. Tseng, C. H. Liu, H. S. Chang, C. T. Lan, W. M. Hsu and W. C. Liao, PloS One, 2013, 8, e53444 CAS.
- Q. L. Zhao, Z. L. Zhang, B. H. Huang, J. Peng, M. Zhang and D. W. Pang, Chem. Commun., 2008, 5116 RSC.
- W. Shang, X. Zhang, M. Zhang, Z. Fan, Y. Sun, M. Han and L. Fan, Nanoscale, 2014, 6, 5799 RSC.
- K. K. Datta, O. Kozák, V. Ranc, M. Havrdová, A. B. Bourlinos, K. Safářová, K. Holá, K. Tománková, G. Zoppellaro, M. Otyepka and R. Zbořil, Chem. Commun., 2014, 50, 10782 RSC.
- E. Oh, J. B. Delehanty, K. E. Sapsford, K. Susumu, R. Goswami, J. B. Blanco-Canosa, P. E. Dawson, J. Granek, M. Shoff, Q. Zhang, P. L. Goering, A. Huston and I. L. Medintz, ACS Nano, 2011, 5, 6434 CrossRef CAS PubMed.
- L. Wang, S. J. Zhu, H. Y. Wang, S. N. Qu, Y. L. Zhang, J. H. Zhang, Q. D. Chen, H. L. Xu, W. Han, B. Yang and H. B. Sun, ACS Nano, 2014, 8, 2541 CrossRef CAS PubMed.
- Y. Song, S. Zhu, S. Xiang, X. Zhao, J. Zhang, H. Zhang, Y. Fu and B. Yang, Nanoscale, 2014, 6, 4676 RSC.
- Z. J. Zhu, T. Posati, D. F. Moyano, R. Tang, B. Yan, R. W. Vachet and V. M. Rotello, Small, 2012, 8, 2659 CrossRef CAS PubMed.
- A. Lesniak, F. Fenaroli, M. P. Monopoli, C. Aberg, K. A. Dawson and A. Salvati, ACS Nano, 2012, 6, 5845 CrossRef CAS PubMed.
- L. Treuel, S. Brandholt, P. Maffre, S. Wiegele, L. Shang and G. U. Nienhaus, ACS Nano, 2014, 8, 503 CrossRef CAS PubMed.
- P. Liu, Y. Sun, Q. Wang, Y. Sun, H. Li and Y. Duan, Biomaterials, 2014, 35, 760 CrossRef CAS PubMed.
- M. Ralser, M. M. Wamelink, E. A. Struys, C. Joppich, S. Krobitsch, C. Jakobs and H. Lehrach, Proc. Natl. Acad. Sci. U. S. A., 2008, 105, 17807 CrossRef PubMed.
- T. Lund, M. F. Callaghan, P. Williams, M. Turmaine, C. Bachmann, T. Rademacher, I. M. Roitt and R. Bayford, Biomaterials, 2011, 32, 9776 CrossRef CAS PubMed.
- W. Rima, L. Sancey, M. T. Aloy, E. Armandy, G. B. Alcantara, T. Epicier, A. Malchère, L. Joly-Pottuz, P. Mowat, F. Lux, O. Tillement, B. Burdin, A. Rivoire, C. Boulé, I. Anselme-Bertrand, J. Pourchez, M. Cottier, S. Roux, C. Rodriguez-Lafrasse and P. Perriat, Biomaterials, 2013, 34, 181 CrossRef CAS PubMed.
- C. Y. Hsu and H. Uludağ, Biomaterials, 2012, 33, 7834 CrossRef CAS PubMed.
- X. Jiang, C. Röcker, M. Hafner, S. Brandholt, R. M. Dörlich and G. U. Nienhaus, ACS Nano, 2010, 4, 6787 CrossRef CAS PubMed.
- W. Li, C. Chen, C. Ye, T. Wei, Y. Zhao, F. Lao, Z. Chen, H. Meng, Y. Gao, H. Yuan, G. Xing, F. Zhao, Z. Chai, X. Zhang, F. Yang, D. Han, X. Tang and Y. Zhang, Nanotechnology, 2008, 19, 145102 CrossRef PubMed.
- R. G. Parton and M. A. del Pozo, Nat. Rev. Mol. Cell Biol., 2013, 14, 98 CrossRef CAS PubMed.
- H. T. McMahon and E. Boucrot, Nat. Rev. Mol. Cell Biol., 2011, 12, 517 CrossRef CAS PubMed.
- G. Sahay, D. Y. Alakhova and A. V. Kabanov, J. Controlled Release, 2010, 145, 182 CrossRef CAS PubMed.
- Z. Wang, C. Tiruppathi, R. D. Minshall and A. B. Malik, ACS Nano, 2009, 3, 4110 CrossRef CAS PubMed.
- C. Wu, C. Wang, T. Han, X. Zhou, S. Guo and J. Zhang, Adv. Healthcare Mater., 2013, 2, 1613 CrossRef CAS PubMed.
- B. Kang, S. Chang, Y. Dai, D. Yu and D. Chen, Small, 2010, 6, 2362 CrossRef CAS PubMed.
- K. C. Partlow, G. M. Lanza and S. A. Wickline, Biomaterials, 2008, 29, 3367 CrossRef CAS PubMed.
- B. J. Nichols, Nat. Cell Biol., 2002, 4, 374 CAS.
- G. Sahay, J. O. Kim, A. V. Kabanov and T. K. Bronich, Biomaterials, 2010, 31, 923 CrossRef CAS PubMed.
- A. M. Schrand, J. J. Schlager, L. Dai and S. M. Hussain, Nat. Protoc., 2010, 5, 744 CrossRef CAS PubMed.
- N. Doshi and S. Mitragotri, Adv. Funct. Mater., 2009, 19, 3843 CrossRef CAS.
- R. Duncan and S. C. Richardson, Mol. Pharm., 2012, 9, 2380 CrossRef CAS PubMed.
- B. M. Prabhu, S. F. Ali, R. C. Murdock, S. M. Hussain and M. Srivatsan, Nanotoxicology, 2010, 4, 150 CrossRef CAS PubMed.
- A. M. Mohs and J. M. Provenzale, Neuroimaging. Clin. N. Am., 2010, 20, 283 CrossRef PubMed.
- R. M. Koffie, C. T. Farrar, L. J. Saidi, C. M. William, B. T. Hyman and T. L. Spires-Jones, Proc. Natl. Acad. Sci. U. S. A., 2011, 108, 18837 CrossRef CAS PubMed.
- C. W. Wu, O. Vasalatiy, N. Liu, H. Wu, S. Cheal, D. Y. Chen, A. P. Koretsky, G. L. Griffiths, R. B. Tootell and L. G. Ungerleider, Neuron, 2011, 70, 229 CrossRef CAS PubMed.
- Y. M. Bae, Y. I. Park, S. H. Nam, J. H. Kim, K. Lee, H. M. Kim, B. Yoo, J. S. Choi, K. T. Lee, T. Hyeon and Y. D. Suh, Biomaterials, 2012, 33, 9080 CrossRef CAS PubMed.
- G. J. Gordillo, Z. Krpetić and M. Brust, ACS Nano, 2014, 8, 6074 CrossRef CAS PubMed.
- P. Nativo, I. A. Prior and M. Brust, ACS Nano, 2008, 2, 1639 CrossRef CAS PubMed.
- Z. Krpetić, S. Saleemi, I. A. Prior, V. Sée, R. Qureshi and M. Brust, ACS Nano, 2011, 5, 5195 CrossRef PubMed.
- A. Salvati, A. S. Pitek, M. P. Monopoli, K. Prapainop, F. B. Bombelli, D. R. Hristov, P. M. Kelly, C. Åberg, E. Mahon and D. A. Dawson, Nat. Nanotechnol., 2013, 8, 137 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/c4ra09525a |
| This journal is © The Royal Society of Chemistry 2014 |