
Steam explosion-ionic liquid pretreatments on wetland lignocellulosic biomasses of Phragmites (sp.) and Thalia dealbata for Bio H2 conversion
Hongyun Peng*a,
Lingling Gaoa,
Mengjiao Lia,
Yibin Shena,
Qiongqiu Qianb and
Xia Lia
aKey Laboratory of Environmental Remediation and Ecological Health, Ministry of Education, College of Environmental & Resource Sciences, Zhejiang University, Zijingang Campus, No. 866 Yuhangtang Rd, Hangzhou 310058, China. E-mail: penghongyun@zju.edu.cn; Fax: +86 571 86435070; Tel: +86 571 86435070
bCollege of Agriculture & Biotechnology, Zhejiang University, Zijingang Campus, No. 866 Yuhangtang Rd, Hangzhou 310058, China
First published on 22nd July 2014
Abstract
Bio H2 conversion from wetland lignocellulosic biomass is one of the promising alternatives to fossil fuels. Both Phragmites (sp.) and Thalia dealbata are holocellulose-rich and lignin-rich wetland plant biomasses. Scanning electron microscopy (SEM) and Fourier transform infrared spectroscopy (FTIR) demonstrated alteration of lignocellulosic structures due to hemicelluloses removal by steam explosion (SE) pretreatment, and further disruption of cellulose crystallinity after treatment with SE followed by [Bmim]Cl (SE-[Bmim]Cl). Thermogravimetry/differential thermogravimetry (TG/DTG) revealed an increase in amorphous cellulose and partial delignification in lignocellulosic structures as a consequence of SE-[Bmim]Cl. The pretreatment of both SE and SE-[Bmim]Cl yielded lignocellulosic substrates with improved properties in terms of their conversion into glucose and bio H2. Five-to-ten folds (by SE) and ten-to-twenty folds (by SE-[Bmim]Cl) more glucose was released from the lignocellulosic substrates of both plants than those of contrast samples. Compared to Phragmites (sp.), the greater destruction in lignocellulosic structure of Thalia dealbata as a result of SE and SE-[Bmim]Cl, increased the accessible surface area and disrupted the cellulose crystallinity much more, thus making it more efficient for bio H2 conversion. The bio H2 of 1.97 ± 0.14 mmol H2 g−1 dry weight (DW) was yielded after sludge anaerobic fermentation of Thalia dealbata treated with SE, and it increased to 4.79 ± 0.86 mmol H2 g−1 DW after SE-[Bmim]Cl treatment. For Phragmites (sp.) it was 1.45 ± 0.42 and 2.75 ± 0.76 mmol H2 g−1 DW after SE and SE-[Bmim]Cl pretreatment, respectively. Therefore, SE-[Bmim]Cl pretreatment can be developed for efficiently enhancing bio H2 conversion from wetland plant Thalia dealbata.
Introduction
The extensive consumption of fossil fuel has created global environmental issues. Eutrophication has become an environmental issue worldwide. Moreover, the increasingly worsening water quality induced by excessive nitrogen (N) and phosphorus (P) inputs has speeded up water eutrophication.1,2 The constructed wetland, being an effective eutrophication treatment technology, is in increasing operation to mitigate deterioration wastewater pollutants and nutrients inputs.3–6 However, a lot of lignocellulosic biomass is produced through wastewater treatment, which makes the constructed wetland the cellulosic biofuel production system, where highly productive plant species have great potential.7 Green reed (Phragmites (sp.)) is a new variety of common reed (Phragmites australis). Not only do both Phragmites species have approximately three to five times greater productivity than the dedicated biofuel crop, i.e. s switchgrass, but also they can reduce N use, thus making them preferable wetland plants.8–10 Water arum (Thalia dealbata) is another highly productive wetland plant for N removal from wastewater in east China.11,12 Because of increase in the requirements of renewable energy, increasing attention has focused on harvest of the existing strands of wetland biomass for cellulosic biofuel conversion.13,14The big challenge in bio H2 transformation from wetland biomass is the pretreatment of lignocellulosic substrates, which is strongly conditioned by bioaccessibility of cellulose.15–17 The aim of pretreatment is to increase the accessibility of enzymes to cellulose by removing lignin and hemicelluloses components and to reduce cellulose crystallinity in substrates.18 Better pretreatment, leading to solubilisation of celluloses, is also recommended as the efficient process for bio H2 conversion.19,20 Therefore, adequate pretreatment is of great interest to fractionate the lignocelluloses better. Steam explosion (SE) pretreatment can provide effective fractionation of lignocellulosic components and generate cellulose-rich fractions through hemicelluloses degradation due to destruction of cell-wall matrix.21,22 Furthermore, cellulose components can be dissolved without derivatization in high concentrations of ionic liquids (ILs), thus making SE and ILs the attractive biomass pretreatments.23–28 In addition, compared to conventional pretreatments, the advantages of SE and ILs have a significantly lower environmental impact and less use of hazardous chemicals is observed.
ILs are known to dissolve cellulose by effectively disrupting the complex network of non-covalent interactions between carbohydrates and lignin in biomass. The 1-butyl-3-methyl-imidazolium chloride ([Bmim]Cl) was found to be the most effective IL capable of dissolving up to 25% (wt/wt) cellulose25 due to chloride anion (Cl−) present responsible for dissolution of cellulose.29 Cellulose was disordered in [Bmim]Cl solution during the dissolution process as measured by high-resolution carbon-13 nuclear magnetic resonance (13C NMR), where no degradation of cellulose occurred.24,30–32 The addition of water, alcohol or acetone as anti-solvent resulted in a precipitation of cellulose from the [Bmim]Cl solution.33,34 The assistance of microwave irradiation can enhance efficiency of dissolution in ILs compared to ordinary thermal treatment.35 In addition, ILs were recovered up to 97% (wt/wt) of the initial mass, and thus it can be reused after ILs pretreatment which did not alter the 1H and 13C NMR spectra of the used ILs compared to the purity of ILs before pretreatments.36 Therefore, the objectives of this study (as illustrated in Scheme 1) are as follows: (1) to perform the pretreatment with SE and IL ([Bmim]Cl as a solvent with microwave-assistance) on lignocellulosic substrates of both Phragmites (sp.) and Thalia dealbata in constructed wetland; (2) to characterize the compositional (cellulose, hemicelluloses, lignin) and structural features (crystallinity of cellulose, lignin, surface area and porosity) of lignocellulosic substrates through scanning electron microscopy (SEM), Fourier transform infrared spectroscopy (FTIR), BET surface area and porosity, and thermogravimetry/differential thermogravimetry (TG/DTG); and (3) to evaluate enzymatic hydrolysis, sludge anaerobic fermentation and bio H2 yield with HPLC and GC measurements.
 | ||
| Scheme 1 The bio H2 conversion strategies of lignocellulosic substrates of wetland plants under the pretreatments by SE and [Bmim]Cl (P(s) and TD referring to Phragmites (sp.) and Thalia dealbata, respectively; SEP(s) and SETD representing SE-pretreated Phragmites (sp.) and Thalia dealbata, respectively; SEILP(s) and SEILTD representing SE-[Bmim]Cl pretreated Phragmites (sp.) and Thalia dealbata, respectively). | ||
Results and discussion
Chemical composition of wetland lignocellulosic biomass
Phragmites (sp.) and Thalia dealbata are large-biomass and fast-growth wetland plants in the subsystem of constructed wetlands located at Linan municipal wastewater treatment plant, Zhejiang Province, China.10,12 Phragmites (sp.) are planted once and can grow for many years with relatively low maintenance, while Thalia dealbata belongs to the annual grass family. The lignocellulosic components consist of cellulose, hemicelluloses and lignin accounting for 37–43%, 23–33% and 13–14% of the dried biomass, respectively, as listed in Table 1. Phragmites (sp.) stalk enriches higher contents of cellulose and hemicellulose as compared to Thalia dealbata stalk.| Sample | Cellulose | Hemicellulose | Lignin |
|---|---|---|---|
| (%) | (%) | (%) | |
| Water arum (Thalia dealbata) | |||
| Raw | 37.7 ± 0.3 | 24.3 ± 1.2 | 14.0 ± 0.3 |
| SE-pretreated (2.5 MPa) | 37.2 ± 0.6 | 5.7 ± 0.8 | 14.8 ± 0.3 |
| [Bmim]Cl-regenerated (2.5 MPa) | 37.1 ± 0.1 | 5.4 ± 0.2 | 14.6 ± 0.1 |
| Green reed (Phragmites (sp.)) | |||
| Raw | 42.8 ± 0.1 | 31.9 ± 1.2 | 13.0 ± 0.1 |
| SE-pretreated (3.0 MPa) | 43.9 ± 0.9 | 6.8 ± 1.0 | 11.5 ± 0.1 |
| [Bmim]Cl-regenerated (3.0 MPa) | 45.0 ± 0.2 | 6.0 ± 0.2 | 11.3 ± 0.2 |
SE pretreatment of wetland lignocellulosic biomass
As seen in Fig. 2, slight increase in cellulose appeared in SE-pretreated Phragmites (sp.) lignocelluloses but not in SE-pretreated Thalia dealbata lignocelluloses. Under lower steam pressure of 2.0 and 2.5 MPa, increasing the residence time from 60 s to 90 s slightly decreased the cellulose in SE-pretreated Phragmites (sp.) lignocelluloses (Fig. 2a). Furthermore, no impact on lignin removal was seen, but slightly increased lignin appeared in SE-pretreated Thalia dealbata lignocelluloses (Table 1). SE-pretreatment disrupted the lignocellulosic structure through degradation of hemicellulosic components and generation of lignin-related components. These impacts were closely related to the type of raw materials. The optimal cellulose-rich fractionation in SE-pretreated Phragmites (sp.) lignocelluloses yielded under steam pressure of 3.0 MPa, while for Thalia dealbata it was at 2.5 MPa.The increase of steam pressure and residence time significantly reduced total lignocellulosic compositions in SE-pretreated lignocelluloses of both plants compared to the raw material with hemicellulose removal taking place much more than cellulose and lignin (Fig. 1 and 2). Lower steam pressure caused the slight removal of hemicelluloses, while higher steam pressure generated the significant removal of hemicelluloses from SE-pretreated lignocelluloses of both plants. The considerable hemicellulose removal occurred at steam pressure of 3.0 MPa for Phragmites (sp.), while for Thalia dealbata it was at 2.5 MPa (Fig. 2, Table 1).
 | ||
| Fig. 1 Total compositions (cellulose, hemicellulose and lignin) of the SE-pretreated stalk lignocelluloses from (a) Phragmites (sp.) and (b) Thalia dealbata under different steam pressure and residence time. | ||
 | ||
| Fig. 2 Contents of cellulose, hemicelluloses and lignin of the SE-pretreated stalk lignocelluloses from (a) Phragmites (sp.) and (b) Thalia dealbata under different steam pressure and residence time. | ||
Enzymatic hydrolysis of SE-pretreated lignocelluloses
The optimization of enzymatic saccharification was performed on SE-pretreated Thalia dealbata lignocelluloses (2.0–3.0 MPa, 90 s) as substrate. Low activities of enzymatic hydrolysis in raw material but considerably increased activities in SE-pretreated lignocelluloses were obtained. The optimal enzymatic hydrolysis appeared at substrate concentration of 40 g L−1 (Fig. 3a), enzyme concentration >20 FPU g−1 (substrate) (Fig. 3b), pH 4.8 (Fig. 3c), 50 °C (Fig. 3d) and hydrolysis time of 24 h (Fig. 3e). Under the optimal hydrolysis conditions, the enzymatic hydrolysis rate of SE-pretreated lignocelluloses from both plants increased significantly, about ten times higher than those of the raw material (Fig. 4). Enzymatic hydrolysis highly depended on the nature of lignocellulosic substrates and SE-pretreated conditions. The increase in steam pressure and residence time presented increased trends in enzymatic hydrolysis rate of SE-pretreated lignocelluloses compared with that of Thalia dealbata, which increased faster than Phragmites (sp.). Compared to Phragmites (sp.), lignocellulosic components of Thalia dealbata were affected much more by SE pretreatment, facilitating the accessibility of cellulose in substrate. Enzymatic hydrolysis rate was around 61.3% for Thalia dealbata under 2.5 MPa, while it was 62.9% under high steam pressure of 3.0 MPa for Phragmites (sp.) (Fig. 4). | ||
| Fig. 3 Optimization of saccharification reaction for SE-fractioned stalks lignocelluloses from Thalia dealbata. (a) Enzyme concentration (g L−1); (b) substrate concentration (FPU g−1 substrate); (c) pH; (d) hydrolysis temperature (°C); (e) hydrolysis time (h). | ||
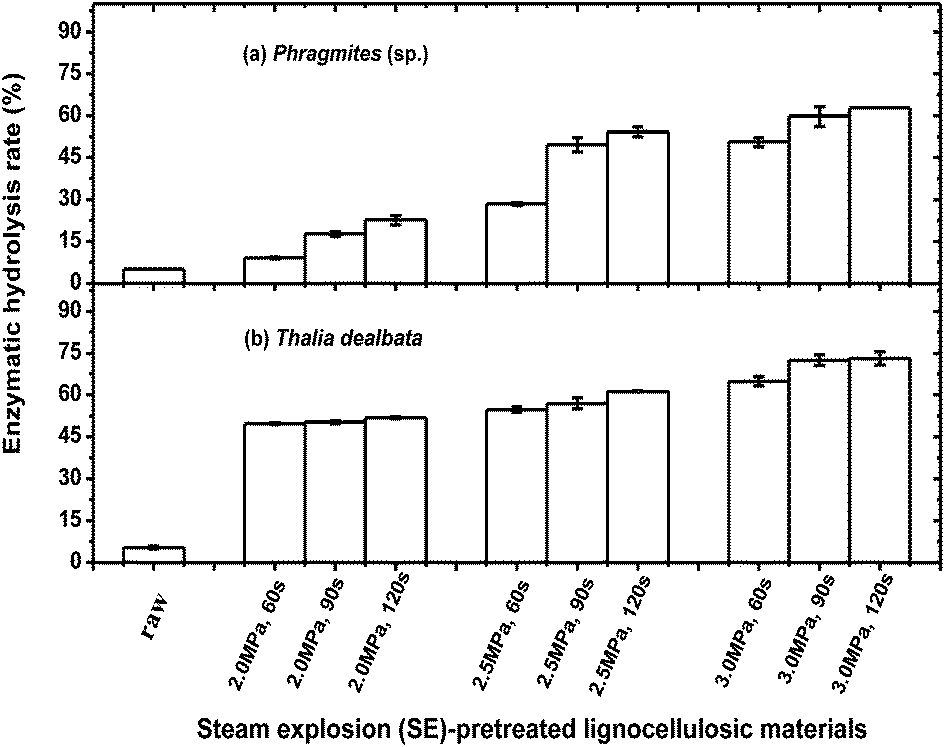 | ||
| Fig. 4 Enzymatic hydrolysis rates of SE-pretreated lignocelluloses from (a) Phragmites (sp.) and (b) Thalia dealbata under different steam pressure and residence time. | ||
[Bmim]Cl dissolution of SE-pretreated lignocelluloses
Lignocellulosic substrates present compositional and structural features limiting their accessibility of celluloses for bio H2 conversion. In this study, the optimization of microwave-assisted solubilisation in IL ([Bmim]Cl as solvent) was developed by using SE-pretreated Phragmites (sp.) lignocelluloses (3.0 MPa, 90 s) as substrate. This solubilisation in [Bmim]Cl depended on the nature of lignocellulosic materials and microwave conditions (microwave time, microwave power, and initial concentration of lignocelluloses in [Bmim]Cl). The optimal microwave-assisted [Bmim]Cl dissolution conditions were 400 W, 40 s and a 1/20 ratio of SE-pretreated lignocelluloses in [Bmim]Cl (Fig. 5), by which cellulose can be regenerated from [Bmim]Cl solution through addition of sufficient de-ionized water as anti-solvent. Therefore, SE-pretreated Phragmites (sp.) lignocelluloses (3.0 MPa) and Thalia dealbata lignocelluloses (2.5 MPa) performed optimal microwave assisted-[Bmim]Cl treatment. As seen in Table 1, the slightly decreased hemicellulose and lignin appeared in [Bmim]Cl-regenerated Phragmites (3.0 MPa, 90 s) and Thalia dealbata (2.5 MPa, 90 s) lignocelluloses. Moreover, slightly enhanced cellulose appeared in [Bmim]Cl-regenerated Phragmites (sp.) lignocelluloses;37 however, significant changes in lignocellulosic structures may occur. | ||
| Fig. 5 Optimization of microwave assisted-[Bmim]Cl dissolution of SE-pretreated Phragmites (sp.) lignocelluloses (3.0 MPa, 90 s) ((a) microwave time; (b) microwave power; (c) solid/liquid ratio of lignocelluloses in [Bmim]Cl). | ||
The raw material untreated with SE (Fig. 4), followed by microwave assisted-[Bmim]Cl treatment (Fig. 6), presented a pretty low enzymatic hydrolysis rate of around 9%. In contrast, a significant increase in enzymatic hydrolysis rate was noticed in [Bmim]Cl-regenerated lignocelluloses of both plants. As compared to SE-pretreated material (Fig. 4), SE-[Bmim]Cl treatment led to 2 times enzymatic hydrolysis rate increase, reaching 110% and 130% of regenerated Phragmites (sp.) and Thalia dealbata lignocelluloses, respectively (Fig. 6). This indicates that all celluloses and a part of hemicelluloses were hydrolyzed from [Bmim]Cl-regenerated lignocelluloses, resulting in ten-to-twenty-fold increase in glucose released in hydrolysates. The cellulose partially hydrolyzed from SE-pretreated lignocelluloses, as is evident by five-to-ten-fold glucose increase in hydrolysates (Table 2). The hydrolytic enzymes worked most efficiently for cellulose and less for hemicellulose; therefore, the enzymatic saccharification byproducts in the hydrolysates were mainly glucose with less xylose and cellobiose (Table 2).
 | ||
| Fig. 6 Enzymatic hydrolysis rates of [Bmim]Cl-regenerated lignocelluloses from (a) SE-pretreated Phragmites (sp.) and (b) SE-pretreated Thalia dealbata under different steam pressure and residence time. | ||
| Sample | Glucose | Xylose | Cellobiose |
|---|---|---|---|
| % (wt/wt biomass) | % (wt/wt biomass) | % (wt/wt biomass) | |
| a NR: no detected results. | |||
| Thalia dealbata | |||
| Raw | 7.81 ± 0.22 | 2.68 ± 0.25 | NR |
| SE-pretreated (2.5 MPa) | 38.65 ± 7.49 | 0.38 ± 0.19 | NR |
| [Bmim]Cl-regenerated (2.5 MPa) | 72.11 ± 0.20 | 0.62 ± 0.15 | 2.59 ± 0.22 |
| Phragmites (sp.) | |||
| Raw | 4.54 ± 0.43 | NR | NR |
| SE-pretreated (3.0 MPa) | 35.33 ± 0.96 | 2.19 ± 0.16 | NR |
| [Bmim]Cl-regenerated (3.0 MPa) | 88.80 ± 0.23 | 1.27 ± 0.03 | 3.17 ± 0.12 |
Differences observed only by SE and SE-[Bmim]Cl pretreatments could be due to their significant difference on degradation process of substrates. Treatment with SE-[Bmim]Cl can depolymerize cellulose more efficiently by disrupting the intra- and inter-hydrogen bonds of cellulose molecules, resulting in the fragmental cellulosic materials with highly amorphous structure.34 Thus, better enzyme accessibility was achieved with the significantly accelerated enzymatic hydrolysis rate and high production of reducing sugar from [Bmim]Cl-regenerated materials.38
Bio H2 fermentation
Thermophilic fermentation of lignocelluloses remained very slow or inhibited in general,39 as evident by the low bio H2 yield of 1.22–1.26 mmol H2 g−1 dried weight (DW) of raw materials (Table 3). After SE pretreatment, bio H2 of 1.45–1.97 mmol H2 g−1 DW was produced in lignocelluloses of both plants, and it was increased to 2.75–4.79 mmol H2 g−1 DW after SE-[Bmim]Cl treatment (Table 3).| Sample | Bio H2 |
|---|---|
| (mmol H2 g−1 DW) | |
| Thalia dealbata | |
| Raw | 1.22 ± 0.25 |
| SE-pretreated (2.5 MPa) | 1.97 ± 0.14 |
| [Bmim]Cl-regenerated (2.5 MPa) | 4.79 ± 0.86 |
| Phragmites (sp.) | |
| Raw | 1.26 ± 0.08 |
| SE-pretreated (3.0 MPa) | 1.45 ± 0.42 |
| [Bmim]Cl-regenerated (3.0 MPa) | 2.75 ± 0.76 |
Fig. 7 summarizes material balance for pretreatment, saccharification and fermentation of both Thalia dealbata and Phragmites (sp.) in two scenarios (also illustrated in Scheme 1): (a) SE pretreatment and (b) SE-[Bmim]Cl pretreatment. On the basis of 100 g of plant biomass, the majority of hemicelluloses degraded, producing cellulose-rich lignocelluloses after SE pretreatment, and enhancing bio H2 yield after saccharification and fermentation (Table 1 and Fig. 7a). As compared to SE-pretreated samples, SE-[Bmim]Cl pretreatment on these cellulose-rich lignocellulosic structures resulted in enhanced bio H2 yield by above 2-fold (Table 3 and Fig. 7b), which could be attributed to 2-fold enhancements in both enzymatic hydrolysis rate and glucose released from [Bmim]Cl-regenerated lignocelluloses (Fig. 4 and 6, Table 2). SE-[Bmim]Cl pretreatment facilitated efficient transformation of cellulose in substrate into soluble polysaccharide monomer sugars (i.e., glucose) with higher yield, causing a positive effect on bio H2 conversion (Tables 2 and 3, Fig. 7b).
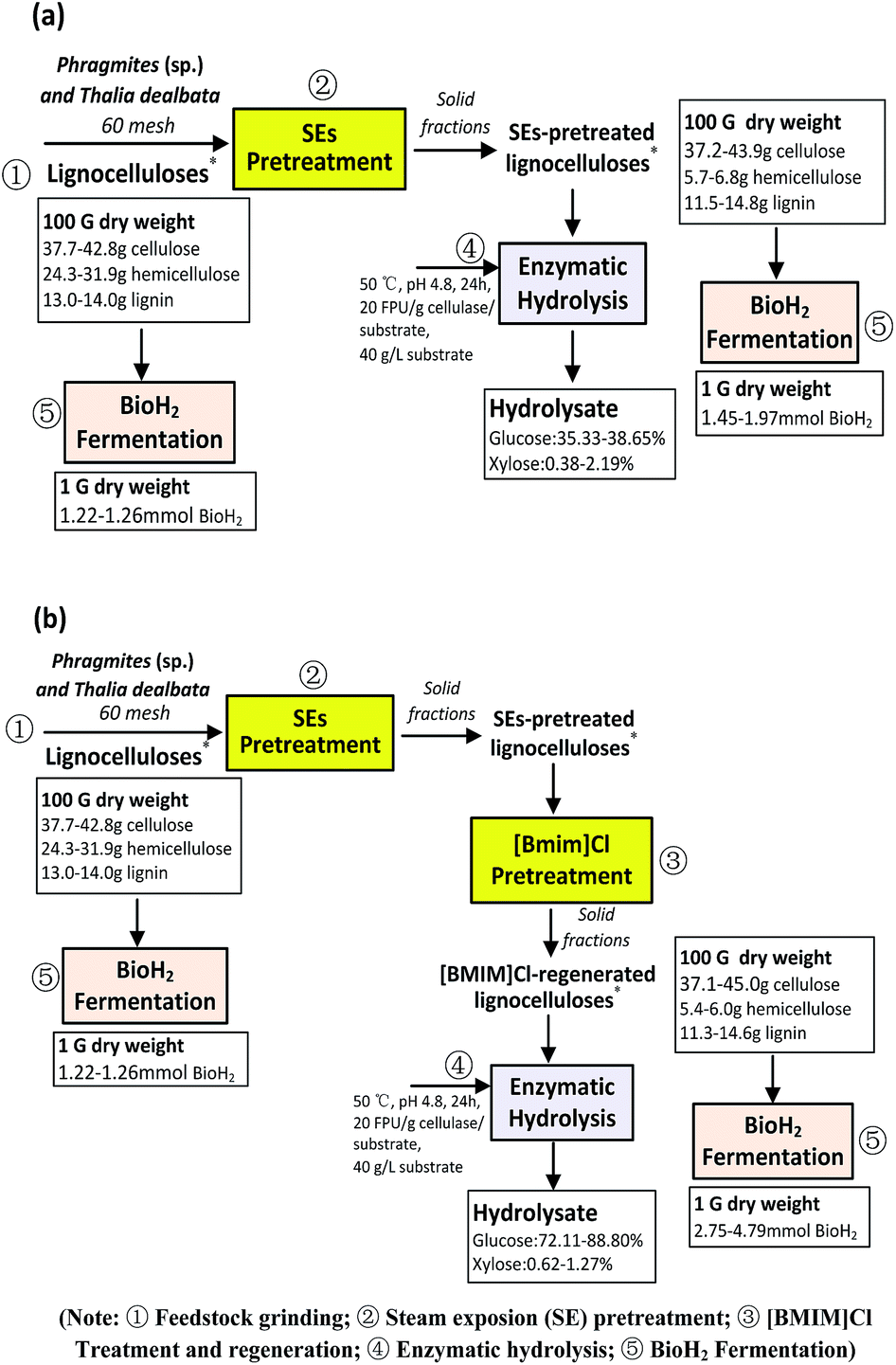 | ||
| Fig. 7 Material balance on proposed pretreatments ((a) SE, (b) SE-[Bmim]Cl), saccharification and fermentation processes. | ||
After SE-[Bmim]Cl treatment, bio H2 yield converted from annual grasses Thalia dealbata was much higher than that from corn or rice straw.14 However, the comparatively lower bio H2 yield of 2.75 ± 0.76 mmol H2 g−1 DW was converted from Phragmites (sp.) (Table 3). The utilization mechanism of lignocelluloses could be different between Thalia dealbata and Phragmites (sp.). The lignocellulosic structures of Phragmites (sp.) exhibited more resistance to deconstruction than annual grass Thalia dealbata. Therefore, there is a need for further elucidation of pretreatment by SE and SE-[Bmim]Cl on compositional and structural features of lignocelluloses during bio H2 conversion.
Structure and crystallinity assessment of lignocelluloses
The major obstacle in utilizing wetland lignocelluloses as fermentation substrates is recalcitrance of the biomass itself. Both Phragmites (sp.) and Thalia dealbata enrich cellulose and hemicellulose as main lignocellulosic components with different lignocellulosic structures, affecting their enzymatic hydrolysis, anaerobic fermentation as well as bio H2 yield. The pretreatments on factors affecting the structural features, such as crystallinity of cellulose, accessible surface area and lignin in lignocellulosic substrates, have already been discussed.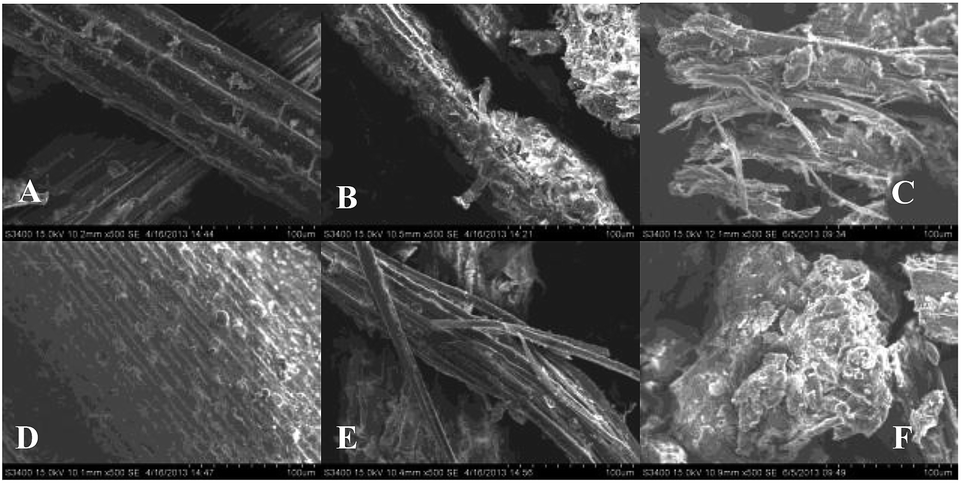 | ||
| Fig. 8 SEM images of raw and pretreated lignocellulosic materials. A, B and C representing the raw, SE-pretreated and [Bmim]Cl-regenerated Thalia dealbata (2.5 MPa, 90 s), respectively; D, E and F representing the raw, SE-pretreated and [Bmim]Cl-regenerated Phragmites (sp.) (3.0 MPa, 90 s), respectively. All images were taken at same scale (×500). | ||
 | ||
| Fig. 9 FTIR spectrum of (a) Thalia dealbata (raw, SE-pretreated and [Bmim]Cl-regenerated Thalia dealbata (2.5 MPa, 90 s)) and (b) Phragmites (sp.) (raw, SE-pretreated and [Bmim]Cl-regenerated Phragmites (sp.) (3.0 MPa, 90 s)). | ||
| Parameters | Raw | SE pretreated |
|---|---|---|
| Thalia dealbata | ||
| BET surface area (m2 g−1) | 1.47 | 2.81 |
| Langmuir surface area (m2 g−1) | 2.46 | 5.04 |
| BJH adsorption cumulative surface area of pores (m2 g−1) | 1.57 | 2.78 |
| BJH adsorption cumulative volume of pores (×10−3 cm3 g−1) | 5.14 | 9.36 |
| BJH adsorption average pore diameter (nm) | 131.13 | 134.75 |
| Phragmites (sp.) | ||
| BET surface area (m2 g−1) | 0.70 | 0.50 |
| Langmuir surface area (m2 g−1) | 0.95 | 0.65 |
| BJH adsorption cumulative surface area of pores (m2 g−1) | 0.27 | 0.27 |
| BJH adsorption cumulative volume of pores (×10−3 cm3 g−1) | 3.49 | 5.21 |
| BJH adsorption average pore diameter (nm) | 524.35 | 784.97 |
![[double bond, length as m-dash]](https://www.rsc.org/images/entities/char_e001.gif) O) between hemicelluloses and lignin (Fig. 9). These were both strengthened in the raw materials, and they were greatly reduced by both pretreatments. In addition, the weakened peak shapes and intensities at 1250 cm−1 (assigned to hemicellulose) appeared after pretreatments. This could be attributed to the significant removal of hemicelluloses by SE and SE-[Bmim]Cl treatments (Fig. 1 and 2). The band at 1631 cm−1, assigned to absorbed water bending vibration, was significantly reduced by SE-[Bmim]Cl pretreatment, which could be related to the decreased O–H group in substrate (Fig. 9). The interaction between free Cl− in [Bmim]Cl and O–H groups disrupted the hydrogen bonds present within and between the lignocellulosic substrates.
O) between hemicelluloses and lignin (Fig. 9). These were both strengthened in the raw materials, and they were greatly reduced by both pretreatments. In addition, the weakened peak shapes and intensities at 1250 cm−1 (assigned to hemicellulose) appeared after pretreatments. This could be attributed to the significant removal of hemicelluloses by SE and SE-[Bmim]Cl treatments (Fig. 1 and 2). The band at 1631 cm−1, assigned to absorbed water bending vibration, was significantly reduced by SE-[Bmim]Cl pretreatment, which could be related to the decreased O–H group in substrate (Fig. 9). The interaction between free Cl− in [Bmim]Cl and O–H groups disrupted the hydrogen bonds present within and between the lignocellulosic substrates.FTIR spectra in the 1800–800 cm−1 regions were characteristic of the cellulose structure. The bands at 1421 (or 1426 cm−1) and 896 cm−1, assigned to CH2 scissoring motion and C–O–C stretching in cellulose, respectively, are quite sensitive to the amount of crystalline and amorphous cellulose.40 The infrared ratio, H1421/H896 (or H1426/H896), is commonly called lateral order index (LOI) used to determine the amount of crystalline cellulose.40,41 Total crystallinity index (TCI), the ratio of H1376/H2918 or H1376/H2921 (CH and CH2 stretching), is also used to study the crystallinity changes.41,42
As seen in Table 5, high values of LOI and TCI in the raw materials indicated that both Thalia dealbata and Phragmites (sp.) had high crystallinity of cellulose and ordered lignocellulosic structures before pretreatments. As compared to the raw material, pretreatment with SE dramatically increased the LOI values for both plants, while TCI values was slightly increased for Thalia dealbata but slightly decreased for Phragmites (sp.). Comparatively, SE-[Bmim]Cl treatment significantly decreased the values of both LOI and TCI for both plants. No destruction of cellulose crystallinity appeared by SE pretreatment, while the disruption of ordered lignocellulosic structures occurred by SE-[Bmim]Cl treatment. This was in good agreement with the observation from SEM images. During solubilisation in [Bmim]Cl, the cellulosic materials lost their crystalline structures and restructured themselves into mostly amorphous forms, which made structures of regenerated lignocelluloses less crystalline. Due to the disruption of ordered structure and reduction in crystallinity, SE-[Bmim]Cl pretreated materials became more susceptible to enzymatic attack in the subsequent hydrolysis process and bio H2 conversion (Tables 2 and 3).
| Substrates | LOI | TCI |
|---|---|---|
| Thalia dealbata | ||
| Raw | 4.12 | 1.11 |
| SE-pretreated (2.5 MPa) | 5.53 | 1.22 |
| [Bmim]Cl-regenerated (2.5 MPa) | 2.43 | 0.79 |
| Phragmites (sp.) | ||
| Raw | 3.00 | 1.20 |
| SE-pretreated (3.0 MPa) | 3.63 | 1.16 |
| [Bmim]Cl-regenerated (3.0 MPa) | 2.34 | 0.80 |
In addition, the peak intensities at 1250 cm−1 (assigned to hemicellulose), 1160 cm−1 (related to carbohydrates) and 1060 cm−1 (assigned to cellulose) were weakened for Thalia dealbata but strengthened for Phragmites (sp.) (Fig. 9), suggesting the greater destruction in lignocellulosic structures of Thalia dealbata occurred by SE pretreatment. Furthermore, the fast decrease in values of both LOI and TCI by SE-[Bmim]Cl indicated the occurrence of greater disruption of cellulose crystallinity for Thalia dealbata. Therefore, the increased amorphous cellulose in [Bmim]Cl-regenerated Thalia dealbata led to a better performance of enzymatic hydrolysis,37,41 resulting in bio H2 conversion with high yield (Table 3).
 | ||
| Fig. 10 TG/DTG curves of (a) Thalia dealbata (raw, SE-pretreated and [Bmim]Cl-regenerated Thalia dealbata (2.5 MPa, 90 s)) and (b) Phragmites (sp.) (raw, SE-pretreated and [Bmim]Cl-regenerated Phragmites (sp.) (3.0 MPa, 90 s)). | ||
The raw Phragmites (sp.) lignocelluloses had a more clear-cut separation between Tshoulder (300 °C, related to hemicelluloses) and Tpeak (350 °C, related to cellulose) (Fig. 10). While the raw Thalia dealbata exhibited a wide peak at 300 °C stemming from the thermal decomposition of cellulose, and the curve of hemicellulose was almost merged compared to cellulose, the similar behavior of lignocellulosic substrate was found in willow and wood.43 As seen from the DTG curves (Fig. 10), the amount of both cellulose and hemicellulose in the raw Thalia dealbata was relatively smaller than those of the raw Phragmites (sp.), which was in good accordance with lignocellulosic components analysis (Table 1).
The degradation of Phragmites (sp.) occurred within a narrower range of temperatures (Tpeak) than that of Thalia dealbata, even after pretreatments with SE and SE-[Bmim]Cl. This was mainly caused by the different sorts of hemicelluloses components and crystalline structures of cellulose as well as their variable distributions in plant species.45 No big changes occurred in Tpeak of Phragmites (sp.) after pretreatments with SE and SE-[Bmim]Cl. In contrast, SE-pretreatment shifted the Tpeak of Thalia dealbata towards higher T (305–325 °C) and a further higher Tpeak (338 °C) was attained by SE-[Bmim]Cl treatment (Fig. 8a). This could be attributed to the increased cellulose contents through removal of hemicelluloses as a consequence of SE pretreatment, while SE-[Bmim]Cl treatment increased amorphous cellulose components, thus increasing solid thermostability. The enhanced thermal stability of SE-[Bmim]Cl pretreated biomass appeared at higher temperatures was in agreement with the results from Labbe et al.46 and Singh et al.47 In addition, the similar TG/DTG pattern occurred in regenerated materials of both plants after SE-[Bmim]Cl treatment, with Tpeak around 338 °C for Thalia dealbata and 350 °C for Phragmites (sp.). This could be indicative of cellulose molecules of different polymerization degree, suggesting that higher polymerization degree of cellulose were in regenerated materials of Phragmites (sp.) than Thalia dealbata.
The presence of lignin was one of the most important factors affecting biotransformation of lignocellulosic substrates into bio H2. A broad peak ranging from 410 to 550 °C mainly corresponded to lignin, which overlapped partially with those of hemicelluloses and cellulose. SE-pretreatment changed intensities of this wide peak and it was shifted towards higher T at 530 °C for Thalia dealbata but towards lower T at 485 °C for Phragmites (sp.) (Fig. 10). This could be related to the formation of different thermolabile chemical bonds in lignin fractionation as a consequence of SE. Furthermore, higher T above 550 °C was attained for both plants as a consequence of SE-[Bmim]Cl (Fig. 10). The alteration of lignin structure occurred in [Bmim]Cl-regenerated materials, which could be attributed to the partially dissolution of lignin components in [Bmim]Cl (delignification). However, the structures and compositions of lignin in lignocellulosic substrates of both plants need be further elucidated before and after treatments. Furthermore, the combined [Bmim]Cl filtrates after use were concentrated and the regenerated [Bmim]Cl solvent was measured for the mass. The recovery rate of regenerated [Bmim]Cl solvent was as high as 91%; therefore, it can be reused for the dissolution of lignocellulosic substrates during bio H2 transformation. Future investigation will focus on more structural elucidation of lignin components in regenerated [Bmim]Cl solvent in order to produce high value products of phenol compounds.
Conclusions
Compared to Thalia dealbata, Phragmites (sp.) displayed higher levels of cellulose and hemicellulose in substrate with high polymerization degree of cellulose, but possessed lower surface area and pores volume compared to cellulose. Pretreatments with SE and SE-[Bmim]Cl were performed on the lignocellulosic substrate of both wetland plants. The optimal cellulose-rich fractionation of Phragmites (sp.) lignocelluloses was obtained as a consequence of SE pretreatment under steam pressure at 3.0 MPa, while for Thalia dealbata lignocelluloses it was under steam pressure at 2.5 MPa. Compared to Phragmites (sp.), SE pretreatment greatly destroyed lignocellulosic structure of Thalia dealbata through hemicelluloses removal, thus increasing the accessible surface area and pore volume of cellulose in substrate. Furthermore, pretreatment with SE-[Bmim]Cl on lignocellulosic structure of plants effectively disrupted crystalline structure of cellulose or even solubilize some lignin components, facilitating the formation of amorphous cellulose in regenerated materials with enhanced accessible surface area.SE-[Bmim]Cl pretreatment led to lignocellulosic structures with improved properties efficient for their transformation into glucose and bio H2 with elevated yields. Most importantly, higher bio H2 conversion was obtained from lignocelluloses of Thalia dealbata compared to Phragmites (sp.), which possess denser lignocellulosic structure less efficient for bio H2 conversion. To better understand the transformation of cell wall components into biofuel from wetland lignocellulosic substrates, further investigations will focus on more structural elucidation of hemicelluloses and lignin components in the products obtained from the pretreated samples as a result of SE and SE-[Bmim]Cl treatments.
Experiment section
Materials
Lignocellulosic substrates used in this study were Phragmites (sp.) and Thalia dealbata from constructed wetlands located in Linan municipal wastewater treatment plant, Zhejiang Province, China. The harvested stalks of Phragmites (sp.) and Thalia dealbata at their maturities in early winter were naturally air-dried, and then ground into 60 meshes using a cutting milling Restch, SM 100. The components of cellulose, hemicelluloses and lignin in stalks were evaluated according to the procedures from Van Soest.48 Lignocellulosic substrates were dried at 60 °C for 48 h, and then maintained under vacuum for use.2 N NaOH aqueous solution was prepared from NaOH pellets (of analytical grade purity), and 4 M HCl aqueous solutions was prepared from 37% (wt/wt) HCl (of analytical grade purity). 1-Butyl-3-methyl-imidazolium chloride ([Bmim]Cl) (>95.0% purity) was purchased from Shanghai Chengjie Chemical Reagent Ltd., China. Prior to use, [Bmim]Cl was dried under vacuum (0.1 Pa) at 70 °C (i.e. above the melting point) for at least 24 h to remove water completely.
Enzymatic hydrolysis was performed using 50 mM citric acid–sodium citrate buffer (pH 4.8), which was prepared from citric acid monohydrate (of analytical grade purity) and trisodium citrate (of analytical grade purity). Commercial enzyme cellulase (activity 100 FPU g−1) was purchased from Wuxi Xuemei Ltd., China. 3,5-Dinitrosalicylic acid (of analytical grade purity) was purchased from Aladdin (Shanghai) Reagent Ltd., China. D-Glucose, D-xylose, and cellobiose of spectroscopic grade purities were purchased from Aladdin (Shanghai) Reagent Ltd., China. Acetonitrile of HPLC grade purity was supplied by Sigma-Aldrich Co. FTIR samples were prepared with KBr (>99.0% trace metal basis) was purchased from Sigma-Aldrich Co.
Pretreatment of lignocellulosic substrate with SE
Steam explosion (SE) pretreatment was performed on a QBS-80 batch steam explosion apparatus (Hebi Gentle Bioenergy Co. Ltd., China), which consisted of a high pressure vessel, a steam generator, a material tank, a receiver and a rapid-opening ball valve. The capacity of vessel was 400 ml with a maximum operating pressure of 4.0 MPa. The ground stalk lignocelluloses (∼120 g) of Phragmites (sp.) or Thalia dealbata were subjected to the saturated steam in the vessel of each batch in apparatus, at ratio of water and lignocelluloses of 2![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :1 (ml g−1). SE conditions were optimized by variation of saturated steam pressures (2.0, 2.5 and 3.0 MPa) and residence times for the pressure of saturated steam (60, 90 and 120 s) at the temperature of 230 °C. The pressure of saturated steam was maintained in high pressure vessel by the controller in apparatus at constant values of 2.0, 2.5 and 3.0 MPa, respectively, when changing residence time of saturated steam pressure in vessel. Then, SE-fractionated lignocellulosic materials were recovered in the receiver and their components (cellulose, hemicelluloses and lignin) were measured following the method of Van Soest.48 The raw and SE-pretreated lignocellulosic materials were dried at 60 °C for at least 48 h and kept under vacuum for use.
:1 (ml g−1). SE conditions were optimized by variation of saturated steam pressures (2.0, 2.5 and 3.0 MPa) and residence times for the pressure of saturated steam (60, 90 and 120 s) at the temperature of 230 °C. The pressure of saturated steam was maintained in high pressure vessel by the controller in apparatus at constant values of 2.0, 2.5 and 3.0 MPa, respectively, when changing residence time of saturated steam pressure in vessel. Then, SE-fractionated lignocellulosic materials were recovered in the receiver and their components (cellulose, hemicelluloses and lignin) were measured following the method of Van Soest.48 The raw and SE-pretreated lignocellulosic materials were dried at 60 °C for at least 48 h and kept under vacuum for use.
Pretreatment of lignocellulosic substrate with SE-[Bmim]Cl
Microwave-assisted solubilisation in [Bmim]Cl solvent were performed on SE-pretreated lignocelluloses, which was optimized by changing ratios of lignocelluloses in [Bmim]Cl (varied from 1/5, 1/10, 1/20, 1/30 to 1/40 (wt/wt)), microwave power (among 80, 240, 400, 560 and 800 W), and microwave heating time (among 28, 40, 52, 64 and 72 s). Vacuum dried SE-pretreated lignocelluloses were mixed with vacuum-dried [Bmim]Cl (at ratio of 1/20 (wt/wt)) in different vials, and then the optimal microwave power was figured out by changing the pressure under 80, 240, 400, 560 and 800 W, respectively, for a heating time of 40 s. Then, the optimal microwave heating time was achieved under the ratio of 1/20 (wt/wt) and 400 W microwave power, and the optimal ratio of lignocelluloses in [Bmim]Cl was obtained under 400 W of microwave power and 40 s of heating time.Mixture of SE-pretreated lignocellulosic materials and [Bmim]Cl (at a 1/20 ratio) was used to carry on the microwave-assisted [Bmim]Cl dissolution under the optimized condition (40 s, 400 W). [Bmim]Cl-regenerated materials were achieved by adding the sufficient deionized water as anti-solvents, centrifuged and dried at 60 °C for 48 h, and then kept for enzymatic hydrolysis. Moreover, the components of cellulose, hemicelluloses and lignin in [Bmim]Cl-regenerated lignocelluloses were measured following the method of Van Soest.48 Regarding the recovery of [Bmim]Cl after use, the combined [Bmim]Cl filtrates were concentrated by Rotary Evaporator (EYELA, Japan) under vacuum at 60 °C and finally freeze-dried under vacuum (0.1 Pa) at 70 °C for at least 48 h. The regenerated [Bmim]Cl was measured for the mass to yield recovery rate.
Enzymatic hydrolysis
Saccharification hydrolysis occurred with 50 ml suspension of lignocellulosic substrates, using 10 ml of 50 mM citric acid–sodium citrate buffer (pH 4.8) and cellulase levels from 2.5 to 50 FPU g−1 of dry substrates. After adding cellulase, the vials were capped and placed in a HZ-9211KB rotary incubator (HUALIDA, China) at 50 °C and 180 rpm. One-milliliter sample was withdrew at specific time intervals, placed in a boiling (100 °C) water bath for 15 min to deactivate the enzymes, and then centrifuged at 13000g (Fresco 17, Thermo, USA). A portion of 0.2 ml was used to measure the total reducing sugars by dinitrosalicylic acid (DNS) assay49 with glucose, D-xylose and cellobiose (Aladdin) as standards. Residual filtrates were stored at −40 °C for subsequent sugar analyses by HPLC. Cellulase activity was determined by standard filter paper assay and expressed as filter paper units per gram (FPU). Hydrolysis rate of the reducing sugars from lignocellulosic materials was calculated as follows: hydrolysis rate (%) = [(reducing sugars weight × 0.9)/(dry lignocellulosic material weight × cellulose content)] × 100.
Bio H2 fermentation
Mixed anaerobic cultures used as the seed were obtained from anaerobic sludge digesters (as inoculum) at Linan municipal wastewater treatment plant, Zhejiang Province, China. The inoculum was first treated at 90 °C for 1 h to inhibit activity of methanogens and enrich in hydrogen producing bacteria. After lignocellulosic substrates were subjected to enzymatic hydrolysis for 24 h, the hydrolysates in 60 ml obtained together with non-hydrolyzed solid fractions were initially introduced into each stoppered 150 ml serum bottle. Seed sludges (as inoculum) in 20 ml were also added to each bottle and batch thermophilic fermentation experiments were continuously conducted at 60 °C with no additional nutrient medium solution. Initial pH was adjusted to 6.0 with 2 M NaOH or 4 M HCl. The headspaces of bottles were flushed with nitrogen gas to reach oxygen-free conditions. When pressure in the bottle headspace started to drop off indicating hydrogen consumption, then the experimental procedures ended. Biogas volume was monitored continuously with a water displacement method in the batch thermophilic fermentation experiments. Alkalified water was used to dissolve carbon dioxide and yield biogas.Characterization
Statistics
All statistical analyses were performed in SigmaPlot (version 11). Two-way ANOVA with Tukey test post hoc procedure was employed to evaluate whether the means were significantly different at p = 0.05.Acknowledgements
This work was supported by grants from the Key Program for International Cooperation Project of the Ministry of Science and Technology of China (no. 2010DFB33960), the Key Laboratory of Yangtze River Water Environment of Ministry of Education (Tongji University) of China (no. YRWEF201102), the China Scholarship Council (no. 2011833072), and National Key Technology R&D Program of the Ministry of Science and Technology of China (no. 2012BAC17B04).Notes and references
- V. H. Smith, G. D. Tilman and J. C. Nekola, Environ. Pollut., 1999, 100, 179–196 CrossRef CAS.
- L. J. Sherwood and R. G. Qualls, Environ. Sci. Technol., 2001, 35, 4126–4131 CrossRef CAS.
- H. Brix, Water Sci. Technol., 1994, 30, 209–223 CAS.
- K. Abe and Y. Ozaki, Soil Sci. Plant Nutr., 1998, 44, 599–607 Search PubMed.
- C. H. House, B. A. Bergmann, A. M. Stomp and D. J. Frederick, Ecol. Eng., 1999, 12, 27–38 CrossRef.
- L. L. Rectenwald and R. W. Drenner, Environ. Sci. Technol., 2000, 34, 522–526 CrossRef CAS.
- D. Liu, X. Wu, J. Chang, B. J. Gu, Y. Min, Y. Ge, Y. Shi, H. Xue, C. H. Peng and J. G. Wu, Nat. Clim. Change, 2012, 2, 190–194 CAS.
- M. R. Schmer, K. P. Vogel, R. B. Mitchell and R. K. Perrin, Proc. Natl. Acad. Sci. U. S. A., 2008, 105, 464–469 CrossRef CAS PubMed.
- N. Sathitsuksanoh, Z. G. Zhu, N. Templeton, J. A. Rollin, S. P. Harvey and Y.-H. P. Zhang, Ind. Eng. Chem. Res., 2009, 48, 6441–6447 CrossRef CAS.
- F. L. Zhao, W. D. Yang, Z. Zeng, H. Li, X. E. Yang, Z. L. He, B. H. Gu, M. T. Rafiq and H. Y. Peng, Biomass Bioenergy, 2012, 42, 212–218 CrossRef CAS PubMed.
- S. X. Zhu, H. L. Ge, Y. Ge, H. Q. Cao, D. Liu, J. Chang, C. B. Zhang, B. J. Gu and S. X. Chang, Ecol. Eng., 2010, 36, 1307–1313 CrossRef PubMed.
- H. Chen and M. M. Zhang, Environ. Sci. Technol., 2013, 47, 8157–8163 CAS.
- B. J. Gu, D. Liu, X. Wu, Y. Ge, Y. Min, H. Jiang and J. Chang, Renewable Sustainable Energy Rev., 2011, 15, 4910–4916 CrossRef CAS PubMed.
- C. L. Cheng, Y. C. Lo, K. S. Lee, D. J. Lee, C. Y. Lin and J. S. Chang, Bioresour. Technol., 2011, 102, 8514–8523 CrossRef CAS PubMed.
- D. Li and H. Chen, Int. J. Hydrogen Energy, 2007, 32, 1742–1748 CrossRef CAS PubMed.
- R. Datar, J. Huang, P. C. Maness, A. Mohagheghi, S. Czemik and E. Chornet, Int. J. Hydrogen Energy, 2007, 32, 932–939 CrossRef CAS PubMed.
- L. Ozkan, T. H. Erguder and G. N. Demirer, Int. J. Hydrogen Energy, 2011, 36, 382–389 CrossRef CAS PubMed.
- M. Yoshida, Y. Liu, S. Uchida, K. Kawarada, Y. Ukagami, H. Ichinose, S. Kaneko and K. Fukuda, Biosci., Biotechnol., Biochem., 2008, 72, 805–810 CrossRef CAS PubMed.
- F. Monlau, C. Sambusiti, A. Barakat, X. M. Guo, E. Latrille, E. Trably, J. P. Steyer and H. Carrere, Environ. Sci. Technol., 2012, 46, 12217–12225 CrossRef CAS PubMed.
- F. Monlau, A. Barakat, E. Trably, C. Dumas, J. P. Steyer and H. Carrere, Crit. Rev. Environ. Sci. Technol., 2013, 43, 260–322 CrossRef CAS.
- J. E. Carrasco, C. M. C. Saiz, A. Navarro, P. Soriano, F. Saez and J. M. Martinez, Appl. Biochem. Biotechnol., 1994, 45–46, 23–24 CrossRef CAS.
- J. B. Li, G. Gellerstedt and K. Toven, Bioresour. Technol., 2009, 100, 2556–2561 CrossRef CAS PubMed.
- A. P. Dadi, C. A. Schall and S. Varanasi, Appl. Biochem. Biotechnol., 2007, 137, 407–421 CrossRef PubMed.
- S. P. M. da Silva, A. M. D. Lopes, L. B. Roseiro and R. Bogel-Lukasik, RSC Adv., 2013, 3, 16040–16050 RSC.
- R. P. Swatloski, S. K. Spear, J. D. Holbrey and R. D. Rogers, J. Am. Chem. Soc., 2002, 124, 4974–4975 CrossRef CAS PubMed.
- I. Kilpeläinen, H. Xie, A. King, M. Granstrom, S. Heikkinen and D. S. Argyropoulos, J. Agric. Food Chem., 2007, 55, 9142–9148 CrossRef PubMed.
- S. H. Lee, T. V. Doherty, R. J. Linhardt and J. S. Dordick, Biotechnol. Bioeng., 2009, 102, 1368–1376 CrossRef CAS PubMed.
- C. L. Li, B. Knierim, C. Manisseri, R. Arora, H. V. Scheller, M. Auer, K. P. Vogel, B. A. Simmons and S. Singh, Bioresour. Technol., 2010, 101, 4900–4906 CrossRef CAS PubMed.
- M. G. Adsul, A. P. Terwadkar, A. J. Varma and D. V. Gokhale, Bioresources, 2009, 4, 1670–1681 CAS.
- J. S. Moulthrop, R. P. Swatloski, G. Moyna and R. D. Rogers, Chem. Commun., 2005, 1557–1559 RSC.
- T. Heinze, K. Schwikal and S. Barthel, Macromol. Biosci., 2005, 5, 520–525 CrossRef CAS PubMed.
- A. Brandt, J. Grasvik, J. P. Hallett and T. Welton, Green Chem., 2013, 15, 550–583 RSC.
- M. E. Zakrzewska, E. Bogel-Lukasik and R. Bogel-Lukasik, Energy Fuels, 2010, 24, 737–745 CrossRef CAS.
- A. Brandt, J. P. Hallett, D. J. Leak, R. J. Murphy and T. Welton, Green Chem., 2010, 12, 672–679 RSC.
- V. M. Egorov, S. V. Smirnova, A. A. Formanovsky, I. V. Pletnev and Y. A. Zolotov, Anal. Bioanal. Chem., 2007, 387, 2263–2269 CrossRef CAS PubMed.
- A. M. D. Lopes, K. G. Joao, E. Bogel-Lukasik, L. B. Roseiro and R. Bogel-Lukasik, J. Agric. Food Chem., 2013, 61, 7874–7882 CrossRef PubMed.
- H. T. Tan, K. T. Lee and A. R. Mohamed, Carbohydr. Polym., 2010, 83, 1862–1868 CrossRef PubMed.
- H. Zhao, C. I. L. Jones, G. A. Baker, S. Xia, O. Olubajo and V. N. Person, J. Biotechnol., 2009, 139, 47–54 CrossRef CAS PubMed.
- Y. Lu, Q. H. Lai, C. Zhang, H. X. Zhao, K. Ma, X. B. Zhao, H. Z. Chen, D. H. Liu and X. H. Xing, Bioresour. Technol., 2009, 100, 2889–2895 CrossRef CAS PubMed.
- I. Spiridon, C. A. Teaca and R. Bodirlau, Bioresources, 2011, 6, 400–413 CAS.
- A. M. D. Lopes, K. G. Joao, D. F. Rubik, E. Bogel-Lukasik, L. C. Duarte, J. Andreaus and R. Bogel-Lukasik, Bioresour. Technol., 2013, 142, 198–208 CrossRef PubMed.
- A. Carrillo, X. Colom, J. J. Sunol and J. Saurina, Eur. Polym. J., 2004, 40, 2229–2234 CrossRef PubMed.
- W. H. Chen and P. C. Kuo, Energy, 2010, 35, 2580–2586 CrossRef CAS PubMed.
- V. I. Sharypov, N. Marin, N. G. Beregovtsova, S. V. Baryshnikov, B. N. Kuznetsov, V. L. Cebolla and J. V. Weber, J. Anal. Appl. Pyrolysis, 2002, 64, 15–28 CrossRef CAS.
- J. M. Heikkinen, J. C. Hordijk, W. de Jong and H. Spliethoff, Thermogravimetry as a tool to classify waste components to be used for energy generation, J. Anal. Appl. Pyrolysis, 2004, 71, 883–900 CrossRef CAS PubMed.
- N. Labbe, L. M. Kline, L. Moens, K. Kim, P. C. Kim and D. G. Hayes, Bioresour. Technol., 2012, 104, 701–707 CrossRef CAS PubMed.
- S. Singh, P. Varanasi, P. Singh, P. D. Adams, M. Auer and B. A. Simmons, Biomass Bioenergy, 2013, 54, 276–283 CrossRef CAS PubMed.
- Van Soest, Van Soest fiber analysis,http://www.aquaculture.ugent.be/Education/coursematerial/online%20courses/ATA/analysis/vsoest.htm.
- G. L. Miller, Anal. Chem., 1959, 31, 426–428 CrossRef CAS.
| This journal is © The Royal Society of Chemistry 2014 |