
Comparison of different probes based on labeled annexin V for detection of apoptosis
Abstract
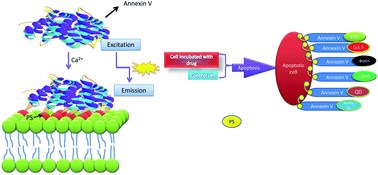
Apoptosis is a well-organized mechanism developed by eukaryotic organisms during maturation. The importance of apoptosis as a major form of cellular suicide with a causative or contributing role in a variety of diseases has become progressively obvious. A large body of evidence implicates the association of apoptosis with an arranged series of physical and biochemical alterations comprising the nucleus, cytoplasm and cell membrane. Understanding the process of apoptosis is not only important for maintaining physiological conditions, but also is critical for therapy development. In this review, we briefly outline the various ways to detect apoptosis with distinct assays and molecules, and provide an outlook on the latest imaging techniques for the visualization of phosphatidylserine (PS) externalization, with specific focus on annexin V based targeting of PS and its conjugates. Furthermore, the distinct advantages and disadvantages of each technique are reviewed. Recognition of the major benefits and drawbacks of each assay based on annexin V conjugates would allow us to use the appropriate method to detect apoptosis for instance, in a variety of diseases, such as cancer and will open novel prospects for treatment approaches.
 Please wait while we load your content...
Please wait while we load your content...

