
 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a Creative Commons Attribution 3.0 Unported Licence
The synthesis and application of a diazirine-modified uridine analogue for investigating RNA–protein interactions †
Christine C.
Smith
,
Marcel
Hollenstein
* and
Christian J.
Leumann
*
Department of Chemistry and Biochemistry, University of Bern, Freiestrasse 3, 3012 Bern, Switzerland. E-mail: hollenstein@dcb.unibe.ch; leumann@dcb.unibe.ch; Fax: +41 31 631 3422; Tel: +41 31 631 4355
First published on 23rd September 2014
Abstract
The roles played by many ncRNAs remain largely unknown. Similarly, relatively little is known about the RNA binding proteins involved in processing ncRNA. Identification of new RNA/RNA binding protein (RBP) interactions may pave the way to gain a better understanding of the complex events occurring within cells during gene expression and ncRNA biogenesis. The development of chemical tools for the isolation of RBPs is of paramount importance. In this context, we report on the synthesis of the uridine phosphoramidite U Dz that bears a diazirine moiety on the nucleobase. RNA probes containing U Dz units were irradiated in the presence of single-stranded DNA binding protein (SSB), which is also known to bind ssRNAs, and shown to efficiently (15% yield) and selectively cross-link to the protein. The corresponding diazirine-modified uridine triphosphate U DzTP was synthesized and its capacity to act as a substrate for the T7 RNA polymerase was tested in transcription assays. U DzTP was accepted with a maximum yield of 38% for a 26mer RNA containing a single incorporation and 28% yield for triple consecutive incorporations. Thus, this uridine analogue represents a convenient biochemical tool for the identification of RNA binding proteins and unraveling the role and function played by ncRNAs.
1. Introduction
In recent years it has become clear that the roles played by non-coding RNAs (ncRNAs) are diverse and complex. 1–3 Examples of ncRNAs include not only the classical transfer RNA (tRNA) 4 and ribosomal RNA (rRNA) 5 involved in translation, but small interfering RNA (siRNA), 6 micro RNA (miRNA), 6,7 small nucleolar RNA (snoRNA), 8 small nuclear RNA (snRNA) 8 and numerous others that do not fit into a distinct category. Each of these classes of ncRNAs fulfils specific functions in all three domains of life ( i.e. Eukarya, Bacteria, and Archaea) during gene expression, early development, 9 and cell maintenance, with an ever-growing list of diseases 10–13 related to unusual ncRNA behavior hinting at a crucial involvement of these RNAs for correct cellular function. 14 Difficulties in identifying the function of these ncRNAs is often connected to complications in differentiating mRNAs from ncRNAs in bioinformatic approaches, 15 alongside problems generating enough material with sufficient separation to distinguish unique ncRNAs in experimental procedures. 16RNA pathways are extensively regulated by proteins called RNA binding proteins (RBPs) 17–19 and vice versa, 20 and together RNA and RBPs function as ribonucleoprotein (RNP) complexes, the best known of which are the spliceosome 21 and ribosome. 5 The inherent association of RNA with proteins has led to the development of experimental approaches for identifying ncRNA and RBP function based on the interactions between the two groups of biomacromolecules. Computational analysis, 22–24 X-ray crystallography 25–27 and cross-linking 28–34 methods of isolation have all been employed to investigate RNA/RBP interactions. One advantage of cross-linking is that no prior knowledge of an interaction between a protein and RNA is required. It may also be used to identify networks of RNA and proteins involved in forming RNP complexes. Cross-linking between oligonucleotides and proteins may be achieved by the addition of external cross-linking reagents, 35–37 or by inducing the cross-link photochemically with UV light. 38,39
The advantage of photochemical cross-linking is that the natural binding is preserved, which may be disrupted by the addition of external reagents. Unfortunately, the wavelengths of UV light required to induce cross-linking between nucleic acids and proteins (∼254 nm) are actually damaging to the molecules and thus have limited applicability in vivo. 40 The introduction of photoreactive groups into nucleic acids, allows for the modulation of the wavelength of UV light needed to achieve reasonable yields of cross-linking and minimize the potential damage caused to biomolecules. 41 Such modifications have included halides, 28,42 inclusion of sulfur atoms, 43–45 and the addition of more sophisticated moieties such as azides, 46,47 and benzophenones. 48 In this context, diazirines 49–52 have a particular appeal over their simpler counterparts, as they form carbenes upon irradiation with long wavelength UV light, 53,54 that then insert into adjacent covalent bonds. 55,56 Particularly, π-stabilized carbenes, 57,58 such as generated by exposure of 3-trifluoromethyl-3-phenyldiazirine 54,59 to 365 nm irradiation, have been used in a wide range of applications. 56,60 Examples include: cross-linking of DNA–protein interactions; 49–52,61,62 photoactive amino acids and peptides for substrate identification; 63–65 and diazirine modified sugars to capture glycoprotein binding events. 66 Further to this, a recent study of commonly used cross-linkers have identified the trifluoromethyldiazirine moiety superior to benzophenone and phenylazide derivatives, as it produces simple and accurate cross-linking profiles of amyloid nanostructures. 67
The use of diazirines to identify DNA binding protein interactions hints at the possible application of this chemistry to identify RNA binding proteins. Although 3-trifluoromethyl-3-phenyldiazirine has been used to cross-link RNA previously, 68 it involved coupling the photoactive moiety to a 2-thiocytidine residue already incorporated into a tRNA anticodon loop before cross-linking it to the ribosome. This strategy has shortcomings as it can lead to poor efficiencies of cross-linking, and the external addition requires accessibility to the modified base before cross-linking can be achieved, thus limiting its application. Moreover, modified RNA triphosphates (N*TPs) have been previously synthesized as photoactive substrates for polymerases; although the modifications have generally been made on the DNA level, rather than RNA. 69 Photo-labelling of viral proteins during transcription with 5-azidouridine triphosphate 70 has been described, but the azido functionality is incompatible with phosphoramidite chemistry, limiting its applicability. In addition, N*TPs have efficiently served in in vitro selection experiments for the generation of modified aptamers and ribozymes, 71 and this methodology could be exploited to detect previously unknown interactions between RNA and proteins and polymerases.
Here, we report on the synthesis of the RNA building block U Dz bearing a diazirine moiety on the nucleobase ( Fig. 1) with the intent of developing a new chemical tool for the investigation of the interaction of RNAs with RBPs.
 | ||
| Fig. 1 Chemical structures of the photoactive uridine analogues U Dz and U DzTP . | ||
2. Results and discussion
2.1 Design of the probe
We postulated that by incorporating the diazirine moiety within an RNA probe, through attachment at the C5-position of the uridine nucleobase, the photoactive RNA probe would be compatible with a wider range of applications. The choice of the location of the diazirine photoreactive group was dictated by the fact that appending modifications at the C5-position of pyrimidines minimizes structural perturbation of duplexes. 72 In addition, the photoreactive group was connected to the nucleobase via a pentynol linker arm that was designed so as to minimize undesired self-binding to the RNA probe. Therefore, a diazirine-containing uridine analogue ( U Dz) was designed to be incorporated into RNA probes using standard phosphoramidite chemistry ( Fig. 1). The DNA analogue dU Dz has been previously synthesized by Carell et al. and incorporated into DNA strands opposite a UV induced pyrimidine lesion and was able to selectively cross-link the DNA lesion repair enzyme DNA photolyase in the presence of other proteins. 48,622.2 Synthesis of the phosphoramidite U Dz and nucleoside triphosphate U DzTP
The synthesis of phosphoramidite U Dz followed the synthetic pathway developed for the corresponding DNA building block. 62 Briefly, diazirine 1 ( ref. 54, 73 and 74) was coupled to 4-pentyn-1-ol in the presence of NaH, before Sonogashira coupling to the protected nucleoside 3 (which was prepared via standard DMT protection of 5-iodouridine) to generate 4, a common precursor to both U Dz and U DzTP ( Scheme 1 and ESI †). Indeed, in order to generate phosphoramidite U Dz, 4 was selectively protected at the 2′-OH with TBDMS before phosphitylation under standard conditions to yield U Dz (56% over 2 steps; Scheme 2).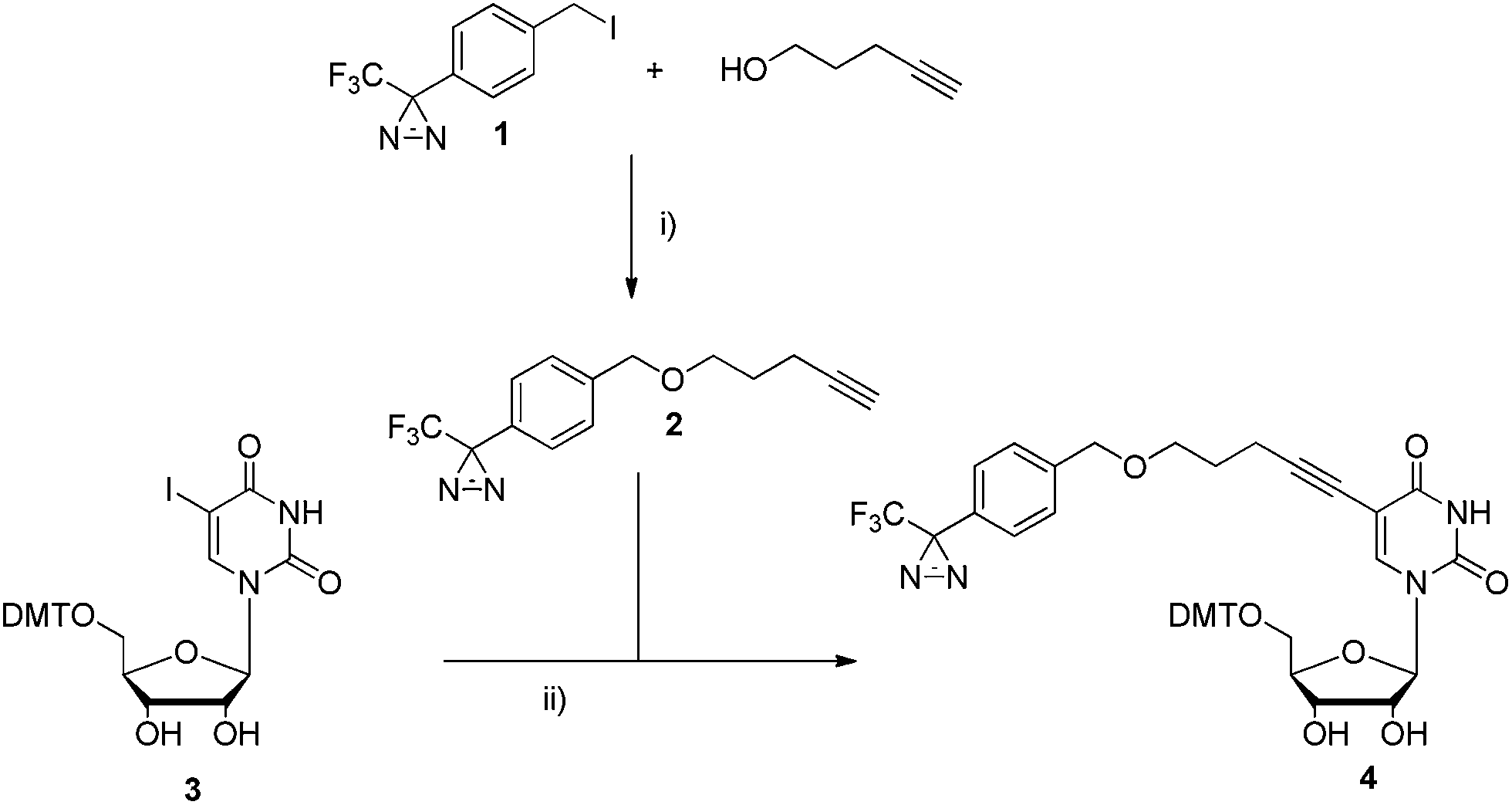 | ||
| Scheme 1 Synthesis of intermediate 4. Reagents and conditions were as follows: (i) 4-pentyn-1-ol, NaH, THF, 0 °C, 2 h, 68%; (ii) Pd(PPh 3) 2Cl 2, Et 3N, CuI, DMF, r.t., o.n., 84%. | ||
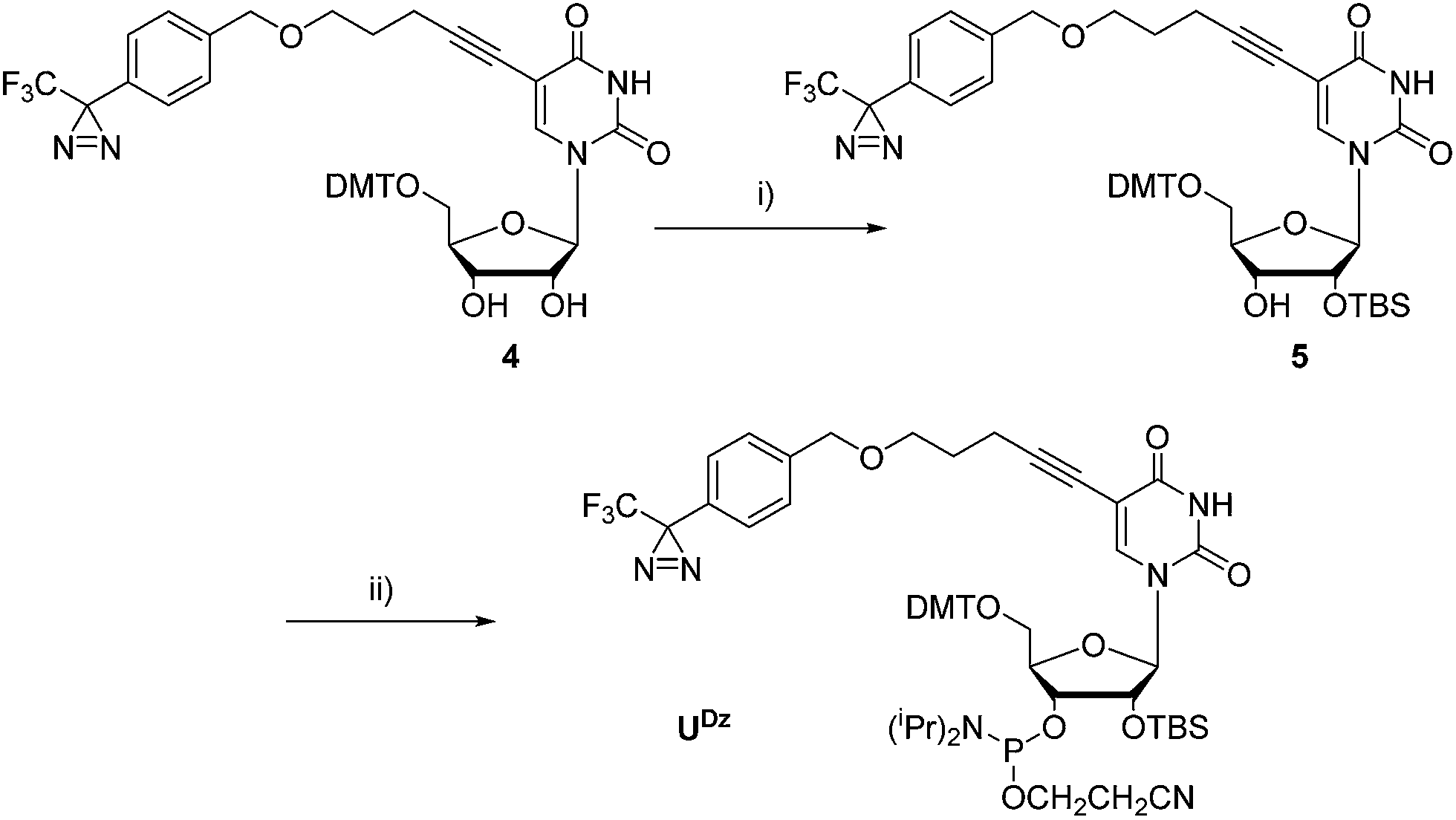 | ||
| Scheme 2 Synthesis of phosphoramidite U Dz. Reagents and conditions were as follows: (i) TBDMS-Cl, pyridine, AgNO 3, THF, r.t., o.n., 72%; (ii) CEP-Cl, DIPEA, THF, r.t., 1 h, 78%. | ||
Moreover, U DzTP could be accessed by first bis-acetylating compound 4 (71%) and then removing the DMT protecting group under acidic conditions ( Scheme 3). The suitably protected intermediate 7 was then converted to the corresponding triphosphate U DzTP by application of the Ludwig-Eckstein protocol 75,76 and was obtained in 21% after a thorough semi-preparative RP-HPLC purification. 76
 | ||
| Scheme 3 Synthesis of triphosphate U DzTP . Reagents and conditions were as follows: (i) Ac 2O, pyridine, DMAP, 0 °C to r.t., 2 h, 71%; (ii) 1% DCAA, CH 2Cl 2, 0 °C, 1.5 h, 48%; (iii) (1) 2-chloro-1,3,2-benzodioxaphosphorin-4-one, pyridine, dioxane; (2) ( nBu 3NH) 2H 2P 2O 7, nBu 3N, DMF; (3) I 2, pyridine, H 2O; (4) NH 3 (aq.), H 2O, 21% (over 4 steps). | ||
2.3 Oligonucleotide synthesis and thermal stability
The 5′-biotinylated 21mer RNA probes were designed for capture on streptavidin magnetic beads after cross-linking and contained single U Dz units at different locations within the sequence (ONs 2–4 in Table 1 and ESI †).| Sequence (5′ → 3′) | |
|---|---|
| ON1 | Biotin-CCUAUGUCACUGAAACUAUCC |
| ON2 | Biotin-CCUAUGUCACUGAAAC XAUCC |
| ON3 | Biotin-CCUAUGUCAC XGAAACUAUCC |
| ON4 | Biotin-CC XAUGUCACUGAAACUAUCC |
Moreover, the sequence composition was chosen so as to exclude the formation of dimers and tertiary structures. UV thermal melting measurements for each sequence versus both the RNA and DNA complements were recorded to assess whether the modification caused any disturbance in the duplex stability ( Fig. 2). The impact of the modification on the duplex stability with both the complementary DNA and RNA strands seems to be negligible (Δ Tm in the range of −1.4 to +0.5 °C per modification; Table 2). A similar behavior has been observed for other C5-modified uridine analogues, 77–79 and therefore modifications located at the C5 position of pyrimidines are well-accommodated in the major groove and do not interfere with duplex formation and stability. 80,81
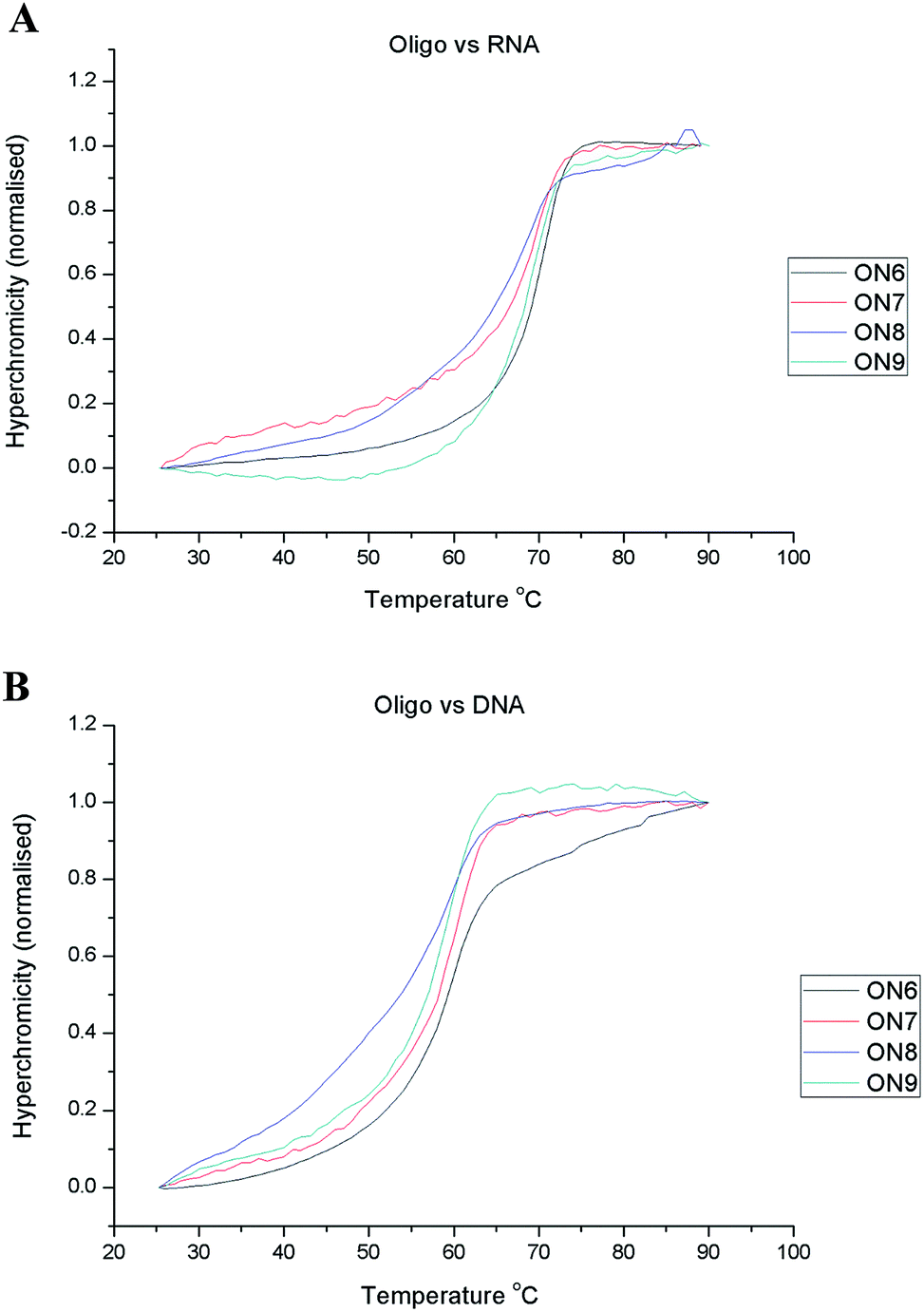 | ||
| Fig. 2 UV thermal melting experiments for ON1-4 versus RNA (A) and DNA (B). | ||
| T m vs. RNA (°C) (Δ Tm (°C) per mod.) | T m vs. DNA (°C) (Δ Tm (°C) per mod.) | |
|---|---|---|
| ON1 | 69.2 ± 1.5 | 59.6 ± 0.6 |
| ON2 | 69.6 ± 1.4 (+0.4) | 60.0 ± 0.5 (+0.4) |
| ON3 | 67.8 ± 1.5 (−1.4) | 60.1 ± 0.5 (+0.5) |
| ON4 | 69.1 ± 0.5 (−0.1) | 59.5 ± 0.6 (−0.1) |
2.4 Cross-linking experiments
Initial irradiation experiments with the modified oligonucleotides were performed in the presence of the single stranded DNA binding protein (SSB). 82 Although SSB is a DNA binding protein with a 10-fold lower affinity for ssRNA compared to ssDNA, 83,84 it was deemed sufficient for a “proof of concept” experiment. ON1–4 were 3′-end 32P-radiolabelled with T4 RNA ligase for visualization on the gel and to avoid photobleaching of potential fluorescent tags. 85The probes and negative controls were irradiated ( λ = 375 nm for 30 minutes, in pH 7.8 buffer) in the presence of a slight excess of the SSB protein, and eluted on a 10% tricine SDS PAGE, 86 followed by phosphorimaging ( Fig. 3). The irradiation conditions were chosen so that the diazirine will photolyze into reactive carbenes, which in turn can insert irreversibly into nearby CH bonds of the target protein. 48,62 The appearance of a slower running radioactive band (highlighted in red) in Lane 4 of each gel, is consistent with cross-linking of the modified RNA after irradiation. Indeed, the appearance of this band was not observed in the natural control (Lane 5) or in the absence of irradiation (Lane 2). Moreover, the appearance of a faint, faster running band in Lane 2 (highlighted in green), indicates that a higher molecular weight species than the free RNA is formed under irradiation conditions, in the absence of the protein.
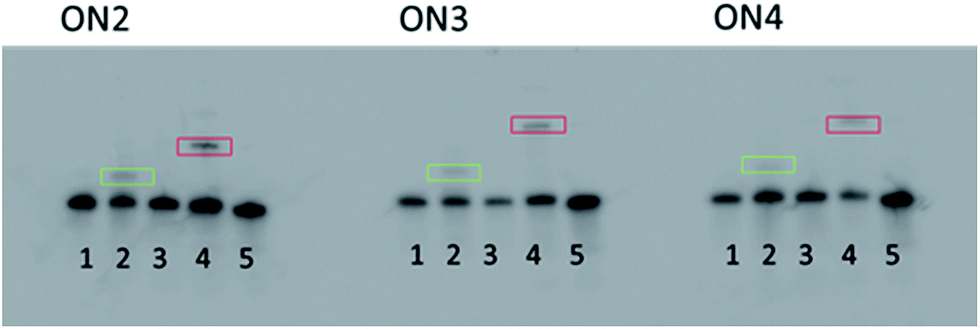 | ||
| Fig. 3 10% SDS PAGE gels of ON2–3 irradiated in the presence of SSB. Lane 1: ON. Lane 2: ON irradiated. Lane 3: ON + SSB, non-irradiated. Lane 4: ON + SSB, irradiated. Lane 5: ON1 + SSB, irradiated. | ||
We hypothesize that interstrand cross-linking might occur by insertion of the carbene generated under irradiation conditions into CH bonds of an adjacent RNA strand, although care was taken when designing the oligonucleotide sequences to ensure no hairpin formation or dimerization was likely. The disappearance of this band in the presence of the protein underscores the high selectivity and effectiveness of the diazirine photoreactive group incorporated in the RNA sequences.
As a second confirmation of the presence of RNA cross-linked to the protein, samples of ON1 and ON4 were irradiated in the presence of SSB, before purification with a size exclusion spin column (30 kDa cut off), then 3′-end 32P-radiolabelled. The resulting samples were eluted on 10% tricine SDS PAGE, alongside the ON4 sample prepared in the first experiment (see ESI Fig. S45 †). The radioactive band corresponding to the RNA–SSB crosslinks in both samples with ON4 eluted at the same rate, and the selective nature of radiolabelling using an RNA ligase confirms the presence of RNA in the modified samples, with no such corresponding band in the natural control, or in the absence of irradiation.
The yields for cross-linking to SSB obtained by quantification of the respective bands can be found in Table 3. The yield of cross-linking for each modified strand of RNA was consistently around 15%, which compares favorably to the crosslinking yields obtained for DNA probes equipped with diazirine groups and DNA photolyases. 62 The main side reaction competing with the carbene CH bond insertion in this experiment is the insertion of H 2O. The insertion of H 2O is unavoidable in the unbound RNA, thus, only the probes in contact with the protein at the moment of excitation are cross-linked. The advantage of the carbene being quenched by small molecules is that the resulting RNA will not interfere with data obtained for cross-linking to macromolecules as they will be resolved easily under electrophoretic conditions, giving specific cross-linking profiles for a mixture of components. Isolation with streptavidin beads was never achieved. Perhaps the bulk of the protein prevented the biotin from interacting with the beads.
| Percentage of cross-linked RNA (relative to free RNA) | ||
|---|---|---|
| Irradiated with SSB | Irradiated RNA | |
| ON2 | 14.0 ± 0.7 | 3.4 ± 2.5 |
| ON3 | 16.2 ± 2.2 | 5.1 ± 3.0 |
| ON4 | 16.3 ± 1.6 | 2.4 ± 1.8 |
2.5 Transcription assays
Nucleoside triphosphates represent a convenient vector for the introduction of chemical diversity into nucleic acids. 87–89 In the context of ribonucleoside triphosphates (N*TPs), the modifications are usually anchored at the 2′-OH of the sugar moiety and only rarely at the level of the nucleobase. 89 Thus, U DzTP was synthesized and used in transcription assays in order to assess the compatibility of this heavily-modified uridine analogue with an RNA polymerase. During transcription RNA polymerases use a double stranded DNA template to generate single stranded RNA transcripts. E. coli. T7 RNA polymerase is commonly used in molecular biology 90,91 and has been used to incorporate N*TPs including 2′-fluoro- 2′-amino-NTPs; 92 unnatural base pairs; 93,94 stabilized RNA analogues; 95 photoactive monomers such as 8-azido ATP, 96 5-bromo and 5-iodo UTP; 28,97 and other 5′-modified UTP analogues. 98,99 Matsuda et al. used a 43 base-pair dsDNA template containing a minimal T7 promoter followed by the sequence of the transcripts to generate 26 mer products containing 4′-thioRNA nucleotides. 95 This dsDNA template was the basis for the design of the system used to test the incorporation of U DzTP into RNA oligonucleotides ( Fig. 4). Indeed, the templates generate 26mer RNA transcripts containing a single (Product P1) or triple consecutive (Product P2) incorporation of U Dz nucleotides. Moreover, in order to assess the efficiency of the incorporation of the modified U DzTP in lieu of its natural counterpart, the transcription efficiencies were measured relative to that using all natural NTPs which was defined as being 100%. | ||
| Fig. 4 Incorporation of U DzTP into RNA via transcription assays using T7 RNA polymerase and dsDNA templates T1/ T1′ and T2/ T2′. Italicized letters represent the T7 promoter sequence. | ||
When the standard conditions for T7 RNA polymerase-mediated transcription reactions were used, little full length products could be observed when U DzTP was included (data not shown). On the other hand, optimized reaction conditions, i.e. increased concentrations of U DzTP and longer reaction times, ( vide infra), led to the appearance of bands corresponding to full length products when U DzTP was used as a substitute for UTP (Lane 3 in Fig. 5A and B). This confirms the ability of T7 RNA polymerase to accept this heavily modified N*TP as a substrate. Moreover, the absence in Lane 3 of similar abortive sequences as shown in the negative control experiments (Lanes 2 and 4) is in agreement with this statement and shows that the polymerization reactions proceed with high yields. Interestingly, the T7 RNA polymerase could readily incorporate three modified U DzTP in a row ( Fig. 5B) without stalling, clearly showing that this N*TP is a rather good substrate for the polymerase.
 | ||
| Fig. 5 Gel images of 20% PAGE of transcription assays using (A) dsDNA template T1/ T1′ or (B) template T2/ T2′. Lane 1: Natural NTPs. Lane 2: Negative control. Lane 3: U DzTP . Lane 4: ATP and GTP only. General procedure: 40 pmol of template DNA, 0.4 mM of natural NTPs, 0.6 mM U DzTP , 100 units of T7 RNA polymerase, 0.6 units of PPiase, 0.5 μl RNasin, and 10 μCi of α- 32P-ATP in the transcription optimized buffer. Reactions were incubated at 37 °C: template 1, for at least 8 hours; template 2 for at least 24 hours; under 20 μl mineral oil. | ||
However, the transcription efficiencies are rather depleted when compared to those obtained with the unmodified UTP ( Table 4) hinting at a slowing of transcription due to the presence of the N*TP. This is confirmed by the fact that the reaction times had to be extended (to at least 8 and 24 hours for the dsDNA templates T1/ T1′ and T2/ T2′, respectively) during optimization to allow the synthesis of the modified RNA. The polymerase was unable to generate the full length transcript with the exact template used by Matsuda et al. which should lead to six incorporations of U DzTP (data not shown). There also seems to be a position dependence on the ability of the enzyme to incorporate U DzTP , since using a template containing complementary A nucleotides at positions 10, 15 and 21 (template T3/ T3′, Table S2, ESI †) only led to abortive sequences (data not shown).
| Product P1 | Product P2 | |
|---|---|---|
| Efficiency compared to natural transcript (%) | 38 | 28 |
However, a longer template T4/ T4′ designed to generate a 52mer RNA product (consisting the sequences of templates T1/ T1′ and T3/ T3′ appended onto one another Table S2, ESI †) containing 4 modifications dispersed throughout the transcript did generate full length transcripts, in a similar yield (37%) and time scale (26 hours) as the other templates (Fig. S46, ESI †).
3. Conclusions
With the intent of developing a new tool for the investigation of RNA–protein interactions, a photoreactive diazirine-containing uridine analogue was designed so that it could be incorporated into RNA, either synthetically via phosphoramidite chemistry ( U Dz) or by enzymatic polymerization ( U DzTP ).The modified RNA oligonucleotides obtained by incorporation of the phosphoramidite building block during solid-phase synthesis served two purposes: (1) assessing the effect of the modification on duplex stability and (2) showing that the inclusion of the photoreactive diazirine moiety into RNAs could serve as a versatile tool for the investigation of RNA–protein interactions. Not surprisingly, UV-melting experiments revealed that the modification did not cause any disruption of the Watson–Crick base-pairing and was well-tolerated in the major groove of the duplexes. Moreover, the resulting synthetic probes were assessed for cross-linking potential to single-stranded binding protein (SSB) in an irradiation assay. The RNA probes containing the modified uridine analogue were selectively cross-linked to SSB. The selectivity of this assay derives from the main competitor of the CH bond insertion of the protein, being the insertion of H 2O, thus only molecules in direct contact with the modified RNA will be cross-linked, preventing non-specific cross-linking. Even though the SSB mainly served as a paradigm for the development of photo-crosslinking RNA probes, this approach can be extended to investigate on the binding mode of other single-stranded nucleic acid binding proteins. Indeed, the virus-encoded single-stranded DNA binding protein ICP8 was shown to bind to ssRNA with an affinity only ∼5 fold lower than that of ssDNA, hinting at a possible implication of ICP8 in the regulation of viral replication. 84 Finally, other yet unknown proteins binding to various forms of coding or non-coding RNAs could be easily targeted by application of this general method.
The ability of T7 RNA polymerase to accept the triphosphate analogue was also investigated in transcription assays, generating modified RNA containing up to four incorporations of the diazirine-modified uridine. The efficient transcription reactions with U DzTP show that bulkier modifications are also tolerated at this particular location. In this context, various investigations have shown that UTP analogues equipped with smaller modifications such as hydrophobic moieties, 98 amines, thiols, 79,100 and azides 101 acted as good substrates for the T7 RNA polymerase. Thus, the T7 RNA polymerase is rather tolerant to a broad variety of modifications anchored at position C5 of pyrimidine nucleoside triphosphates.
However, the rate of transcription seems to be retarded by the presence of the modified UTP. A kinetic investigation on the effect of the analogue U DzTP on the catalytic efficiency ( kcat/ KM) is currently under way and will shed some light on this phenomenon. Finally, the substrate acceptance of the noncanonical U DzTP analogue might be further improved by using mutant RNA polymerases. 102
Overall, the good substrate acceptance of triphosphate U DzTP and the photo-crosslinking ability of the probes containing the uridine analogue U Dz bode well for the isolation and identification of RNA binding proteins and ncRNAs as well as understanding their interactions.
4. Experimental section
4.1 Cross-linking experiments
Single-stranded DNA binding protein (SSB) (1 μg, 10 pmol) and oligonucleotide (3 pmol of radiolabelled RNA) were incubated in the irradiation buffer (50 mM Tris-HCl, 10 mM MgCl 2, 10 mM DTT, pH 7.8) in a total volume of 15 μl in a PCR vial at room temperature for at least 30 minutes prior to irradiation. The sample was irradiated at 375 nm for 30 minutes. Loading buffer (70% formamide, 50 mM EDTA, bromophenol blue and xylene cyanol FF (each 0.1%) 15 μl) was then added and heated to 95 °C for 5 minutes. The resulting RNA–protein complexes were then resolved on 10% SDS-PAGE at 10 W for 2 hours. The gel was exposed on an imaging plate at −18 °C overnight before visualizing on a phosphorimager.4.2 Transcription assays
General procedure: 40 pmol of ds template DNA were annealed by heating to 95 °C and then gradually cooling down to room temperature. 0.4 mM of natural NTPs, 0.6 mM U DzTP , 100 units of T7 RNA polymerase, 0.6 units of PPiase, 0.5 μl RNasin, and 10 μCi of α- 32P-ATP were then added at 0 °C to the transcription optimized buffer composed of 40 mM Tris–HCl (pH 7.9), 6 mM MgCl 2, 10 mM NaCl, 2 mM spermidine, 10 mM DTT and 0.05% of the polysorbate detergent Tween, and made up to a total volume of 20 μl with ultra-pure H 2O. Reactions were incubated at 37 °C for different reaction times: template T1, 8 hours; template T2 24 hours; under 20 μl mineral oil. The reactions were quenched by adding of 20 μL of stop solution (70% formamide, 50 mM EDTA, 0.1% bromophenol, 0.1% xylene cyanol) and resolved on 15–20% PAGE.5. Abbreviations
| U Dz | Diazirine modified uridine phosphosphoramidite |
| U DzTP | Diazirine modified uridine triphosphate; |
| N*TP | Modified nucleoside triphosphate |
| Tris | Tris(hydroxymethyl)aminomethane |
| TBDMS | tert-Butyldimethylsilyl |
| RP-HPLC | Reversed phase high-performance liquid chromatography |
| T m | Melting temperature |
| ON | Oligonucleotide |
| T | Template |
| P | Product |
| DMF | Dimethyl formamide |
| nBu 3N | n-Tributylamine |
| DCAA | Dichloroacetic acid |
| DIPEA | N, N-Diisopropylethylamine |
| CEP-Cl | 2′-Cyanoethyl N, N-diisopropylchlorophosphoramidite |
| r.t. | Room temperature |
| o.n. | Overnight |
| DMAP | 4-Dimethylaminopyridine |
Acknowledgements
This work was supported by grants from the Swiss National Science Foundation (grant-no.: PZ00P2_144595 to M.H. and grant-no.: 200020-13037 to C.J.L.).Notes and references
- M. Morlando, M. Ballarino, A. Fatica and I. Bozzoni, ChemMedChem, 2014, 9, 505–510 CrossRef CAS PubMed.
- J. S. Mattick and I. V. Makunin, Hum. Mol. Genet., 2006, 15, R17–R29 CrossRef CAS PubMed.
- A. Huttenhofer, P. Schattner and N. Polacek, Trends Genet., 2005, 21, 289–297 CrossRef PubMed.
- A. Rich and U. L. Rajbhandary, Annu. Rev. Biochem., 1976, 45, 805–860 CrossRef CAS PubMed.
- R. N. Nazar, IUBMB Life, 2004, 56, 457–465 CrossRef CAS PubMed.
- R. W. Carthew and E. J. Sontheimer, Cell, 2009, 136, 642–655 CrossRef CAS PubMed.
- D. P. Bartel, Cell, 2004, 116, 281–297 CrossRef CAS.
- A. G. Matera, R. M. Terns and M. P. Terns, Nat. Rev. Mol. Cell Biol., 2007, 8, 209–220 CrossRef CAS PubMed.
- U. A. Orom, T. Derrien, M. Beringer, K. Gumireddy, A. Gardini, G. Bussotti, F. Lai, M. Zytnicki, C. Notredame, Q. Huang, R. Guigo and R. Shiekhattar, Cell, 2010, 143, 46–58 CrossRef CAS PubMed.
- M. Esteller, Nat. Rev. Genet., 2011, 12, 861–874 CrossRef CAS PubMed.
- M. A. Varela, T. C. Roberts and M. J. A. Wood, Neurotherapeutics, 2013, 10, 621–631 CrossRef CAS PubMed.
- S. D. Rege, T. Geetha, S. R. Pondugula, C. A. Zizza, C. M. Wernette and J. R. Babu, ISRN Neurol., 2013, 2013, 375852 Search PubMed.
- T. A. Cooper, L. L. Wan and G. Dreyfuss, Cell, 2009, 136, 777–793 CrossRef CAS PubMed.
- J. E. Wilusz, H. Sunwoo and D. L. Spector, Genes Dev., 2009, 23, 1494–1504 CrossRef CAS PubMed.
- M. E. Dinger, K. C. Pang, T. R. Mercer and J. S. Mattick, PLoS Comput. Biol., 2008, 4, e1000176 Search PubMed.
- A. Huttenhofer and J. Vogel, Nucleic Acids Res., 2006, 34, 635–646 CrossRef PubMed.
- D. J. Hogan, D. P. Riordan, A. P. Gerber, D. Herschlag and P. O. Brown, PLoS Biol., 2008, 6, 2297–2313 CAS.
- T. Glisovic, J. L. Bachorik, J. Yong and G. Dreyfuss, FEBS Lett., 2008, 582, 1977–1986 CrossRef CAS PubMed.
- A. E. Pasquinelli, Nat. Rev. Genet., 2012, 13, 271–282 CAS.
- X. Wang, S. Arai, X. Song, D. Reichart, K. Du, G. Pascual, P. Tempst, M. G. Rosenfeld, C. K. Glass and R. Kurokawa, Nature, 2008, 454, 126–130 CrossRef CAS PubMed.
- Z. L. Zhou, L. J. Licklider, S. P. Gygi and R. Reed, Nature, 2002, 419, 182–185 CrossRef CAS PubMed.
- S. Jones, D. T. A. Daley, N. M. Luscombe, H. M. Berman and J. M. Thornton, Nucleic Acids Res., 2001, 29, 943–954 CrossRef CAS PubMed.
- M. Terribilini, J.-H. Lee, C. Yan, R. L. Jernigan, V. Honavar and D. Dobbs, RNA, 2006, 12, 1450–1462 CrossRef CAS PubMed.
- H. Kim, E. Jeong, S. W. Lee and K. Han, FEBS Lett., 2003, 552, 231–239 CrossRef CAS.
- T. Ritschel, C. Atmanene, K. Reuter, A. Van Dorsselaer, S. Sanglier-Cianferani and G. Klebe, J. Mol. Biol., 2009, 393, 833–847 CrossRef CAS PubMed.
- J. A. Doudna, Curr. Biol., 1999, 9, R731–R734 CrossRef CAS.
- C. Oubridge, N. Ito, P. R. Evans, C. H. Teo and K. Nagai, Nature, 1994, 372, 432–438 CrossRef CAS PubMed.
- M. C. Willis, B. J. Hicke, O. C. Uhlenbeck, T. R. Cech and T. H. Koch, Science, 1993, 262, 1255–1257 CAS.
- J. Ule, K. B. Jensen, M. Ruggiu, A. Mele, A. Ule and R. B. Darnell, Science, 2003, 302, 1212–1215 CrossRef CAS PubMed.
- D. D. Licatalosi, A. Mele, J. J. Fak, J. Ule, M. Kayikci, S. W. Chi, T. A. Clark, A. C. Schweitzer, J. E. Blume, X. Wang, J. C. Darnell and R. B. Darnell, Nature, 2008, 456, 464–469 CrossRef CAS PubMed.
- J. L. Corden, Chem. Biol., 2010, 17, 316–318 CrossRef CAS PubMed.
- M. Hafner, M. Landthaler, L. Burger, M. Khorshid, J. Hausser, P. Berninger, A. Rothballer, M. Ascano Jr, A. C. Jungkamp, M. Munschauer, A. Ulrich, G. S. Wardle, S. Dewell, M. Zavolan and T. Tuschl, Cell, 2010, 141, 129–141 CrossRef CAS PubMed.
- J. Koenig, K. Zarnack, G. Rot, T. Curk, M. Kayikci, B. Zupan, D. J. Turner, N. M. Luscombe and J. Ule, Nat. Struct. Mol. Biol., 2010, 17, 909–915 CAS.
- M. J. Luo and R. Reed, et al., Current protocols in molecular biology, ed. Frederick M. Ausubel, 2003, ch. 27, Unit 27 22 Search PubMed.
- I. Wower, J. Wower, M. Meinke and R. Brimacombe, Nucleic Acids Res., 1981, 9, 4285–4302 CrossRef CAS PubMed.
- M. Osswald, B. Greuer and R. Brimacombe, Nucleic Acids Res., 1990, 18, 6755–6760 CrossRef CAS PubMed.
- L. L. G. Carrette, E. Gyssels, J. Loncke and A. Madder, Org. Biomol. Chem., 2014, 12, 931–935 CAS.
- R. Pelle and N. B. Murphy, Nucleic Acids Res., 1993, 21, 2453–2458 CrossRef CAS PubMed.
- B. M. Merrill, K. R. Williams, J. W. Chase and W. H. Konigsberg, J. Biol. Chem., 1984, 259, 10850–10856 CAS.
- G. J. Kantor and D. R. Hull, Biophys. J., 1979, 27, 359–370 CrossRef CAS.
- M. M. Konarska, Methods, 1999, 18, 22–28 CrossRef CAS PubMed.
- M. C. Willis, K. A. Lecuyer, K. M. Meisenheimer, O. C. Uhlenbeck and T. H. Koch, Nucleic Acids Res., 1994, 22, 4947–4952 CrossRef CAS PubMed.
- A. Favre, G. Moreno, M. O. Blondel, J. Kliber, F. Vinzens and C. Salet, Biochem. Biophys. Res. Commun., 1986, 141, 847–854 CrossRef CAS.
- E. J. Sontheimer, Mol. Biol. Rep., 1994, 20, 35–44 CrossRef CAS.
- D. Temiakov, P. E. Mentesana, K. Ma, A. Mustaev, S. Borukhov and W. T. McAllister, Proc. Natl. Acad. Sci. U. S. A., 2000, 97, 14109–14114 CrossRef CAS PubMed.
- R. K. Evans, J. D. Johnson and B. E. Haley, Proc. Natl. Acad. Sci. U. S. A., 1986, 83, 5382–5386 CrossRef CAS.
- N. Demeshkina, M. Repkova, A. Ven'yaminova, D. Graifer and G. Karpova, RNA, 2000, 6, 1727–1736 CrossRef CAS.
- M. Winnacker, V. Welzmiller, R. Strasser and T. Carell, ChemBioChem, 2010, 11, 1345–1349 CrossRef CAS PubMed.
- T. Yamaguchi and M. Saneyoshi, Nucleosides Nucleotides, 1996, 15, 607–618 CAS.
- T. Yamaguchi, K. Suyama, K. Narita, S. Kohgo, A. Tomikawa and M. Saneyoshi, Nucleic Acids Res., 1997, 25, 2352–2358 CrossRef CAS PubMed.
- M. Liebmann, F. Di Pasquale and A. Marx, ChemBioChem, 2006, 7, 1965–1969 CrossRef CAS PubMed.
- U. K. Shigdel, J. Zhang and C. He, Angew. Chem., Int. Ed. Engl., 2008, 47, 90–93 CrossRef CAS PubMed.
- G. F. Bradley, W. B. L. Evans and I. D. R. Stevens, J. Chem. Soc., Perkin Trans. 2, 1977, 1214–1220 RSC.
- J. Brunner, H. Senn and F. M. Richards, J. Biol. Chem., 1980, 255, 3313–3318 CAS.
- R. A. Moss, Acc. Chem. Res., 2006, 39, 267–272 CrossRef CAS PubMed.
- J. Das, Chem. Rev., 2011, 111, 4405–4417 CrossRef CAS PubMed.
- P. H. Mueller, N. G. Rondan, K. N. Houk, J. F. Harrison, D. Hooper, B. H. Willen and J. F. Liebman, J. Am. Chem. Soc., 1981, 103, 5049–5052 CrossRef CAS.
- D. L. S. Brahms and W. P. Dailey, Chem. Rev., 1996, 96, 1585–1632 CrossRef CAS PubMed.
- M. Nassal, Liebigs Ann. Chem., 1983, 1983, 1510–1523 CrossRef.
- L. Dubinsky, B. P. Krom and M. M. Meijler, Bioorg. Med. Chem., 2012, 20, 554–570 CrossRef CAS PubMed.
- T. Yamaguchi and M. Saneyoshi, Nucleic Acids Res., 1996, 24, 3364–3369 CrossRef CAS PubMed.
- M. Winnacker, S. Breeger, R. Strasser and T. Carell, ChemBioChem, 2009, 10, 109–118 CrossRef CAS PubMed.
- Y. Ambroise, F. Pillon, C. Mioskowski, A. Valleix and B. Rousseau, Eur. J. Org. Chem., 2001, 3961–3964 CrossRef CAS.
- H. Nakashima, M. Hashimoto, Y. Sadakane, T. Tomohiro and Y. Hatanaka, J. Am. Chem. Soc., 2006, 128, 15092–15093 CrossRef CAS PubMed.
- D. P. Smith, J. Anderson, J. Plante, A. E. Ashcroft, S. E. Radford, A. J. Wilson and M. J. Parker, Chem. Commun., 2008, 5728–5730 RSC.
- M. R. Bond, H. Zhang, P. D. Vu and J. J. Kohler, Nat. Protoc., 2009, 4, 1044–1063 CrossRef CAS PubMed.
- G. W. Preston, S. E. Radford, A. E. Ashcroft and A. J. Wilson, ACS Chem. Biol., 2014, 9, 761–768 CrossRef CAS PubMed.
- T. Doring, P. Mitchell, M. Osswald, D. Bochkariov and R. Brimacombe, EMBO J., 1994, 13, 2677–2685 CAS.
- S. Verma and F. Eckstein, Annu. Rev. Biochem., 1998, 67, 99–134 CrossRef CAS PubMed.
- D. C. Hammond, R. K. Evans and J. A. Lesnaw, J. Gen. Virol., 1992, 73, 61–66 CrossRef CAS PubMed.
- M. Kuwahara and N. Sugimoto, Molecules, 2010, 15, 5423–5444 CrossRef CAS PubMed.
- E. T. Kool, Curr. Opin. Chem. Biol., 2000, 4, 602–608 CrossRef CAS.
- L. B. Shih and H. Bayley, Anal. Biochem., 1985, 144, 132–141 CrossRef CAS.
- J. H. Robson and J. Reinhart, J. Am. Chem. Soc., 1955, 77, 498–499 CrossRef CAS.
- J. Ludwig and F. Eckstein, J. Org. Chem., 1989, 54, 631–635 CrossRef CAS.
- M. Hollenstein, C. C. Smith and M. Raez, J. Visualized Exp., 2014, e51385 Search PubMed.
- M. Terrazas and E. T. Kool, Nucleic Acids Res., 2009, 37, 346–353 CrossRef CAS PubMed.
- M. Terrazas and R. Eritja, Mol. Diversity, 2011, 15, 677–686 CrossRef CAS PubMed.
- T. Schoetzau, J. Langner, E. Moyroud, I. Roehl, S. Vonhoff and S. Klussmann, Bioconjugate Chem., 2003, 14, 919–926 CrossRef CAS PubMed.
- P. Čapek, H. Cahová, R. Pohl, M. Hocek, C. Gloeckner and A. Marx, Chem.–Eur. J., 2007, 13, 6196–6203 CrossRef PubMed.
- M. Hollenstein, Chem.–Eur. J., 2012, 18, 13320–13330 CrossRef CAS PubMed.
- A. Sancar, K. R. Williams, J. W. Chase and W. D. Rupp, Proc. Natl. Acad. Sci. U. S. A., 1981, 78, 4274–4278 CrossRef CAS.
- J. W. Chase and K. R. Williams, Annu. Rev. Biochem., 1986, 55, 103–136 CrossRef CAS PubMed.
- P. E. Boehmer, Nucleic Acids Res., 2004, 32, 4576–4584 CrossRef CAS PubMed.
- C. I. Richards, S. Choi, J.-C. Hsiang, Y. Antoku, T. Vosch, A. Bongiorno, Y.-L. Tzeng and R. M. Dickson, J. Am. Chem. Soc., 2008, 130, 5038–5039 CrossRef CAS PubMed.
- H. Schaegger, Nat. Protoc., 2006, 1, 16–22 CrossRef CAS PubMed.
- M. Hocek and M. Fojta, Org. Biomol. Chem., 2008, 6, 2233–2241 CAS.
- M. Hollenstein, Molecules, 2012, 17, 13569–13591 CrossRef CAS PubMed.
- L. H. Lauridsen, J. A. Rothnagel and R. N. Veedu, ChemBioChem, 2012, 13, 19–25 CrossRef PubMed.
- J. F. Milligan, D. R. Groebe, G. W. Witherell and O. C. Uhlenbeck, Nucleic Acids Res., 1987, 15, 8783–8798 CrossRef CAS PubMed.
- K. E. McGinness and G. F. Joyce, J. Biol. Chem., 2001, 277, 2987–2991 CrossRef PubMed.
- H. Aurup, D. M. Williams and F. Eckstein, Biochemistry, 1992, 31, 9636–9641 CrossRef CAS.
- M. Kimoto, M. Endo, T. Mitsui, T. Okuni, I. Hirao and S. Yokoyama, Chem. Biol., 2004, 11, 47–55 CrossRef CAS PubMed.
- R. Kawai, M. Kimoto, S. Ikeda, T. Mitsui, M. Endo, S. Yokoyama and I. Hirao, J. Am. Chem. Soc., 2005, 127, 17286–17295 CrossRef CAS PubMed.
- Y. Kato, N. Minakawa, Y. Komatsu, H. Kamiya, N. Ogawa, H. Harashima and A. Matsuda, Nucleic Acids Res., 2005, 33, 2942–2951 CrossRef CAS PubMed.
- S. Gopalakrishna, V. Gusti, S. Nair, S. Sahar and R. K. Gaur, RNA, 2004, 10, 1820–1830 CrossRef CAS PubMed.
- J. M. Gott, M. C. Willis, T. H. Koch and O. C. Uhlenbeck, Biochemistry, 1991, 30, 6290–6295 CrossRef CAS.
- J. D. Vaught, T. Dewey and B. E. Eaton, J. Am. Chem. Soc., 2004, 126, 11231–11237 CrossRef CAS PubMed.
- A. A. Tanpure and S. G. Srivatsan, ChemBioChem, 2014, 15, 1309–1316 CrossRef CAS PubMed.
- N. K. Vaish, A. W. Fraley, J. W. Szostak and L. W. McLaughlin, Nucleic Acids Res., 2000, 28, 3316–3322 CrossRef CAS PubMed.
- H. Rao, A. A. Tanpure, A. A. Sawant and S. G. Srivatsan, Nat. Protoc., 2012, 7, 1097–1112 CrossRef CAS PubMed.
- J. Attwater, S. Tagami, M. Kimoto, K. Butler, E. T. Kool, J. Wengel, P. Herdewijn, I. Hirao and P. Holliger, Chem. Sci., 2013, 4, 2804–2814 RSC.
Footnote |
| † Electronic supplementary information (ESI) available: Experimental details of the synthesis of U Dz and U DzTP , cross-linking experiments, transcription assays, and additional gels. See DOI: 10.1039/c4ra08682a |
| This journal is © The Royal Society of Chemistry 2014 |