
Carbon dioxide bio-fixation and wastewater treatment via algae photochemical synthesis for biofuels production
Yafei Shen
*
Department of Environmental Science and Technology, Interdisciplinary Graduate School of Science and Engineering, Tokyo Institute of Technology, G5-8, 4259 Nagatsuta, Midori-ku, Yokohama, 226-8502, Japan. E-mail: yafeisjtu@gmail.com; shen.y.ad@m.titech.ac.jp; Fax: +81-45-924-5518; Tel: +81-45-924-5507
First published on 8th September 2014
Abstract
We are faced with the problem of energy/carbon dioxide (CO2) in the coming decades. Microalgae has been considered as one of the most promising biomass feedstocks for the production of biofuels. Meanwhile, the productivity of these photosynthetic microorganisms in converting CO2 into carbon-rich lipids, which are only a step or two away from biodiesel, significantly exceed that of agricultural crops without competing for arable land. Worldwide, research and demonstration programs are being carried out to develop the technologies required to expand algal lipid production from a craft to a major industrial process. This paper describes the recent advances on microalgae used for biofuel (e.g., biohydrogen, biodiesel and bioethanol) production, including their cultivation, harvesting, and processing. In addition, the various aspects associated with the design of microalgae production units are described, providing an overview of the current state of the development of algae cultivation systems (photobioreactors and open ponds). Algal cultivation systems integrated with algae-based biorefineries could yield diverse bioresources, such as biodiesel, green gasoline, bio-jet fuel, isolated proteins, food starches, textiles, organic fertilizers, which can mitigate the costs of biofuel production. Algae cultivation becomes a closed-loop engineered ecosystem by utilizing the energy, nutrients and CO2 held within residual waste materials to provide all necessary inputs except sunlight. Consequently, developing this biotechnology is a tangible step towards a waste-free sustainable society.
 Yafei Shen | Yafei Shen received an MSc degree (2012) in environmental engineering from Shanghai Jiaotong University (China). Currently, he is finishing his PhD degree at the Tokyo Institute of Technology (Japan), supported by the China Scholarship Council (CSC). He is also an associate editor for the Journal of Ecology and Environmental Sciences. His research interests focus on solid waste management, bioresource technology, bio-oil (e.g., algae oil) and tar catalytic reforming. He has published more than 20 academic papers in these fields. |
1. Introduction
Natural photosynthesis is the process in which sunlight is captured and converted into the energy of chemical bonds of organic molecules that are the building blocks of all living organisms, oil, gas and coal. These fossil fuels, which are the products of photosynthetic activity millions of years ago, could provide the energy to power our technologies, heat our homes and produce a wide range of chemicals and materials that support our life. Because of the ever-growing utilization of fossil fuels, we are faced with a severe problem of increasing levels of CO2 and other greenhouse gases in the atmosphere with implications toward global climate change.Photosynthesis as a successful energy generation and storage system is derived from the fact that the raw materials and power required for biomass synthesis are available in almost unlimited amounts, i.e. sunlight, water and CO2. The core process of photosynthesis is water splitting by sunlight into oxygen and hydrogen equivalents. Oxygen is released into the atmosphere, where it is available for living organisms to breathe and for burning fuels that drive our technologies. Hydrogen equivalents are used to reduce CO2 to sugars and other organic molecules. When fossil fuels, biomass and other biofuels are burned to release energy, the ‘hydrogen’ stored in these organic molecules is simply combined with atmospheric oxygen to form water. Similarly, energy is also released from organic molecules constituting our food when they are metabolized within our bodies during respiration. Thus, in the biological world, photosynthesis brings about the splitting of water into oxygen and hydrogen, whereas respiration is the reverse, combining oxygen and hydrogen in a carefully controlled and highly efficient manner to produce metabolic energy. From an energetic viewpoint, the synthesis of organic molecules implies a way of storing hydrogen and solar energy in the form of chemical bonds.1,2
This article comprehensively reviews the current progresses on green biofuels production from algae and mainly consists of four parts. The first part describes energy utilization along with the CO2 problem within the coming decades and discusses the possible contributions from photosynthetic biofuels based on the successful principles of photosynthesis. The global energy situation, CO2 and solar energy capture, and photosynthetic biofuels are also presented. In particular, it emphasizes the potential of exploiting the vast amounts of solar energy available to produce biofuels via an algae photosynthetic reaction combining advanced technologies. The second part describes the current barriers and challenges of biofuels production from algal biomass, including the new technologies for cultivation, harvesting and processing. The third part discusses the production of main biofuels (i.e., biohydrogen, biodiesel and bioethanol) from algal biomass. In addition, the integration of biodiesel and bioethanol production in biorefinery approaches have been presented for a better understanding of microalgae biofuel production and for future research and commercialization. Ultimately, the integrated algal systems for wastewater treatment and bioremediation to capture carbon (C), nitrogen (N) and phosphorus (P) from specialty industrial, municipal and agriculture wastes are introduced. To bring more profits, value added biofuels and chemicals can be developed by sustainable and applicable methods.
1.1. Global energy consumption and demands
Currently the global annual energy consumption rate is about 16.3 TW 3 with the USA and the extended EU each representing about 40%. In the future, this value will increase due to industrialization in underdeveloped and developing countries coupled with the increase in world population. Based on the current projections, the global annual energy consumption will reach 20 TW or even more by 2030, will double by 2050 and triple by the end of the century.4–6 About 85% of the total global energy consumed at present comes from burning fossil fuels with the proportion approaching 90% for developed countries. Oil, gas and coal contribute approximately equally to this demand. The remaining sources of energy are hydroelectric, nuclear, biomass and renewable, which includes solar, wind, tide and wave. At present, the utilization of biomass plays a dominant role in underdeveloped regions, such as Africa, where woody biomass and other organic matters are used as fuels.The low level contribution of non-fossil fuels to present-day global energy demand reflects the readily available resources of oil, gas and coal. Even when oil reserves become limited, large reservoirs of gas (including from shale) and, particularly, coal will remain to exploit.7 Therefore, globally, the problem for the immediate future is not a limitation of fossil fuel reserves but the consequences of its combustion. If the entire fossil fuel reserve is burnt, CO2 levels would rise to values equivalent to those that existed on our planet long before humankind evolved.8 Despite this, it is certain that fossil fuels will continue to be a major source of energy for some years to come, but it is vital that they should be used in such a manner that minimizes CO2 release into the atmosphere. Technologies for CO2 sequestration have been developed.9 With these technologies, efficiency of energy use and supplementation is improved whenever possible from non-fossil fuel sources. With this background, we must also strive to develop new technologies based on principles that are yet to be revealed from basic studies and in particular those that focus on using enormous amount of energy available to us as solar radiation.10 The sun provides solar energy to our planet on an annual basis at a rate of 1 × 105 TW. Therefore, the energy from 1 h of sunlight is equivalent to all the energy humankind currently uses in a year. Existing technologies can capture sunlight and produce electricity, and the efficiency and robustness of these photovoltaic systems is improving daily.11–13 Compared with the present-day cost of fossil fuels, photovoltaic systems represent an expensive method of generating electricity because of high construction costs. With time, these costs will decrease relative to the cost of fossil fuel. Moreover, a combination of the principles of photovoltaic systems, especially those using cheap organic or inorganic materials, with concepts derived from natural photosynthetic systems may provide a long-term solution via artificial photosynthesis technology.6,10
1.2. Carbon dioxide (CO2) and solar energy bio-capture
Since 1850, atmospheric CO2 levels, which were stable between 200 and 280 ppm for the past 4 × 105 years,14 have risen sharply to 370 ppm.15 Although the increased atmospheric CO2 level is now widely accepted to be a major contributor to global warming, its potential effects are only beginning to be understood. For example, recent high profile reports indicate that atmospheric CO2 levels of 450 ppm are likely to result in severe and probably irreversible coral reef damage.16 At levels of 550 ppm, the melting of the West Antarctic ice sheet will cause a 4–6 m rise in sea level16 and the extinction of about 24% of plant and animal species.17 A level of 650 ppm has been predicted to result in the disruption of thermohaline circulation (e.g., switching off the Gulf Stream), major local climate changes16 and the extinction of 35% of plant and animal species.17 More recent global climate change models18 suggest that the effects may be even more pronounced than previously predicted, emphasizing the importance of stabilizing levels as close to 450 ppm as possible and preferably below.15,16,19 However, it appears highly unlikely that CO2 levels will be maintained below this target due to the high CO2 emission levels and the long residence time of CO2 in the atmosphere. Hoffert and colleagues reported that about 11 TW CO2-emission-free fuel by 2025 was required to stabilize atmospheric CO2 levels at a level of 450 ppm.3 If Hoffert's predictions are correct, we are faced with the challenge of installing systems capable of producing energy free of CO2 emissions at a level almost equivalent to the total current global energy demand in 2000 (13 TW) within twenty years. This means that an abundance of zero-CO2 emission fuels (e.g., biohydrogen) is required urgently. Even biofuels, such as biodiesel and bioethanol, produce CO2, the difference will depend on the overall life cycle analysis, which takes carbon assimilation during feedstock production into account.In the current society, the development of zero-CO2 emission fuels is one of the greatest energy challenges for two urgent reasons. The first one is the rapid depletion of oil reserves, which requires the development of replacement fuels and infrastructure. Second, future fuels will increasingly have to be free of CO2 emissions because fossil fuel combustion causes anthropogenic CO2 emissions that exacerbate global warming. The constraints of global warming clearly indicate that the implementation of clean fuel technologies must take place much more quickly. The non-CO2 emitting energy options currently considered to be the most viable include nuclear power, coal-fired power stations coupled to anticipated CO2 sequestration systems, and renewable energy sources such as solar, geothermal, wind and hydroelectric. Of these, only renewable energy sources can sustain long term supplies and energy security (millennia) due to their borderless distribution. The promise of clean energy by nuclear fusion remains inaccessible. Among the renewable resources, incident solar energy is by far the largest (1.78 × 105 TW per year)20 and is capable of supplying 1.35 × 104 times the total global energy demand (13 TW per year in 2000). However, solar energy capture is both expensive and inefficient.
Nearly all life on earth captures solar energy and converts it into chemical energy and biopolymers by photoautotrophic organisms. Many organisms have developed complex molecular machinery for converting efficiently sunlight into chemical energy over the past 3 billion years, but man-made technologies capable of this conversion do not exist. Chlorophyll photochemistry within photosystem II (PSII) drives the water-splitting reaction efficiently at room temperature, in contrast with the thermal dissociation reaction that requires a temperature of ca. 1550 K. The high-resolution structure of PSII, particularly the structure of its Mn4Ca cluster,21–24 has successfully provided an invaluable blueprint for designing solar powered biotechnologies for the future. Combining this knowledge with new molecular genetic tools, fully sequenced genomes, and physiological processes of oxygenic phototrophs, researchers have been strongly inspired to develop new biotechnological strategies to produce renewable CO2-neutral energy from sunlight.25
An obvious target is manipulating photosynthesis to increase the initial capture of light energy, which at present is less than 2%. Recently, this approach has had some success using engineered genes from plants and photosynthetic bacteria. For example, ribulose-1,5-bisphosphate carboxylase-oxygenase (RuBisCO), the plant enzyme that converts CO2 to organic carbon by carboxylation during photosynthesis, also performs a competing, less efficient oxygenation reaction. When an inorganic carbon transporter gene from Cyanobacteria is expressed in plants, the more efficient carbon fixing photosynthetic reaction of RuBisCO is favored. In another approach, the cyanobacterial versions of two rate-limiting enzymes in the chloroplast's carbon-fixing ‘dark reaction’ were overexpressed in tobacco, resulting in an elevated rate of photosynthesis and increased plant dry weight.26 The manipulation of genes involved in nitrogen metabolism has also been a successful approach to increasing biomass.27,28
1.3. Photosynthetic biofuels
Most life-cycle studies have found that replacing gasoline with ethanol modestly reduces greenhouse gas emissions if made from corn and substantial reductions are observed if made from cellulose or sugarcane.29–46 These studies compare emissions from the separate steps of growing or mining the feedstocks (e.g., corn or crude oil) and processing them into the transportation fuels. Corn and cellulosic ethanol emissions exceed or match those from fossil fuels and therefore produce no greenhouse benefits. However, because growing biofuel feedstocks removes CO2 from the atmosphere, biofuels can in theory reduce greenhouse gas emissions relative to fossil fuels. Studies assign biofuels a credit for this sequestration effect, which is called the feedstock carbon uptake credit. It is typically large enough that overall greenhouse gas emissions from biofuels are lower than those from fossil fuels, which do not receive such a credit because they use carbon from the ground. It is our belief that the next generational change in the use of bioresources will come from a total integration of innovative plant resources, synthesis of biomaterials, and generation of biofuels and biopower. The premise of photosynthesis for the direct generation of fuels (Photosynthetic Biofuels) is that a single organism can serve both as a photo-catalyst and a producer of ready-made fuel. This concept is exemplified in the schematic of Fig. 1, where H2O, sunlight and CO2 are inputs and O2, biomass, fuels (H2, hydrocarbons) and chemicals are outputs. In this model, the conversion of solar-to-chemical energy, and biohydrogen, hydrocarbons, or other chemicals occurs within a single cell, possibly involving the photosynthetic apparatus and the adjacent cellular metabolism.47–49 | ||
| Fig. 1 Schematic depicting the concept of ‘Photosynthetic Biofuels’, where a single organism converts, via the process of oxygenic photosynthesis, H2O and CO2 into biomass and O2. Alternatively, photosynthate can be directed toward the generation of fuels and chemicals. Oxygen is a by-product of photosynthesis.47 | ||
So far, biofuels production from plants or algae photosynthesis has focused on the closure of the carbon cycle but not the nitrogen cycle. Plant-based and algae-based biofuels require the application of nitrogen fertilizer produced from the Haber–Bosch process. The reduced nitrogen is assimilated by the plant or algae to produce proteins and nucleic acids, which are not utilized for fuel production. Instead, the high-nitrogen containing residuals are used mainly as animal feed, and eventually result in the dispersion of reduced nitrogen on earth, which increases the production of nitrous oxide (N2O), a greenhouse gas 300 times worse than CO2.50 Feeding biofuel production residues to animals is currently economically attractive and may offset the energy and environmental cost of feed production, however, this is not a scalable solution if biofuels are to replace the majority of the liquid fuel used today. Recycling ammonia from protein-rich residuals as a fertilizer for photosynthetic feedstocks can close the nitrogen cycle. Corn ethanol, algal biodiesel, and other traditional feedstocks (Fig. 2A) do not utilize proteins and thus the reduced nitrogen is lost from the biofuel production cycle.51 Only the utilization of protein in a controlled manner will allow the recycling of ammonia. Fig. 2B shows the conceptual scheme for closed carbon and nitrogen cycles to optimize biofuel production. This concept can be practically implemented in both plant (Fig. 3A) and algal (Fig. 3B) biofuel production processes to recycle nitrogen fertilizer.51
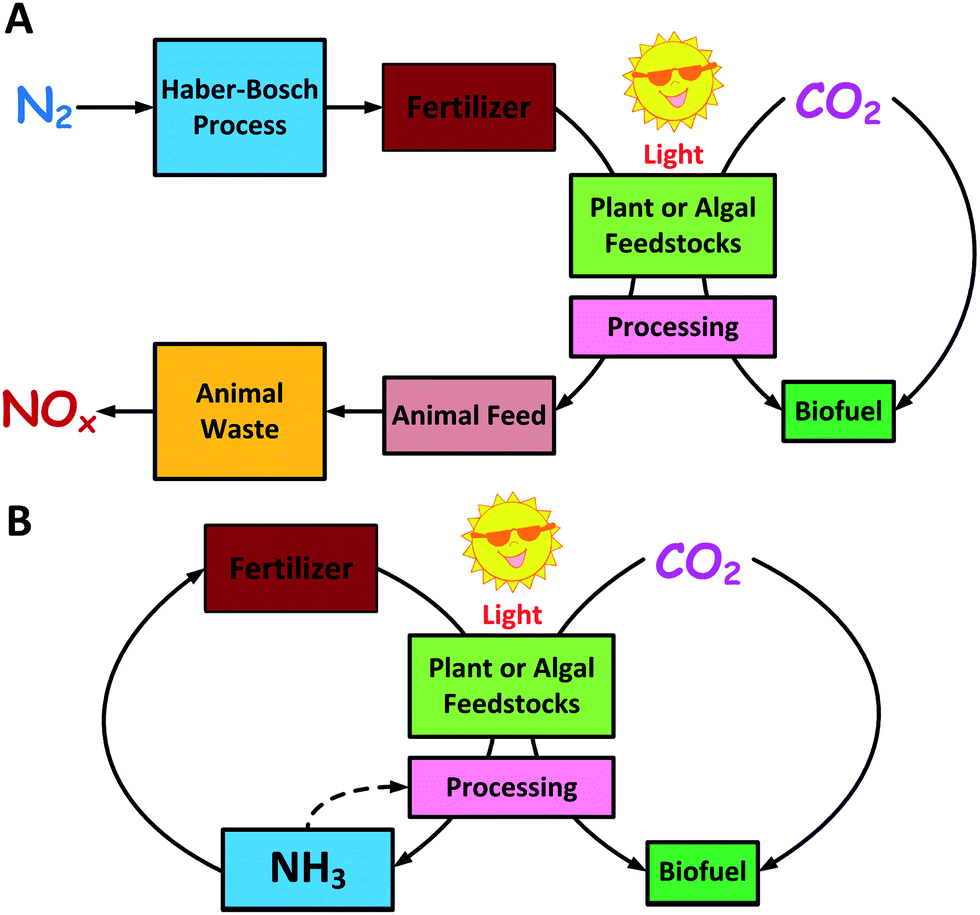 | ||
| Fig. 2 Carbon and nitrogen cycles in biofuel production. (a) Traditional biofuel production from plant or algal feedstocks closes the carbon cycle but imbalances the global nitrogen flux. Nitrogen is fixed through the Habor–Bosch process to synthesize fertilizer, which is assimilated to proteins in biomass. The nitrogen-rich residual is commonly sold as an animal feed by-product and leads to NOx emissions from animal wastes. (b) Utilization of proteins for fuel production can close both the carbon and nitrogen cycles. Protein conversion releases ammonia as a by-product. This ammonia may be reapplied as a fertilizer or nitrogen source for fermentation. | ||
 | ||
| Fig. 3 (A) Conceptual process for biofuels from plant biomass can recycle fertilizer when protein residual is utilized for ammonia recycling and fuel production; (B) biofuel production from protein-rich algae also releases ammonia that can be directly reapplied for subsequent algae growth. | ||
Because of the low photosynthetic efficiency and the competition of energy plants and food plants for agricultural land, some researchers suggested that it is unreasonable to grow plants for biofuel production.52–54 The main reason is that the growth of such energy plants will undoubtedly lead to an increase in food prices. Most prior studies reported that substituting biofuels for gasoline will reduce greenhouse gas emissions because biofuels sequester carbon through the growth of the feedstock. These analyses have failed to account for the carbon emissions that occur as farmers worldwide respond to higher prices and convert forest and grassland to new cropland to replace the grain (or cropland) to biofuels.53 Converting biomass into valuable building blocks for chemical syntheses may be the best choice. Compared with biofuel production, available biomass, instead of fossil fuels, is more preferable to be used for generating electricity. The saved fossil fuels could be used for transportation purposes. Clearing rainforests in the tropics and converting them into oil palm plantations is highly dangerous because the underlying layers of peat are oxidized and much more CO2 is released by the oxidation of organic soil material than can be fixed by oil palms. The rainforests play an important role in the climate and constitute a valuable resource for novel compounds and for drug discovery. With respect to carbon footprint, it will be much better to reforest the land used for growing energy plants because at a 1% photosynthetic efficiency, growing trees would fix around 2.7 kg m−2 of CO2, whereas biofuels produced with a net efficiency of 0.1% would only replace fossil fuels, which release about 0.31 kg m−2 CO2 upon combustion.54
2. Algae photosynthesis for biofuels
2.1. Photosynthesis
Photosynthesis is the process of converting light energy into organic molecules, which are mainly composed of carbohydrates, CH2O. Globally, photosynthetic efficiency is considerably lower than for agricultural and energy crops or algal cultures growing under optimal conditions because of seasonal changes and large portions of land and oceans, which do not sustain higher photosynthetic activity.55 Thus, the rate of energy storage averaged over a year by photosynthesis is 100 TW, representing just 0.1% conversion considering that solar energy is reaching our planet at a rate of 1 × 105 TW over the same period of time. This energy is mainly stored in wood and fibers of terrestrial trees and plants. A similar amount of photosynthetic activity occurs in the oceans but the fixed carbon is rapidly recycled in the food chain.56 Therefore, the global photosynthetic efficiency is about 0.2% with only half being stored as biomass (i.e., 0.1%). Terrestrial biomass was the major source of energy for humankind prior to the exploitation of fossil fuels. Therefore, it is not surprising that there is a growing interest in returning to the use of biofuels as an alternative to fossil fuels because of their CO2 neutral characteristics. Nevertheless, the scale required for satisfying the global energy requirement is far from attainable because of competition with large-scale food production and general land use required to sustain a global population of seven billion.Although it is possible to engineer plants and other photosynthetic organisms (i.e., algae) as energy-converting ‘machines’ and ‘chemical factories’, the overall efficiency of solar energy conversion will rarely exceed 1% and will usually be much less. Therefore, this approach can make only a minor contribution to our future energy requirements. However, the efficiencies of early photochemical and chemical reactions of photosynthesis, which are not directly involved in biomass production, are significantly higher. As a result, there are alternative and complementary approaches for using solar energy. It may develop a highly efficient, artificial, molecular-based, solar-energy-converting technology that exploits the principles of the ‘front-end’ of natural photosynthesis. Indeed, our knowledge of the natural process is to provide a blueprint for the design. An assembly of such ‘artificial photosynthetic’ devices can be described as follows. The process is based on the light-driven water-splitting reaction that occurs in the PSII of plants, algae and cyanobacteria (Fig. 4). First, solar energy is absorbed by chlorophyll and other pigments, which is then transferred efficiently to the PSII reaction center where charge separation occurs. This initial conversion of light energy into electrochemical potential occurs in the PSII reaction center with a maximum thermodynamic efficiency of 70% and generates a radical pair state P680˙+Pheo˙, where P680 is a chlorophyll a molecule, and Pheo is a pheophytin a molecule. The redox potential of P680˙+ is highly oxidized (about +1.2 V), while that of Pheo˙ is about 20.5 V. The latter is sufficiently negative because it can drive the formation of hydrogen. Instead, the reducing equivalent is passed along an electron transport chain to PSI, where it is excited by energy of a second ‘red’ photon absorbed by a chlorophyll molecule, known as P700, to lift it to a reducing potential of 21 V or even more. In this manner, sufficient energy is accumulated to drive CO2 fixation, which not only requires the generation of the reduced hydrogen carrier, i.e., nicotinamide adenine dinucleotide phosphate (NADPH), but also the energy-rich molecule adenosine triphosphate (ATP) formed by energy during electron transfer releasing from PSII to PSI in the form of an electrochemical potential gradient of protons.
 | ||
| Fig. 4 Simplified scheme of the light reactions of photosynthesis. | ||
2.2. Algae photosynthesis and CO2 biomitigation
Algae are recognized as one of the oldest life-forms57 and are present in all existing earth ecosystems, representing a diverse species living in a wide range of environmental conditions.58 They are primitive plants (thallophytes), i.e., lacking roots, stems and leaves, have no sterile covering of cells around their reproductive cells and have chlorophyll a as their primary photosynthetic pigment.59 Under natural growth conditions, phototrophic algae absorb sunlight and assimilate CO2 from air and nutrients derived from the aquatic habitat.60 The term ‘microalgae’ is not a biological but rather a practical description and its scope may differ depending on the context and the author. In its widest definition, microalgae are unicellular, photosynthetic microorganisms from several related branches of the tree of life, comprising, for example, prokaryotic cyanobacteria, eukaryotic green algae, red algae and heterokonts (e.g., brown algae and diatoms).58,61 Microalgae can produce large amounts of lipids, proteins and carbohydrates over short periods of time. These products can be processed into both biofuels and valuable co-products.60 However, the production of lipids, proteins and carbohydrates may be limited by available sunlight due to diurnal cycles and the seasonal variations; thereby limiting the viability of commercial production to areas with high solar radiation.62 Microalgae can fix CO2 from three different sources, viz. atmosphere, discharge gases and soluble carbonates.63 Under natural growth conditions, microalgae can assimilate CO2 from the air by tolerating and utilizing substantially higher levels of CO2 (up to 1.5 × 105 ppmv).64 Therefore, in common production units, CO2 is fed into the algae growth media either from external sources, such as power plants,65,66 or in the form of soluble carbonates such as Na2CO3 and NaHCO3.67,68 Other required inorganic nutrients for algae production include nitrogen, phosphorus and silicon.69 Algal cells are veritable miniature biochemical factories, and appear more photo-synthetically efficient than terrestrial plants because they are very efficient CO2 fixers. The ability of algae to fix CO2 has been proposed as a method of removing CO2 from flue gases to reduce greenhouse gas emissions from power plants. Many algal cells have been found to be exceedingly enriched with oil globules, which could be converted into biodiesel.70 Three distinct algae production mechanisms, namely, photoautotrophic, heterotrophic and mixotrophic are in use, all of which follow the natural growth processes. Photoautotrophic production is autotrophic photosynthesis, and heterotrophic production requires organic substances (i.e., glucose) to stimulate growth, while some algae strains can combine autotrophic photosynthesis and heterotrophic assimilation of organic compounds in a mixotrophic process.60 Many microalgae strains have a high lipid content (20–50% dry weight), which can be enhanced by optimizing the growth determining factors.71,72Most of the current research and developmental efforts are focused on microalgae due to their high growth rate and oil content. Algae contain oils, sugars, and functional bioactive compounds that can be used for commercial products. Recently, special attention has been given to cultivate microalgae as an energy crop with the aim of replacing traditional oil crops for biodiesel and bio-oil production. Algae have the potential to produce up to ten times more oil per acre than traditional biofuel crops such as oil palm. They can survive where agricultural crops cannot such as in salt water and on marginal land. They thrive on a diet of waste CO2 and nutrients in agricultural run-off and municipal wastewater. In addition to fuels, valuable co-products, such as biopolymers, proteins and animal feed, can be produced during the process. The concept of using algae to produce fuel was first discussed more than 50 years ago and a concerted effort began with the oil crisis in the 1970s.69 From 1978 to 1996, the US Department of Energy (DOE) devoted $25 million to algal fuels research in its aquatic species program at the National Renewable Energy Lab (NREL) in Golden, Colorado. The program yielded important advances that set the stage for algal biofuel research today.73
In the 1980s and 1990s, researchers tried various approaches. They grew algae in outdoor open ponds and enclosed photo-bioreactor tanks, experimented with breeding, fed algae smokestack CO2 emissions to boost their growth, and tested species that can tolerate extreme salt and pH environments. The first genetic transformation of microalgae was reported 1994. A few years later, scientists successfully isolated and characterized the first algal genes that express enzymes that were considered to enhance oil production. From 1990 to 2000, the Japanese government funded algae research through an initiative at the Research Institute of Innovative Technology for the Earth (Kyoto). The program focused on CO2 fixation and improved algal growth with concentrated mirrors that collect light. These approaches yielded some success and many are still the focus of scientists today, however, none have been proven to be economical on a large scale. The DOE program closed partly in 1996 because algal systems could not compete with the cheap crude oil of the late 1990s. The NREL-Chevron partnership started in 2007 and concluded in 2011, many efforts have been ongoing for the revived algae research program. Similar to all photosynthetic organisms, with a little water, a few nutrients and CO2, microalgae-pond scum use energy from the sun to grow. With just these inputs, they can easily double their population in a day. Faced with stresses, such as nutrient deprivation, algae put their energy into storage often in the form of natural oils such as neutral lipids or triglycerides and their growth diminishes. Similar to the oils from crops, such as soybeans, jatropha and oil palm, algal oil can be extracted from the organisms and refined into biodiesel by transesterification with short-chain alcohols (i.e., methanol) or by esterification of fatty acids.73 Algae can also be synthesized into other fuel products, such as hydrogen, ethanol and long-chain hydrocarbons, that resemble crude-like oil. Microalgal H2 is the direct product of the light reactions of photosynthesis. To bypass the H2 storage problems, an alternative approach would be to enable and harvest biofuel products from the carbon reactions of photosynthesis. Of particular interest is the process of generating and accumulating hydrocarbons via the fatty acid or terpenoid biosynthetic pathways.74–77 Hydrocarbons can be viewed as a biological method of storing hydrogen (Fig. 5).
 | ||
| Fig. 5 Photosynthetic products generated from the light and carbon reactions of photosynthesis. | ||
Vegetable and animal oils have long served as important raw materials for a number of applications, including surfactants, lubricants, polymers and foodstuffs.78 The primary precursors for these products are mono-, di- and poly-functional linear alkyl alcohols, aldehydes and acids are derived from the oxidative or reductive functionalization of acyl lipids and fatty acids.79–81 These modifications generally occur at either the carboxyl or olefinic moieties on the lipid, and the resulting products thus depend on both the tail length and the degree of unsaturation of the lipid precursor.82 Algal lipids are very similar to many plant lipids, with the notable exception that algal lipids are more likely to contain fatty acid components having higher degrees of unsaturation.69,83 Fig. 6 presents the values of both tail length and unsaturation for several representative algae and plant crops.78,84–87 It can be observed that many plants and algal crops have an average tail length in the 17/18 carbon range. In addition, highly unsaturated lipids in algae occur more frequently in polar lipid fractions, specifically phospholipids.88 Depending on species and growth conditions, phospholipids can compose anywhere from 8–47% of the total fraction of algal oil.89 In contrast, soy oil contains only 2–3% phospholipids.90 Because of the presence of the phosphate moiety, these lipids complicate many transesterification, reduction and combustion processes,91,92 and are therefore not desirable for biodiesel production without pre-treatment.
 | ||
| Fig. 6 Lipid compositions of selected algae and plant crops; the circle size corresponds to the average degree of unsaturation per lipid tail.78,84–87 | ||
Most of the current biofuel production is from the fermentation of sugar produced from grains by conventional yeast strains or transesterification by acid/alkali or enzyme based catalysts. The first generation of biofuel production is considered to have negative impacts on food security and possess a controversial energy balance.93 Second generation biofuels involve the biological processing of lignocellulosic biomass to overcome the fuel vs. food dilemma.94 Both 3rd and 4th generation biofuels use photosynthetic microorganisms to create renewable fuels: the former is basically the processing of algae biomass for biofuel production, while the latter is regarding the metabolic engineering of algae for producing biofuels from oxygenic photosynthetic organisms (Fig. 7). Algae metabolic engineering forms the basis for 4th generation biofuel production. It utilizes recombinant DNA and other biological and bioengineering techniques for the directed modification of cellular metabolism and properties through the introduction, deletion, or modification of algal metabolic networks to create or enhance biofuel production.95,96
 | ||
| Fig. 7 Four generations of biofuel production: from agricultural products to algae. | ||
Additionally, algae are more productive than plants. Under suitable culture conditions, the oil lipid productivity of microalgae can considerably exceed that of vascular plants.97,98 For example, the median value of the maximum specific growth rate of microalgal species is approximately 1 per day whereas for higher plants it is 0.1 per day or less.99 Each algal cell is photosynthetically active whereas only a fraction of plant biomass is engaged in photosynthesis. Each algal cell can absorb nutrients directly from its surroundings such that algae do not have to rely on energy-consuming, long-distance transport of nutrients via roots and stem. In addition to light, photosynthesis requires CO2. In plants, photosynthetic tissue can access CO2 only through pores known as stomata. These pores are not always open and CO2 must move through them against a flow of water vapor. The CO2 diffusion pathway from the surface of the photosynthetic tissue to a photosynthesizing cell is much longer in plants than in microalgae and increases with increasing the thickness of the photosynthetic structure.99,100 Therefore, algae can access CO2 more easily than vascular plants and this contributes to the relatively rapid growth. Because of their high solubility in water, the equilibrium concentration of CO2 in an algal suspension is greater than in the atmosphere above the suspension. Water enriching CO2 effectively is essential for photosynthesis. This also improves algal productivity relative to plants. Furthermore, because of a short life-cycle, algal biomass can be harvested daily or hourly, whereas plant biomass typically remains in the field for much longer. Unfortunately, because of the low productivity of plants, existing plant-derived biofuels cannot displace petroleum-based transport fuels to a significant extent. This severe limitation can only be overcome with a newer biofuels such as algae-based fuels. Unlike the existing crop-derived biofuels, algal fuels can be produced without encroaching on cropland and further deforestation. The production of algal biofuels need not reduce the supply of food, feed, other agricultural products and freshwater.97,98
The production of some existing biofuels demands unsustainable inputs of nitrogenous fertilizers, which are produced from fossil fuels and require huge inputs of energy for production.100,101 Plant-symbiotic bacteria, algae and other photosynthetic microorganisms can naturally convert atmospheric nitrogen to a form that can be used by life-forms, however, most crop plants and microalgae being considered for producing biofuels are not capable of this. Therefore, engineering plants and algae for nitrogen fixation capability is important for the sustainable production of biofuels. Production of all kinds of biofuels can be improved substantially by genetic and metabolic engineering,97,102–112 bioprocess engineering,113–115 the use of extremophilic species,116 and in other ways.117 The future of biofuels is intertwined with genetic and metabolic engineering. No form of renewable energy can fuel infinite growth and, therefore, society will have to learn to live within limits, including limits on population. Increasing the efficiency of energy use is essential and needs to be achieved without changing the lifestyle that we are accustomed to in the developed world. Within the constraints of sustainability, all humanity must attain an equitable quality of life. Algal biofuels have a clear potential for contributing to environmental, social and economic sustainability.118
Photosynthesis is the fundamental system required for all potential bioenergy surrogates production from photosynthetic microorganisms. However, it is a relatively low-efficiency process in terms of energy conversion when compared to the downstream synthesis of targeted products. More than 90% of the photon energy delivered to a given photosynthetic footprint can be dissipated as heat or fluorescence, and current estimates for realistic photosynthetic conversion efficiency are around 6% of total incident light energy.119–121 Maximizing photosynthetic potential is one of the most important and complex challenges in current efforts to exploit primary productivity for bioenergy applications (Fig. 8). Doan et al.122 reported that some researchers tried to directly exploit abundant algae or plants from marine or lakes for biofuels production. However, it should be noted that excessively utilizing algal biomass (i.e., marine algae) in existence for biofuels production might destroy the earth's aquatic ecosystem and change the global climate. However, according to the mechanisms of microalgae photosynthesis, algae could be rapidly grown and harvested in small-scale aquatic artificial systems under optimum conditions. Crucial components for the photosynthetic process are antenna proteins, which absorb light and transmit the resultant excitation energy between molecules to a reaction center. The efficiency of these electronic energy transfers has inspired considerable research on antenna proteins isolated from photosynthetic organisms to uncover the basic mechanisms.123–127 Intriguingly, recent works have documented128–130 that light-absorbing molecules in some photosynthetic proteins capture and transfer energy according to quantum-mechanical probability laws instead of classical laws at temperatures up to 180 K. This contrasts with the long-held view that long-range quantum coherence between molecules cannot be sustained in complex biological systems, even at low temperatures.131 Collini et al.132 and Richards et al.133 used two-dimensional photon echo spectroscopy measurements134–137 to study coherently wired and vibronic coupling light-harvesting in photosynthetic marine algae, respectively, at ambient temperature. These observations provide compelling evidence for quantum coherent sharing of electronic excitation across 5 nm-wide proteins under biologically relevant conditions, suggesting that distant molecules within the photosynthetic proteins are ‘wired’ together by quantum coherence for more efficient light-harvesting in cryptophyte marine algae.132
 | ||
| Fig. 8 Generic chloroplast of a green alga showing the placement of fuel-relevant primary metabolites and their integration into bioenergy production. The major components of photosynthesis and carbon fixation are also shown, including elements with the potential to be engineered for the optimization of these pathways, as described in the text (specifically BT, CA, FP, HYD, LHC, RuBisCO, SBPase, VAZ, water–water cycle). APX: ascorbate peroxidase, BT: bicarbonate transporter, CA: carbonic anhydrase, Cyt b6f: cytochrome b6f, FDX: ferredoxin, FFA: free fatty acids, FNR: ferredoxin–NADP+ reductase, FP: fluorescent protein, G3P: glyceraldehyde 3-phosphate, HCO3−: bicarbonate, HYD: hydrogenase, LHC: light-harvesting complex, PAR: photosynthetically active radiation, PC: plastocyanin, PS: photosystem, PQ pool: plastoquinone pool, SBPase: sedoheptulose-1,7-bisphosphatase, SOD: superoxide dismutase, SST: soluble sugar transporter, TAG: triacylglycerol, UV: ultraviolet light, VAZ: xanthophyll cycle.122 | ||
During the photosynthetic process, microalgae utilized CO2 from the atmosphere as a carbon source to grow and reproduce. Microalgae cells contain approximately 50% carbon, in which 1.8 kg CO2 are fixed by producing 1 kg of microalgal biomass.97 Hence, this method is recognized to be more environmentally friendly and technologically feasible to bio-mitigate CO2 compared to physicochemical adsorption or direct injection into deep ocean. However, low concentration of CO2 in the atmosphere (0.04%) with a poor mass transfer rate in water have resulted in the use of expensive air pump to deliver CO2 efficiently to microalgae rather than relying on natural diffusion from the atmosphere.138 On the other hand, flue gases from industry usually contain more than 15% (v/v) of CO2 (ref. 139) and therefore could be a prospective carbon source for microalgae. This is a win–win strategy in which air pollution from industry can be controlled through microalgae cultivation while the microalgae biomass can be used to produce biofuels.
Currently, extensive research has been focused toward identifying suitable microalgae strains that can grow under high concentration of CO2 while producing lipids for subsequent biodiesel production. The desired microalgae strains should have the following characteristics: (1) high growth rate and biomass productivity; (2) high tolerance to trace the amount of acidic components from flue gases such as NOx and SOx; (3) ability to sustain growth even under extreme culture conditions (e.g., high temperature of water due to direct introduction of flue gases). A few recent studies have reported that Chlorella sp., Scenedesmus sp., and Botryococcus braunii are among the microalgae strains that have shown promising results in mitigating CO2 emission with a typical CO2 consumption rate of 200–1300 mg L−1 d−1.140–144 Moreover, a pilot-scale system has been successfully developed to culture microalgae using industrial flue gases, and Scenedesmus obliquus was able to tolerate a high concentration of CO2 up to 12% (v/v) with an optimal removal efficiency of 67%.145 Moreover, supplying a high concentration of CO2 to microalgae can enhance the accumulation of polyunsaturated fatty acid in the microalgae cells.146 This is an encouraging observation as higher content of polyunsaturated acid tends to reduce the pour point of biodiesel produced making it feasible to be used in cold climate countries.
2.3. Algae cultivation and photobioreactors (PBRs)
One of the most understudied methods for CO2 mitigation is using biological processes (via microalgae) in a direct CO2 to biomass conversion from point source emissions of CO2 in engineered systems such as PBRs. Microalgal biofixation in PBRs has recently gained renewed interest as a promising strategy for CO2 mitigation. PBRs utilized for microalgal CO2 sequestration offers the principal advantages of increasing microalgae productivity because of controlled environmental conditions, optimized space/volume utilization, and more efficient use of expensive land. In fact, the photosynthetic solution when scaled up would present a far superior and sustainable solution for both environmental and economic considerations.147 Fig. 9 shows the integrated diagram of the applications of PBRs for waste CO2 capture and wastewater treatment by microalgae. The produced microalgal biomass can be used for biofuel production (e.g., biodiesel and methane) and other by-products such as animal feeds and polymers.148,149 | ||
| Fig. 9 Schematic diagram of microalgae photo-bioreactors applications for CO2 capture and biofuels production. | ||
In general, microalgae could be cultivated in open (pond) systems or closed systems. Considering all the limitations of the pond systems, most researchers have oriented their research toward the development of an unconventional method for microalgae culture, which should be fully closed and compact with a high surface-to-volume ratio and all the growth factors should be optimized. Closed reactors could be tubes, plates or bags made of plastics, glass or other transparent materials, in which the algae are supplied with light, nutrients and CO2.150,151 However, only a few of these designs can be practically used for the mass production of algae.152,153 For energy production, algal biomass is still expensive. On one hand, this price is governed by the perceived nutritional value of algal biomass that is mostly produced for animal feed and not for energetic usage. On the other hand, the price is caused by the low productivities of open ponds, high demands of auxiliary energy and high costs of classical PBRs designs. However, the problems are being addressed by engineering and science. Encouraging results have been obtained using new reactor geometries, optimized aeration and mixing strategies.154–157
An experimental helical-tubular PBR has been designed by Briassoulis et al.158 for controlled, continuous production of Nanochloropsis sp. Its main advantages include the combination of a large ratio of culture volume to surface area along with the optimized light penetration depth, easy control of temperature and contaminants, effective spatial distribution of fresh air and CO2, better CO2 transfer through extensive interfacing of the surfaces between fresh air and culture-liquid medium and a novel automated flow-through sensor providing continuous cell concentration monitoring. Henrard et al.159 evaluated the potential of semi-continuous cultivation of Cyanobium sp. in a closed tubular bioreactor by combining factors such as blend concentration, renewal rate, and sodium bicarbonate concentration. Cultivation was carried out in vertical tubular PBR for 2 L, in 57 d, at 30 °C, 3200 Lux, and a 12 h light/dark photoperiod. The maximum specific growth rate was observed to be 0.127 per day when the culture had a blend concentration of 1.0 g L−1, a renewal rate of 50%, and a sodium bicarbonate concentration of 1.0 g L−1. The maximum values of productivity 0.071 g L−1 d−1 and the number of cycles (10) were observed at a blend concentration of 1.0 g L−1, renewal rate of 30%, and bicarbonate concentration of 1.0 g L−1. These results showed the potential of semi-continuous cultivation of Cyanobium sp. in a closed tubular bioreactor by combining factors such as blend concentration, renewal rate, and sodium bicarbonate concentration.
The hydrodynamic and mass transfer characteristics of a flat-panel airlift PBR with a high light-path are more efficient than those reported elsewhere for tubular and other flat-plate PBRs, which opens the possibility of using PBRs with higher light paths than proposed so far.160 Janssen et al.152 studied light regime, photosynthetic efficiency, scale-up, and future prospects of an enclosed outdoor PBR. In this study, they showed that the productivity of PBRs is determined by the light regime inside the bioreactor. In addition to the light regime, oxygen accumulation and shear stress limit productivity in certain designs. In short light-path systems, high efficiencies of 10–20% based on photosynthetic activate radiation (PAR 400–700 nm) can be reached at high biomass concentrations [>5 kg m−3 (dry weight)]. However, it is demonstrated that these and other PBR designs are poorly scalable (maximal unit size 0.1–10 m3) and applicable for the cultivation of monocultures. Therefore, a new PBR design is proposed in which light capture is physically separated from photoautotrophic cultivation. This system can possibly be scaled to larger unit sizes, 10 to >100 m3, and the reactor liquid as a whole is mixed and aerated. It is deduced that high photosynthetic efficiencies of 15% on a PAR-basis can be achieved. Future designs from optical engineers should be used to collect, concentrate, and transport sunlight, followed by redistribution in a large-scale PBR. The co-operation project between The Norwegain Institute for Agricultural and Environmental research in Norway, Uppsala University in Sweden and IIT Kharagpur in India, the BioCO2 project (2008–2011), has designed, constructed and tested a flat panel, rocking PBR for algae cultivation (non-rocking mode) and hydrogen production (rocking mode). It consists of two glass plates fixed between an inner frame made of stainless steel and outer frames made of aluminum, an air bubbling tube and a tube designed for temperature regulation.161
2.4. Algae genetic and metabolic engineering
In recent years, new biotechnological approaches related to genome perturbation of microalgal cells to endow them with different properties are dramatically increasing. However, the full potential of genetic engineering of some microalgal species, particularly diploid diatoms, can be fully realized only if conventional breeding methods become firmly established, thereby allowing useful mutations to be easily combined.112,162 Significant advances in microalgal genomics have been achieved during the last decade.112,162–165 Expressed sequence tag databases have been established; nuclear, mitochondrial, and chloroplast genomes of several microalgae strains have been sequenced. Historically, the green algae Chlamydomonas reinhardtii has been the focus of molecular and genetic phycological research. Therefore, most of the tools developed for the expression of transgenes and gene knockdown are specific for this kind of species. Current genetic engineering pursuits are toward microalgae that are of greater interest in industrial applications and environmental conservation.162 To improve microalgal biomass or lipid production and CO2 capturing efficiency, several approaches have been developed.Up to now, efforts to increase the lipid content of microalgae have been mainly focused on the optimization of growth and induction conditions such as temperature, light, salinity and nutrient content/depletion, for instance,165–167 reported genetic modifications of microalgae to alter either lipid quantity or quality (i.e., composition) are still sparse. The main reason is probably the lack of a generally applicable transformation protocol for microalgae. Because microalgae are such a diverse group or organisms, it is not guaranteed that a method that works for one species can be applied to another one. For example, some species, such as D. Salina, do not have a rigid cell wall, whereas diatoms often have a very rigid silicate shell. This directly affects the method of gene transfer into the cell.168 Another problem is the limited range of available markers. Although auxotrophy markers are available for some species, such as C. reinhardtii, stable transformation of other species still has to rely on co-transformed genes conferring resistance to antibiotics. However, some substances routinely used in the transformation of plants, such as kanamycin and hygromycin, are sensitive to increased NaCl concentrations and cannot be used for strains requiring sea water. In addition, heterologous gene expression (e.g., the expression of genes not originating from the organisms) in microalgae suffers from the lack of available promoter sequences to control expression, and the possibility of codon usage bias. In summary, any protocol for the genetic transformation of a new microalgal strain (not necessarily a new species) has to be carefully modified to meet and overcome its specific requirements and limitations. Despite the obstacles described above, genetic modification is one of the main tools to study metabolic pathways in microalgae, and is strongly contributing to our knowledge regarding their biology. Metabolic engineering by genetic modification is expected to be one of the main steps that will lead to versatile, sustainable and economically viable biofuels from algae.96,111,166–174 As shown in Fig. 10, unicellular algae are capable of synthesizing a range of biofuels. Lipids and carbohydrates represent the main energy storage molecules in algae, and a broad understanding of primary metabolism is necessary to manipulate electron flux toward these products or H2 for bioenergy applications. The distinct metabolic processes that occur within algal organelles and the numerous enzyme isoforms present in a cell complicate these efforts.111,175–177
 | ||
| Fig. 10 Photosynthetic and glycolytic pathways in green algae related to biofuel and biohydrogen production. Simplified illustration of the pathways used for lipid, starch, and H2 production in Chlamydomonas reinhardtii.111 | ||
Research interest into microalgal lipid production for biofuels is at an all time high with a whole range of studies being conducted from growth optimizations178–180 to induced mutagenesis of microalgae to improve lipid yield.169–172 It can be envisaged that careful strain selection and improvements in microalgae for a variety of useful traits holds significant promise and can be compared with efforts in conventional agricultural crop breeding. The current bottlenecks for large-scale cultivation appear to be in harvesting/extraction processes as well as in cheap and energy efficient cultivation systems. The commercial production of biodiesel from algae depends on lipid productivity in industrial scale cultivation systems, production costs, and the energy ratio of production.181 Against each of these aspects, microalgae lipid production presents a mixed picture. A positive energy balance will require technological advances and highly optimized production systems. The mitigation of environmental impacts, and in particular water management, presents both challenges and opportunities, many of which can only be resolved at the local level. Existing cost estimates need to be improved, which require empirical data regarding the performance of systems designed specifically to produce biofuels. Currently, it appears that the sustainable production of biofuels from microalgae requires a leap of faith, but there are some grounds for optimism. Because of the diversity of algal species, it is highly likely that new applications and products will be found. As experience with algal cultivation increases, biofuels may have a role to play.182–184
3. Algae harvesting and processing for biofuels
3.1. Algae harvesting
A major challenge in the downstream processing of microalgae is separating the microalgae from their growth medium, i.e., the harvesting process. A high biomass concentration leads to the mutual shading of the microalgal cells and thus a reduction in productivity. Therefore, biomass concentrations in microalgal cultures are usually low, from 0.5 g L−1 in open pond reactors to about 5 g L−1 in PBRs. This implies that a large volume of water needs to be removed to harvest the biomass. Because of the small size of microalgal cells (2–20 μm) and their colloidal stability in suspension, harvesting by sedimentation or simple screening is not feasible, except perhaps for larger species such as Arthrospira. When microalgae are produced for high-value added products, harvesting is performed by centrifugation. Besides, due to lower costs, flocculation, electro-coagulation-flocculation and membrane filtration are recommended. However, centrifugation is very expensive and energy-intensive if biomass is to be used for low-value products, such as biofuels, because of the large volumes of culture medium that require processing. Finding an alternative technology that is capable of processing large volumes of culture medium at a minimal cost is essential for reducing the cost and increasing the scale of microalgal biomass production.60,185–187To realize large-scale production of microalgal biomass for low-value applications, new low-cost technologies are required to produce and process microalgae requiring the separation of a low amount of culture medium. Flocculation is considered to be one of the promising low-cost harvesting methods.188 Methods available for harvesting algae from broth include centrifugation, filtration, flocculation, and gravity sedimentation. The method chosen, to a great extent, depends on the final product and the processes subsequently used; some processes require the algae to be completely dewatered while others do not.189,190 The cost and energy demand for harvesting microalgae could be significantly reduced if the cells are pre-concentrated by flocculation.191,192 During flocculation, single cells form larger aggregates that can then be separated from the medium by simple gravity sedimentation. When flocculation is used for harvesting microalgae, it is a part of a two-step harvesting process. Flocculation is used as a first step to concentrate a dilute suspension of 0.5 g L−1 dry matter 20–100 times to form a slurry of 10–50 g L−1. Further dewatering using a mechanical method, such as centrifugation, is then required to obtain an algal paste with 25% dry matter content.193 The energy requirements for this final mechanical dewatering step are acceptable because the particles are relatively large and the volumes of water to be processed are small.187 The economics are very different when flocculation is used for harvesting microalgal biomass than when it is used for removing impurities from a liquid. In addition, contamination is a major issue because chemicals added to induce flocculation end up in the harvested biomass. These chemicals can interfere with the final applications of the biomass (i.e., food or feed) or with further processing of the biomass (e.g., lipid extraction).186 Flocculation could be achieved in several ways, which have been widely explored for microalgae harvesting in recent years. These approaches range from traditional flocculation methods that are widely used in other industries (e.g., chemical flocculation) to novel ideas based on the biology of microalgae (e.g., bioflocculation) and the utilization of emerging technologies (e.g., magnetic nanoparticles utilization).194
Recently, Rashid et al.205,206 used chitosan as a flocculant to harvest freshwater microalgae Chlorella vulgaris. In the chitosan-based microalgae harvesting process, bridging was the primary mechanism of flocculation. Chitosan is a promising choice due to its high molecular weight and charge density. It contains positively charged amino groups (NH3+ and NH2+), which have a tendency to adsorb negatively charged microorganisms, including microalgae.207 When chitosan co-exists with negatively charged algal cells in a solution, electrostatic repulsion between the cells decreases. The decrease in electrostatic repulsion reduces zeta-potential and promotes flocculation.208 If chitosan binds partly with microalgae cells, the empty cell surface attaches to another cell and forms a chain-like structure called bridging. At high flocculant concentration, microalgae cells are covered by cationic polymer leaving insufficient empty sites, generating a net positive charge.208 This positive charge also attaches with the surrounding negatively charged cells to form flocs. This phenomenon is called patching. Chitosan holds tremendous potential for high biomass recovery from microalgae culture. Low dose requirement and short settling time are the distinct advantages of chitosan over common flocculants. Microalgal culture can be concentrated up to 10 times at optimal pH (6.0) and flocculant dose (120 mg L−1 chitosan). Further studies should be carried out to explore the possible ways to reduce the chitosan dose for cost-effective microalgae harvesting. Farid et al.209 studied a nano-chitosan method for harvesting microalga Nannochloropsis sp. Nano-chitosan showed better biomass recovery. The dosage of chitosan consumption decreased from 100 to 60 mg L−1 and biomass recovery increased about 10% by using nano-chitosan. The best initial cell density was 665 × 106 cells per mL for minimum flocculant dosage consumption and minimum process cost. The presence of acetic acid in recycled water from harvesting showed an increase in microalgae growth. Using recycled water increases biomass concentration and at the same time has no treatment cost.
Lee et al.210–212 also utilized aminoclays having a high density of amino sites (–NH2) and water-soluble, transparent, and less ecotoxic effects in aqueous solution213 for the rapid harvesting of freshwater and marine microalgae. The aminoclays placed at the metal (i.e., Fe3+) center were synthesized by a sol-gel reaction with 3-amino-propyltriethoxysilane as a precursor, producing –(CH2)3NH2 organo-functional pendants, which are covalently-bonded onto cationic metals. Protonated amine groups in aqueous solution lead the efficient sedimentation (harvesting) of microalgal biomass within approximately 5 min and 120 min for fresh and marine species, respectively.210 Significantly, the aminoclays did not depend on microalgae species or media for microalgae harvesting. In particular, the harvesting efficiency (%) did not decrease in a wide pH region. The harvesting mechanism can be explained by the sweep flocculation of microalgae, which was confirmed by measuring the zeta potential of the aminoclay in aqueous solution and determined that the aminoclay surface is positively charged over a wide pH range. To reduce the cost of aminoclays and simplify the harvesting procedures, the membrane process using an aminoclay-coated cotton filter was employed for the treatment of 1 L-scale microalgae stocks. This was successfully performed with three recycles using the same aminoclay-coated cotton filter after removing the harvested microalgae. In conclusion, the aminoclay-based microalgae harvesting systems are a promising for reducing the cost of downstream processes in microalgae-based biorefinery.210
Recently, several studies have explored the use of magnetic nanoparticles to harvest microalgae. Magnetite (Fe2O3) nanoparticles may adsorb directly on the microalgal cells, upon which the cells can be separated from the medium by applying a magnetic field. Thus, this method combines flocculation and separation in a single process step.225,226 Magnetite nanoparticles appear to adsorb more easily on some microalgal species than on others.227 Adsorption can be improved by coating the nanoparticles with cationic polymers.228,229 An advantage of using magnetite nanoparticles for harvesting microalgae is that the nanoparticles can be recovered after harvesting and can be subsequently reused.225 Bejor et al.230 investigated the low cost harvesting of microalgal biomass from water using a physical method. Four fabric filters (stretch-cotton, polyester-linen, satin-polyester and silk) were used for microalgae harvesting using a filtration method. For three algae communities with a cell size of 2–20 μm, stretch-cotton filter showed a harvesting efficiency of 66–93%, followed by polyester-linen (54–90%), while satin-polyester and silk fabrics achieved the harvesting efficiencies of 43–71% and 27–75%, respectively. The research revealed that for wastewater generation of 1500 m3 d−1 and algae concentration of 200 mg L−1, microalgae harvesting cost per m2 per kg of algae per m3 would be ≤£0.15 using stretch cotton filter. Thus, fabric filters utilized for algae harvesting have been proven to be a cheap and reliable harvesting technique especially in areas where acquiring skilled labor is not feasible.
Bacteria or fungi can also induce the bioflocculation of microalgae. Some fungi, for instance, have positively charged hyphae that can interact with the negatively charged microalgal cell surface and cause flocculation.237,238 Specific consortia of bacteria can also induce the flocculation of microalgae.239,240 These flocculating fungi or bacteria can be cultivated separately or in combination with the microalgae. Cultivating bacteria or fungi in combination with microalgae requires a carbon source in the medium. In wastewater, a carbon source is usually present that allows the cocultivation of microalgae and bacteria. This results in a culture of mixed algal–bacterial flocs that can be easily harvested.241,242 The use of bacteria or fungi as a flocculating agent avoids chemical contamination of the biomass but results in microbiological contamination, which may also interfere with food or feed applications of microalgal biomass.243
The energy intensive harvesting of tiny microalgae cells (1–70 μm) from culture broth can account for at least 20–30% of the total costs of algal biomass production. Recently, Zhou et al.244 developed an alternative fungus pelletization assisted bioflocculation method for harvesting microalgae (Chlorella vulgaris UMN235) using a pellet-forming fungal strain (Aspergillus oryzae) isolated from municipal wastewater sludge. Under heterotrophic growth conditions, the key factors, including spore inoculums, organic carbon concentration in medium as well as pH variation had significantly positive effects on fungus–algae pellet formation. The process parameters of 1.2–104 spores per mL, 20 g L−1 glucose, and pH ranging from 4.0 to 5.0 were found to be optimal for efficient fungus–algae pellet formation. For autotrophic growth, when the pH of the culture broth was adjusted to 4.0–5.0 with the addition of organic carbon (10 g L−1 glucose), almost 100% harvesting efficiency of microalgae was obtained. Moreover, it was observed that the diameter and concentration of fungus–algae pellets were affected by shaker rotation. The novel harvesting technology might reduce the cost of microalgae harvesting and will have potential to be applied to all types of microalgae species as an alternative to other traditional harvesting methods. Lee et al.245 proved that bacteria play an important role in flocculation by increasing the floc size resulting in the sedimentation of microalgae. The collective presence of certain bacteria was the determining factor in the flocculation of C. vulgaris.
The membrane separation efficiency is considerably affected by fouling. It can be further explained that microorganisms accumulated on membrane surface or in membrane pores causes a decline in permeate flux.254 Many efforts have been made to understand and reduce fouling, including membrane surface modification and new membrane material development.255 Conventional polymeric materials membranes have been widely used in the filtration and concentration of microalgae.249,256–258 Rossignol et al.259 evaluated the performances of inorganic filtration membranes. Liu260 utilized a thin, porous metal sheet membrane to harvest microalgae, which exhibited high properties of membrane area packing density, chemical/thermal stability, mechanical strength, high permeability and low cost. Sun et al.261 evaluated several commercial MF and UF membranes for the filtration and concentration of Chlorella from dilute culture media. The results showed that permeate fluxes increased with the increase in feed solution temperature, and that the fluxes were probably limited by released extracellular polymeric substances (EPS) at higher temperatures. Moreover, MF and UF membranes showed similar flux in this work, indicating that pore size and porosity are not important for this application. This suggests that the permeate flux of different membranes is controlled by the fouling layer that acts as the membrane selective layer. The work also demonstrated that a membrane with hydrophilic surface shows very little fouling during algae harvesting.
To reduce fouling, Hwang et al.262 proposed a membrane technology by the surface-coating with a functional coating material, i.e., hydrophilic polyvinyl alcohol (PVA) polymer. The PVA coating caused the membrane surface to become more hydrophilic, which was confirmed by reduced contact angles up to 64% compared to the unmodified membranes. The surface-coated membrane was found to exhibit substantially enhanced performance: a maximum flux increase of 36% and an almost 100% recovery rate. It showed that the membrane performance could be simply improved (even to the level of economic feasibility) by applying a surface-active coating.
The enhancement of membrane shear-rates has long been recognized to be one of the most efficient factors for fouling control. It is implemented either by moving the fluid or the membrane. The membrane can be moved in a circular rotation, a torsional vibration or in vertical and horizontal oscillation systems.263,264 Application of a rotating disk system for algal harvesting showed that it almost doubled the membrane productivity compared to a reference cross-flow system, which is attributed to high shear-rates at the liquid-membrane interface.265,266 However, Ladner et al.267 found a very significant impact of enhanced shear on microalgal cells. The algal organic matter released from sheared microalgal cells caused increased membrane pore blocking. This phenomenon was not observed in other studies265,266 probably due to different types of microalgae (cell wall), type of pumps, filtration experimental designs (shorter time-frame), etc. Therefore, a process that would maintain a high shear-rate only at the liquid-membrane interface, and not in the entire bulk, would be beneficial to achieve an efficient filtration process. Bilad et al.268 investigated the effectiveness of submerged microfiltration to harvest both a marine diatom Phaeodactylum tricornutum and Chlorella vulgaris in a magnetically induced membrane vibrating (MMV) system. They assessed the filtration performance by conducting the improved flux step method (IFM), fed-batch concentration filtrations and membrane fouling autopsy using two lab-made membranes with different porosity (Fig. 11). The full-scale energy consumption was also estimated. Overall results suggested that the MMV offered a good fouling control and that the process was proven to be economically attractive. By combining the membrane filtration (15 × concentration) with centrifugation to achieve a final concentration of 25% w/v, the energy consumption to harvest P. tricornutum and C. vulgaris was as low as 0.84 and 0.77 kW h m−3, corresponding to 1.46 and 1.39 kW h kg−1 of the harvested biomass, respectively.
 | ||
| Fig. 11 Experimental set-up for (A) improved flux stepping filtration method (IFM) test in a total permeate recycle filtration mode, showing the parallel view of the narrow edges of the two vibrating membranes, and (B) fed-batch concentration filtration showing the set-up in a full surface view of the vibrating membranes.268 | ||
Development of an efficient flocculation technology for microalgae may yield major cost and energy savings in large-scale production. In general, chemical flocculation could result in the contamination of microalgal biomass because the use of natural polymers may minimize this problem. Alkaline flocculation promises to be a low-cost flocculation method but results in the contamination of the biomass, albeit with mineral precipitates with low toxicity. Bioflocculation by fungi or bacteria holds a potential feasibility when microalgae production is combined with wastewater treatment because wastewater can provide the necessary carbon source for flocculating microorganisms. Physical flocculation can avoid biomass contamination due to chemicals or microorganisms. Fundamental research into infochemicals that induce flocculation in microalgae are necessary because they may contribute to a highly controllable method for inducing flocculation that avoids contamination. The same holds true for approaches to induce flocculation through genetic modification. Further studies should examine the flocculation efficiency under specific conditions, and investigate how flocculation is affected by the properties of the microalgal cells or culture conditions, particularly interfered by organic matters in the culture medium. Cost evaluation should take account of the flocculation step and the entire production process.
3.2. Algae hydrothermal (HT) processing
Several processing approaches for biofuels from land-based biomass have been developed and partly commercialized. Nevertheless, algae also contain carbohydrates that can be converted by similar processes. In regards to biomass, not every process is suitable for application in an efficient and economical manner. Therefore, well-known processes have to be tested for algal biomass. In addition to this, microalgae are offering novel pathways of producing biofuels, which have to be taken into account.269 One of the economic and energetic drawbacks in the processing of microalgae is the dewatering stage because microalgae typically grow to a solid concentration of 1–5 g L−1.60 The challenges of concentrating and drying are energy intensive. Macroalgae can be harvested more easily due to their large size but their moisture content is still very high compared with terrestrial biomass.270 Microalgae biofuel is usually produced by the extraction of lipids and the subsequent transesterification to form biodiesel. Most common lipid extraction techniques require a dry feedstock before transesterification. This can account for as much as 25% of the energy contained in algae.271Hydrothermal (HT) processing avoids the step of drying because algae are treated as a slurry in hotcompressed water. Operating conditions depend on the desired product: at low temperatures, less than 200 °C, the process is referred to as HT carbonization (HTC) and predominantly produces a char; at intermediate temperatures of 200–375 °C, the process is known as HT liquefaction (HTL), primarily producing an oil; at the higher end of the temperature range, greater than 375 °C, the process is called HT gasification (HTG), predominantly producing a syngas. These HT processing routes are expected to generate a product with higher energy density. The char produced from HTC can be co-fired with coal or used as biochar for soil amendment,272 the biocrude from HTL can be upgraded into a variety of fuels and chemicals, while the syngas from HTG can be used for combustion or converted into hydrocarbons by either biological or catalytic processing, e.g., Fisher–Tropsch synthesis. Other than the abovementioned HT processes, there are some additional wet processing methods that have been used for algal biomass as wet extraction techniques offer a distinct advantage of energy requirement. For example, Levine et al. proposed the in situ lipid hydrolysis of wet algae followed by supercritical transesterification with ethanol.273 Alternatively, Patil et al. suggested the wet transesterification to fatty acid methyl esters in supercritical methanol.274 There are limited studies on the coliquefaction of algal biomass with coal or organic solvents to improve the yields and quality of biocrude.275,276 During the carbonization stage, carbon content is enhanced and the oxygen content and mineral matter is reduced; the gaseous product is low and a biochar is produced by carbonization reactions. During liquefaction, biomass is decomposed to smaller molecules, which are reactive and can repolymerize into oily compounds.277–280
The products from HTL consist of a biocrude fraction, a water fraction containing some polar organic compounds, a gaseous fraction and a solid residue fraction. Under the more severe conditions in HTG, the desired product is a syngas, consisting of varying amounts of H2, CO, CO2, CH4 and light hydrocarbons. The initial reaction steps are the same as liquefaction but more severe conditions lead to small fragments decomposing even further to lowmolecular weight gaseous compounds. At high temperatures (>500 °C) H2 production is favored, while CH4 production is favored at 350–500 °C, however, all these conversion pathways can be influenced with the use of catalysts.281–286 The high ionic product supports acid or basecatalyzed reactions and can act as an acid or base catalyst precursor because of the relatively high concentrations of H3O+ and OH− ions from the self dissociation of water.283 The advantage of this method is that additional acid or base catalysts can be avoided. The ion concentration can reach a maximum at 275 °C, which is the optimum temperature for acid or basecatalyzed reactions. Above 350 °C, the ionic product decreases rapidly by five orders of magnitude or more above 500 °C.284 Between 300 and 450 °C, the density at 30 MPa changes from a liquidlike 750 kg m−3 to a gaslike 150 kg m−3; however, phase change does not occur. The density change is directly associated with properties such as solvation power, degree of hydrogen bonding, polarity, dielectric strength, diffusivity and viscosity.287
Chemical reactions under hydrothermal conditions and in supercritical fluids can provide new, potentially cheaper paths to renewable fuels from wet algal biomass.288 The methods used to make large quantities of liquid fuels from algae involve extracting the oil with an organic solvent, such as hexane, and converting the oil into either biodiesel by catalyzed transesterification with alcohol or to green diesel by catalytic hydrotreating. Drying algae prior to extracting takes time, consumes energy, and is expensive. Producing fuel directly from wet algal biomass could improve the economics and environmental sustainability of algal biofuels. Thus, some alternative methods, such as hydrothermal and solvothermal processes, have been developed.289 HT processing is an energy efficient approach favoring the required reactions.290 Hot compressed water (e.g., 300 °C, 8.6 MPa) could readily dissolve organic compounds, and its elevated ion product (10−11 versus 10−14 for ambient water) could accelerate acid-catalyzed, hydrolytic decomposing biomacromolecules.290,291 Algal biomass contains macromolecular proteins, polysaccharides, and lipids, along with inorganic components. The lipid fraction is usually targeted for fuels but the protein and polysaccharide fractions also have heating value. Thus, the conversion of the entire biomass into fuels can lead to biocrude yields exceeding the lipid content of the algae, while a greater partition of the heating value originally present in the biomass into the final fuel products (Fig. 12). HT processing can also facilitate the reuse of nitrogen (N) and phosphorus (P) required for sustainable processing.289,292 Herein, Fig. 13 illustrates a photobioreactor for microalgae cultivation, where nutrients, water, light and CO2 are the only required inputs. A similar concept could be described for open pond cultivation or for macroalgae, where the cultivation layout could include growth in either closed tanks or in marine environments. More importantly, when the algal biomass is treated by the HT processing, some dewatering is still required. Lowcost dewatering has more challenges for microalgae than macroalgae but many processes are available such as flocculation described above.289
 | ||
| Fig. 12 Hydrothermal and supercritical fluid processing approaches for transformation of wet algal biomass into fuels and other products fractionate the biomass first or process the entire biomass first. | ||
 | ||
| Fig. 13 Integrated hydrothermal process with nutrient and CO2 recycling for algae photosynthesis. | ||
In summary, culturing microalgae for biofuels production could be combined with wastewater treatment to minimize the heavy dependency on inorganic nutrients. Apart from this, the incorporation of a baffled system in an open pond and closed-photobioreactor is recommended to enhance the mixing intensity between microalgae, nutrient sources and CO2 while reducing the energy input. In addition, effective harvesting and drying of microalgal biomass can be easily achieved through immobilization technology; however, extensive research is still required to strengthen this visionary strategy. For downstream processing, lipid extraction from microalgae presents a complicated task. Physical extraction methods, which are suitable for extracting oil from crops, are not efficient in extracting lipid from microalgae because the lipid is embedded within a layer of cell wall. Cell disruption methods, such as chemical or thermal extraction, are necessary to effectively recover the lipid. However, some of the cell disruption methods require large amounts of energy input that could lead to negative energy balance. In addition, it is noteworthy that the choice of cell disruption methods, chemical solvents and extraction conditions significantly rely on the choice of microalgae strains. In other words, no single method can give optimum lipid extraction for all the types of microalgae strains. Several breakthrough technologies, such as supercritical extraction/transesterification, in situ transesterification, hydrothermal processing and transesterification assisted with ultrasonication or microwave, are yet to be discovered to enhance microalgae biocrude production. Moreover, biodiesel derived from microalgae still would ideally be the main product. In addition, diversified biofuels (i.e., biohydrogen, bioethanol) production from microalgae is necessary to improve the overall energy balance. For instance, microalgal biomass after lipid extraction can be recycled for bioethanol production because a high concentration of carbohydrates remains in the biomass. Another potential biofuel derived from the microalgal biomass residue is bio-oil from pyrolysis or hydrothermal process. This is a win–win strategy for recycling the waste to produce another source of energy, which significantly amplifies the sustainability of microalgae biofuels. Nevertheless, bioethanol and bio-oil production from microalgae is still at the infancy stage and its real potential is yet to be discovered. In the next section, the main biofuels, such as biohydrogen, biodiesel, and bioethanol, derived from algal biomass will be presented in detail.
3.3. Biohydrogen
Uniquely, among the organisms of oxygenic photosynthesis, many green microalgae encode for the genes of hydrogen metabolism, including two [Fe–Fe] hydrogenases,293,294 and genes encoding proteins required for the [Fe–Fe] hydrogenase assembly.295,296 Hydrogen metabolism-related proteins in green microalgae are localized and are functional in the chloroplast such that the [Fe–Fe] hydrogenase can receive high potential energy electrons directly from reduced ferredoxin (Fd) towards the end of the photosynthetic electron transport chain (Fig. 14A). Since the green microalgal H2 metabolism discovered by Hans Gaffron et al. in the early 1940s,297–299 it is well-known that green microalgae can serve as the photosynthetic producers of H2, essentially derived from sunlight and H2O.299,300 Green microalgal hydrogen production can be estimated under the consideration of existing about 3 × 106 photosynthetic electron transport chains per green algal cell,301–303 each capable of transporting 100 electrons per second. Theoretically, a 1 L culture containing 10 × 106 cells per mL could produce 200 mL h−1 of hydrogen. In practice, anoxic conditions are a strict requirement for the expression and activity of the H2 production machinery in the green microalgal cell. Oxygen, produced at the H2O-oxidation site of the photosynthetic apparatus (Fig. 14A), is a potent inhibitor of the [Fe–Fe] hydrogenase and a positive suppressor of H2-related gene expression, blocking the transcription of all the genes associated with hydrogen metabolism. This may be seen as nature's provision of a powerful and effective mechanism that prevents the co-production of hydrogen and oxygen from the photosynthetic apparatus. Thus, upon turning on the illumination of a dark-adapted anoxic green microalgal culture, hydrogen production has been observed to last for as long as 90 seconds before the oxygen fully inhibits hydrogen production.304 | ||
| Fig. 14 (A) Linked H2O oxidation and H2 production in the photosynthetic apparatus of green microalgae; (B) coordinated photosynthetic and respiratory electron transport leading to anoxia (absence of oxygen) and H2 production in green microalgae; (C) cycling of a green microalgal culture between the stages of H2 production and normal photosynthesis (Normal P); (D) light-driven green microalgal H2 production, sequestration, and quantification measurements conducted in the laboratory.47 | ||
The conundrum of O2 inhibiting H2 production could not be solved in 70 years of related research.305 However, an experimental approach was designed and applied to bypass the O2 problem in 2000. Continuous photosynthetic H2 production continued for several days and achieved a regulated slow down of O2 evolution in the green algae Chlamydomonas reinhardtii.306,307 This breakthrough successfully employed the cell's own respiration to consume the photosynthetically generated O2 (ref. 298 and 308) in a process where internal starch reserves were used to sustain the cells' respiration.309 Fig. 14B depicts the coupling of cellular chloroplast photosynthesis with mitochondrial respiration to explain how anoxic conditions can be maintained in a cell permitting the expression of the HydA hydrogenase and enabling sustained hydrogen metabolism in the chloroplast. Initially, a balance of photosynthesis and respiration was achieved by the sulfur-deprivation of the algae,307 a condition that lowered the level of photosynthesis to just below that of respiration, resulting in an anoxic environment that supported hydrogen production. Maintenance of anoxia by the cell's own respiration has already become the platform of green algal H2 production in the field, and is currently employed by many labs in several countries as a vehicle to further explore the properties and premise of green microalgal H2 production.310–312 Fig. 14B also depicts the mechanism of H2 production, which depends on the availability of starch or endogenous substrates to sustain cellular respiration for the consumption of photosynthetic O2. In wild type microalgae, starch reserves can suffice to sustain hydrogen production for about 4–5 days. When starch reserves are consumed, cells need to go back to normal photosynthesis, where biomass accumulation and O2 evolution would occur. The latter is necessary and sufficient to replenish endogenous substrates and to rejuvenate the microalgae such that the stage of H2 production can be repeated. Experimental results from such cycling of the ‘stages’ are shown in Fig. 14C, where alternating O2 and H2 production could be sustained ad infinitum.308 Furthermore, the critical role of endogenous substrate in maintaining anoxia in the cells was demonstrated with the mutants of Chlamydomonas reinhardtii that over-accumulated starch. These were able to sustain H2 production for about twice as long, and achieved yields about twice as high compared to those measured with wild type strains.313 In the laboratory, the sequestration and quantification of hydrogen can be achieved by collecting H2 in upside-down graduated cylinders or burettes by the method of water displacement (Fig. 14D). The method of H2 storage by the displacement of water in glass containers satisfies the requirement of easy H2 sequestration and a subsequent easy retrieval for use. However, the method is not practical for large-scale commercial exploitation, where substantial amounts of hydrogen must be reversibly stored-and-retrieved without significant energetic expenditure.314 To date, there are no simple storage alternatives, especially when considering H2 as a fuel in the transportation sector. A main barrier is the requirement of high capacity storage and on-demand retrieval in a reversible process, where the energetic requirements of storing-and-retrieving are low. Different approaches have been investigated, including hydrogen liquefaction,315,316 compression up to 5000 psi,317,318 storage in metal hydrides,319–323 boron–nitrogen (B–N) based hydrides324–331 and adsorption (physisorption) in porous materials, notably carbon nanotubes.332,333 Current problems associated with these approaches include a combination of low capacity, high cost, high energetic requirement, and safety. Alternative methods of storing H2 in N2 (conversion to NH3), CO2 (conversion to CH4 or CH3OH) have also been proposed. Energetic and economic feasibility of the latter has not been established yet. Difficulties in hydrogen storage are an impediment in transportation, distribution, and on-board storage, all of which raise questions to the present-day practicality of renewable hydrogen in industrial and automotive applications.
In green algae, there are three pathways including two light-dependent pathways, and possibly one light-independent fermentative pathway for hydrogen evolution mediated by either [Fe]- or [Fe–Fe]-hydrogenases, both of which are unidirectional.334,335 In all the three algal pathways, the reduced Fd acts as a key station to supply electrons to the hydrogenase via the irreversible reaction: 2H+ + 2e− → H2. Given that the first two pathways are light-dependent, the electron transport chain is used to shuttle electrons (gained through the oxidation of various compounds) for the reduction of Fd. In the first pathway, water is the source of electrons and is photosynthetically oxidized via the catalytic activity of PSII. In the second pathway, however, electrons are gained through the catabolism of endogenous carbohydrate stores (i.e., the glycolysis pathway and citric acid cycle) or other organic macromolecules such as lipids. The catabolism of these compounds generates NAD(P)H molecules, which are subsequently oxidized by NADP-PQ oxidoreductase (NPQR) to liberate electrons (in addition to protons and NAD(P)+). The electrons are fed to the electron transport chain medially at the level of plastoquinone (PQ). Finally, the analysis of algal cultures placed under dark anoxic conditions revealed a putative third pathway for hydrogen evolution. Under dark anoxia, algae degrade its endogenous starch reservoirs to sustain a basal level of metabolism, generating fermentative end products such as formate, acetate, ethanol, and possibly hydrogen. Because the electron transport chain is inactive during dark periods, pyruvate provides the electrons to reduce Fd which is mediated by pyruvate ferredoxin oxidoreductase (PFR1) (Fig. 15).335,336
 | ||
| Fig. 15 Hydrogenase-catalyzed H2-photoproduction pathways in green algae.335 | ||
Because hydrogenases are the most active molecular catalysts for hydrogen production and uptake,337,338 they facilitate the development of new types of fuel cells.339–341 In [Fe–Fe]-hydrogenases (i.e., HydA1), catalysis occurs at a unique di-iron centre (the [2Fe] subsite), which contains a bridging dithiolate ligand, three CO ligands and two CN− ligands.342,343 Through a complex multi-enzymatic biosynthetic process, this [2Fe] subsite is first assembled on a maturation enzyme (i.e., HydF), and is then delivered to the apo-hydrogenase for activation.344 Synthetic chemistry has been used to prepare remarkably similar mimics of that subsite 1 but has failed to reproduce the natural enzymatic activities thus far. Berggren et al.345 proved that three synthetic mimics (containing different bridging dithiolate ligands) can be loaded onto bacterial HydF (Thermotoga maritime), and then be transferred to apo-HydA1 (one of the hydrogenases of Chlamydomonas reinhardtii algae). The full activation of HydA1 was achieved only when using the HydF hybrid protein containing the mimic with an aza dithiolate bridge, confirming the presence of this ligand in the active site of native [Fe–Fe]-hydrogenases.346,347 This is an example of controlled metalloenzyme activation using the combination of a specific protein scaffold and active-site synthetic analogues. This simple methodology provided both new mechanistic and structural insights into hydrogenase maturation and a unique tool for producing recombinant wild-type and variant [Fe–Fe]-hydrogenases with no requirement for complete maturation machinery. Because this procedure has been shown to work with proteins (HydF from Thermotoga maritima and HydA1 from Chlamydomonas reinhardtii) from two completely different organisms, it is very likely that [Fe–Fe]-hydrogenases from other microorganisms, overexpressed in their apo formin E. coli, which lacks the maturation machinery, could also be activated through simple reaction with 2-HydF. Thus, this reaction could be used for exploring a large variety of [Fe–Fe]-hydrogenases, for instance, from different species or derived from directed mutagenesis with the aim of finding the most active and stable enzymes for exploitation in biotechnological processes of H2 production,348 as well as in bioelectrodes in (photo)electrolysers or fuel cells.339–341
Cyanobacteria have three different hydrogen evolution pathways, which are different from algal pathways because of two different [Ni–Fe]-hydrogenases (bidirectional [Ni–Fe]-hydrogenases and uptake [Ni–Fe]-hydrogenases), and a [Mo–Fe]-nitrogenase found exclusively in nitrogen-fixing cyanobacteria.349 Two of them use water and organic compounds as the electron donor, and releasing electrons that are supplied to bidirectional [Ni–Fe]-hydrogenase for hydrogen evolution. However, organic compounds (i.e., glycogen) that are catabolized for hydrogen production are formed through CO2 fixation with the electrons supplied by splitting water. In this case, water is an indirect electron donor for hydrogen evolution. Because of the bidirectional nature of the cyanobacterial [Ni–Fe]-hydrogenases, hydrogen can be either produced or consumed via the reversible reaction: 2H+ + 2e− → H2. Bidirectional [Ni–Fe]-hydrogenases are considered to be associated with the cytoplasmic membrane and accept electrons from both NAD(P)H and H2 (Fig. 16).
 | ||
| Fig. 16 Hydrogenase-catalyzed H2-photoproduction pathways in cyanobacteria.335 | ||
Studies from a small number of cyanobacterial mutant strains suggests that the hydrogen evolution pathway mediated by bidirectional [Ni–Fe]-hydrogenase is possibly coupled to the photosynthetic electron transport chain. This putative pathway is different from the algal pathway as it does not solely depend on reduced Fd as an electron donor. Electrons could be shuttled directly to bidirectional [Ni–Fe]-hydrogenase at the level of NPQR (located near the photosynthetic membrane, between PSII and PQ). Electrons that are not diverted via NPQR continue along the electron transport chain through various electron acceptor intermediates (i.e., PQ, Cytb6f, PC, and PSI) for Fd reduction. While the majority of electrons gained by Fd are siphoned into other more essential assimilatory pathways (e.g., CO2 fixation), a small number of them are relayed back to NPQR through cyclic electron flow. At the onset of dark anoxia, when the electron transport chain is nonfunctional, the second hydrogen production pathway can become active. This pathway is the most widely accepted hydrogen production pathway for cyanobacteria and is analogous to the aforementioned putative fermentation pathway for hydrogen production in green algae, where NAD(P)H generated through the catabolism of endogenous glycogen stores is oxidized by NPQR to yield the electrons required for hydrogen evolution.350 The third hydrogen production pathway is found only in nitrogen-fixing cyanobacteria, in which nitrogenase fixes atmospheric nitrogen to form ammonia and hydrogen: N2 + 8H+ + 8e− + 16ATP → 2NH3 + H2 + 16ADP + 16Pi.335,351 However, this pathway is energetically expensive because 2ATP molecules are required for every electron being transferred. The electrons and ATP molecules fed to nitrogenase are obtained from either the electron transport chain associated with photosynthesis or the catabolism of carbohydrates. The electrons gained from these oxidations are first relayed to NPQR or ferredoxin/NAD(P)H oxidoreductase (FNR). NPQR donates these electrons to the electron transport chain at the level of PQ, while FNR can directly ferry the electrons to nitrogenase. Furthermore, the spent reducing power for hydrogen evolution during nitrogen fixation can be regained by hydrogen consumption via uptake [Ni–Fe]-hydrogenase. The electrons gained from hydrogen uptake are recycled back into the photosynthetic electron transport chain via the PQ pool and can be used by cytochrome c oxidase (cyt. c) for the reduction of O2 to water (i.e., Mehler reaction) or transferred back to nitrogenase via PSI and a heterocyst-specific Fd.350,352
Biohydrogen production by microalgae is considered to be the most favorable pathway.353 Microalgae splits water into proton (H+) and oxygen (O2) in the presence of light. The process can convert H+ into hydrogen via hydrognease, known as direct-photolysis.354 However, hydrogen production in this process is low because of two main reasons that H2 and O2 are produced concomitantly, mixing and reacting into H2O immediately, and hydrognease itself is sensitive to oxygen.355,356 This inhibitory effect can be fixed by adopting indirect bio-photolysis, consisting of two stages of stage-I and stage-II called the aerobic and anaerobic stages, respectively. In stage-I, the cells undergo photosynthesis to accumulate organic compounds (mostly glucose) and oxygen is evolved. In stage-II, the cells degrade stored organic compounds under anaerobic conditions.357 In a two-stage process, oxygen (in stage-I) and hydrogen (in stage II) are evolved separately. Stage-II can be under a light condition called photo-fermentation or without light named dark fermentation.358 Fig. 17 illustrates the concept of two-staged hydrogen production by microalgae. Several factors affect hydrogen yield in stage-I and stage-II. Healthy cells in stage-I produce hydrogen efficiently. Microalgae growth in stage-I is controlled by different parameters such as light, nutrients, carbon source, temperature, pH, and bioreactor design. These parameters are also equally important in stage-II. Immobilization and sulfur deprivation are the key intermediate steps of stage-I and stage-II. For immobilization, the cells are suspended in a solidifying material and cut into small pieces. Immobilized cells are easy to handle, have high stability and produce more hydrogen than free cells. Sulfur deprived (S-deprived) cells yield more hydrogen than sulfur-provided cells. In the presence of sulfur, the cell synthesizes protein which suppresses the hydrogen production.359
 | ||
| Fig. 17 Concept of two-staged biohydrogen production by microalgae. | ||
To determine energy consumption and CO2 emissions, Ferreira et al.367 presented a life cycle inventory of biohydrogen production by Clostridium butyricum through the fermentation of whole Scenedesmus obliquus, which was accomplished by the fermentation of microalgal biomass cultivated in an outdoor raceway pond and the preparation of the inoculum and culture media. The scale-up scenarios are discussed aiming for a potential application to a fuel cell hybrid taxi fleet. The H2 yield obtained was 7.3 g H2 kg−1 of S. obliquus dried biomass. A total energy consumption of 88 (71–100) MJ per MJH2 and 5776 (5119–6268) gCO2 per MJH2 emissions was obtained, which is considerably high and unsustainable if pilot/industrial scale is envisaged. The stage of microalgae culture required the highest energy consumption (55 MJ per MJH2) and emitted the maximum CO2 (3605 gCO2 per MJH2), respectively, and contributing 62.4% of energy consumption in the overall process. When CO2 absorption is considered, the microalgae culture becomes responsible for 41.1% of the overall CO2 emissions with 1516 gCO2 per MJH2. Other studies and production technologies were taken into account to discuss an eventual process scale-up. Increased production rates of microalgal biomass and biohydrogen are necessary to become competitive with conventional production pathways.
Recent comprehensive evaluation studies on the feasibility of algal biofuel production, performed by the Solar Biofuels Consortium, concluded that diversification into various co-products is important for the development of a standalone microalgal biofuels industry.117 Consequently, new biorefinery concepts are required to combine hydrogen with the production of other biofuels, such as biogas (methane), oils (i.e., biodiesel), and the separation of valuable co-products. Such biorefinery concepts can be designed with the aim of achieving CO2 neutral systems in which CO2 and nutrients are recycled (Fig. 18). Because H2 is a volatile product that can be readily collected from the culture, hydrogen can be considered to be an excellent component of such new bio-refinery concepts.311
 | ||
| Fig. 18 Concept of a biorefinery system for bio-energy and bio-products in algae. Bio-products include bio-energy products such as hydrogen, oils for bio-diesel, sugars for bio-ethanol and biomass for bio-methane, intermediate value products such as proteins for animal feedstocks, and high value products (HVPs), for example, for pharmaceutical purposes. CO2 and nutrients released during the fermentation of residual biomass during the production of bio-methane will be recycled. Biomass can also be pyrolyzed to produce ‘sequestered carbon’ in the form of biochar, which has value as a soil enhancer.311 | ||
High-yield biohydrogen production in combination with photosynthesis will require an oxygen-tolerant hydrogenase (i.e., [Fe-only]-hydrogenase). This could be achieved by the intelligent combination of random mutagenesis, site-directed mutagenesis and directed evolution, which has already been applied successfully to improve other enzymes.375 For instance, the existing oxygen-tolerant hydrogenases of Ralstonia eutropha with its identified maturation apparatus376 is a valuable starting point. Most recent strategies in this field are summarized.377 If successful, the future scenario for a designed organism with engineered biophotolytical hydrogen production might be similar to the model (Fig. 19).378 Future energy balances for such systems should consider the following parameters: (1) progress in energy transformation efficiency that can be hopefully obtained using designed organisms with improved hydrogenases; (2) development of high energy content algal biomass, low-cost fermenters and media; (3) decreasing the doubling time of algal culture; (4) the option to use sunlight instead of artificial light (indoor systems would also be possible using fiber optics). The environmental benefits derived from zero-CO2 emission and the increasing costs of gasoline and natural gas should eventually make the natural system, which still has potential for improvement, more competitive.
 | ||
| Fig. 19 Circuit of water and hydrogen in a system consisting of hydrogen-producing microalgae and a fuel cell that transforms hydrogen into electrical energy.378 | ||
For biohydrogen applications, it is mentioned above that biohydrogen produced from microalgae could be widely used for hydrogen–oxygen fuel cells driving fuel cell powered vehicles (FCVs). This is not only environmentally friendly and highly energy-efficient, but can also be produced using a variety of readily available raw materials. Because of these characteristics, FCVs are ideal for achieving sustainable mobility. Therefore, many automobile manufactures have tried their best to make this vehicle technology widely available as soon as possible, as shown in Fig. 20. Some significant components, such as hydrogen, oxygen, catalysts, membrane, circuit, have attracted more attention and need to be developed further for designing superior performing FCVs.
 | ||
| Fig. 20 Schematic diagram of biohydrogen fuel cell vehicle. | ||
 | ||
| Fig. 21 A new concept for enhanced hydrogen production and electricity generation by microalgae. | ||
Biohydrogen is usually produced via dark fermentation, which emits CO2 and produces soluble metabolites (e.g., volatile fatty acids) with a high chemical oxygen demand (COD) and by-products that require further treatment. Liu et al.384 successfully demonstrated the feasibility of a novel integration of dark fermentation and mixotrophic microalgae culture, allowing efficient biohydrogen production with minimal CO2 emissions and no COD discharge, by circulating the byproducts of dark fermentation and biomass from microalgae culture. The results showed that when this integrated system was used the production rate of H2 was 205 mL L−1 h−1 with only 5 mL L−1 h−1 of CO2 emission. The microalgae-based COD removal of dark fermentation effluent was the most efficient when C. vulgaris was grown as a food for microorganism (F/M) ratio of 4.5 with a light intensity of 150 mmol m−2 s−1. The addition of CO2 for mixotrophic microalgae growth would improve the overall microalgal biomass production performance but lead to a decrease in butyrate consumption efficiency due to the competition of organic and inorganic carbon sources. Meanwhile, Kumar et al.385 confirmed the pretreated algal biomass of 10 g L−1 with 2% (v/v) HCl-heat was found most suitable for hydrogen production yielding 9 ± 2 mol H2 (kg COD reduced)−1 and was found to fit with a modified Gompertz equation. Furthermore, hydrogen energy recovery in dark fermentation was significantly enhanced compared to the earlier reports of hydrogen production by the biophotolysis of algae.
To enhance the efficiency of H2 production from pretreated feedstock, the pretreatment method and hydrolysis conditions should be optimized.386,387 Yun et al.386 optimized the individual pretreatment (acid and ultrasonic) and a combination of these pretreatments to enhance the efficiency of dark fermentative hydrogen production (DFHP) from microalgal biomass. They showed that the maximum H2 production performance of 42.1 mL H2 per g dry cell weight (dcw) was predicted at 0.79% (v/w) HCl at a specific energy input of 49![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 600 kJ kg−1 dcw in the combined pretreatment, while it was limited in both the individual pretreatments. Moreover, the combined pretreatment conditions for DFHP from microalgal biomass were successfully optimized by increasing the solubilization of the feedstock and reducing the formation of the toxic 5-hydroxymethylfurfural (HMF).
600 kJ kg−1 dcw in the combined pretreatment, while it was limited in both the individual pretreatments. Moreover, the combined pretreatment conditions for DFHP from microalgal biomass were successfully optimized by increasing the solubilization of the feedstock and reducing the formation of the toxic 5-hydroxymethylfurfural (HMF).
Recently, Xia et al.388 investigated for the first time the thermodynamic comparison in dark fermentation between amino acids and reducing sugars released from Nannochloropsis oceanica. A three-stage method comprising dark fermentation, photo fermentation and methanogenesis388,389 was proposed to improve hydrogen and energy yields from N. oceanica. The total utilization efficiencies of amino acids and reducing sugars are both about 95% in dark fermentation, but the consumption time of most amino acids is about twice as long as that of most reducing sugars in dark fermentation. Overall, the maximum hydrogen yield of 183.9 mL g−1-total volatile solids (TVS) and a methane yield of 161.3 mL g−1-TVS are achieved from N. oceanica biomass through the three-stage method. The total energy yield of hydrogen and methane from microalgae biomass through the three-stage method is 1.7 and 1.3 times higher than those through the two-stage (dark fermentation and methanogenesis) and single-stage (methanogenesis) methods, respectively. During the stages of hydrogen production there are energy demands, mainly of electricity and associated CO2 emissions. Fig. 22 shows the microalgal biomass production and the entire fermentation process and corresponding inputs. The main stages considered were the microalgal biomass production, the fermentation medium preparation, which included BM1 preparation and hydrolysis of microalgal biomass, degasification and fermentation.
 | ||
| Fig. 22 Scheme of the experimental stages of biomass production and the entire fermentation process and corresponding imputs/outputs: (A) Scenedesmus obliquus biomass production, (B) BM1 preparation, (C) biomass hydrolysis and (D) fermentation. | ||
Ferreira et al.390 presented the life cycle inventory of hydrogen production by Clostridium butyricum fermentation of Scenedesmus obliquus hydrolysate to evaluate the potential of H2 production from microalgae and the respective energy consumption and CO2 emissions in the bioconversion process considering the microalga production, acid hydrolysis of S. obliquus, preparation of the inoculum and culture media, and fermentation. In this work, the H2 yield was 2.9 ± 0.3 mol H2 per mol sugars in S. obliquus hydrolysate. Results showed that this process of biological production of hydrogen can achieve 7270 MJ per MJH2 of energy consumption and 670 kg CO2 per MJH2. Microalgal culture was responsible for 98% of these total final values with the use of artificial lighting. All the stages and processes with the highest values of energy consumption and CO2 emissions were identified for future energetic and environmental optimization.
To decrease the energy consumption and associated CO2 emissions, the experimental procedure must be optimized aiming at processing a larger amount of biomass to achieve production at an industrial scale. With the present results, it is possible to identify the most critical steps of the entire fermentation process that can be optimized in terms of energy saving and CO2 emissions reduction. In this study, the microalgae were produced indoor with artificial light. A possible solution to reduce energy consumption and CO2 emissions in the experimental hydrogen production is to replace artificial light used for the microalgal growth by sunlight with much less electricity consumption. The dryness process could also be performed by wind or solar energy especially because this study is conducted in a country with favorable climatic conditions. Therefore, it would be possible to reduce the values obtained of 308–441 MJ per MJH2 and 28.5–36.3 kg CO2 per MJH2 by reducing 98.5% of the total electricity used. In addition, other possible scenarios could include the substitution of the “degasification 1” (Fig. 22) by a unique step of degasification of BM1 medium, rendering a 0.13% of electricity saving. Moreover, using whole acid-treated S. obliquus as carbon substrate would avoid the steps of centrifugation and filtration for the solid–liquid separation, resulting in a further decrease of 0.1% in electricity consumption. With all these possibilities it would be possible to reduce final energy consumption and CO2 emissions by 98.7%. Fig. 23 shows the scheme of the optimized microalgal biomass production and the entire fermentation process. Advanced techniques such as electrocoagulation for microalgae culture harvesting, dewatering of microalgal biomass in solar ovens and wind tunnels, the use of hybrid fermentation systems and recombinant microorganisms should also be considered for further process improvement.
 | ||
| Fig. 23 Scheme of the optimized biomass production and the entire fermentation process and corresponding inputs/outputs: (A) Scenedesmus obliquus biomass production, (B) BM1 preparation, (C) biomass hydrolysis and (D) fermentation. | ||
Metabolic engineering is also a tool that can bring a major breakthrough in biohydrogen production. By exploring the pathway of hydrogen production using molecular biology, this technique can eliminate bottlenecks and increase carbon flow to hydrogen-producing pathways. It can also be favorable to increase substrate utilization by engineering more efficient and oxygen resistant hydrogen evolving enzymes.391 The C. reinhardtii genome sequence showed several unexpected pathways such as inorganic carbon fixation, fermentation, and vitamin biosynthesis.391–393 Each of them can be exploited to improve the biohydrogen yield. Exploring nutrient limitation and substrate utilization can benefit from discovering particular chromosomal genes in microalgae for hydrogen production enhancement.391 Random and direct mutagenesis has succeeded in improving tolerance 10-fold. One approach to address this problem is gene shuffling, which has been used to generate a diverse recombinant hydrogenase library to screen for enhanced O2 tolerance and stability.391,394 Algal hydrogenase (HydA) is in charge of catalyzing the reaction: 2H+ + 2e− ↔ H2 but it is usually inhibited by O2, which is a byproduct of photosynthesis. Therefore, Lin et al.395 studied to knockdown PsbO, a subunit concerned with O2 evolution, so that it would lead to HydA induction. The green alga (Chlorella sp. DT) was then transformed with short interference RNA antisense-psbO (siRNA-psbO) fragments. The algal mutants were selected by checking for the presence of siRNA-psbO fragments in their genomes and the low amount of PsbO proteins. The HydA transcription and expression were observed in the PsbO-knockdown mutants. Under semi-aerobic conditions, PsbO-knockdown mutants could photobiologically produce H2, which increased as much as 10-fold in comparison to the wild type.
A new strategy has been introduced to search natural diversity through the use of degenerate polymerase chain reaction (PCR) primers.391 Developments are required for the optimum design of PBRs. Another critical issue is to find a cheaper carbon source that could produce hydrogen efficiently. To address the economy of this process, the shortening of the total time of hydrogen production should be the top priority. The use of optical fiber is an approach to strikingly decrease the lag time for hydrogen production. Biohydrogen is still more expensive than other fuels. Thus, if technology improvements succeed in reducing the costs, it can attain considerable attention as a sustainable biofuel. The optimization of key experimental factors, genetic modification, and metabolic engineering of microalgae are the ultimate approaches to make hydrogen production cost-effective and sustainable. Catabolism of glycogen stored by cyanobacteria occurs during anaerobic auto-fermentation and produces a range of C1–C3 fermentation products and hydrogen via hydrogenase. Kenchappa et al.396 investigated both augmenting and rerouting this carbon catabolism by engineering the glycolysis pathway at the NAD+-dependent glyceraldehyde-3-phosphate dehydrogenase (GAPDH-1), its major regulation site at the nexus of two pathways [e.g., oxidative pentose phosphate (OPP) pathway and glycolysis/gluconeogenesis] (Fig. 24). Null (gap1::aphII) and overexpression (gap1+) strains of Synechococcus sp. strain were constructed to produce more NADPH (via rerouting carbon through OPP) and more NADH (via opening the glycolytic bottleneck), respectively. For gap1::aphII quantitative analyses, after four-days of dark auto-fermentation showed an undiminished glycogen catabolism rate, significant increases of intracellular metabolites in both OPP and upper-glycolysis, a decrease in lower-glycolysis intermediates, 5.7-fold increase in NADPH pool, 2.3-fold increase in hydrogen and 1.25-fold increase in CO2 vs. wild type (WT). These changes demonstrate the expected outcome of the redirection of carbon catabolism through the OPP pathway with significant stimulation of OPP product yields. The gap1+ strain exhibits a large 17% increase in the accumulation of glycogen during the prior photoautotrophic growth stage (gluconeogenesis), in parallel with a 2-fold increase in the total [NAD+ + NADH] pool, foreshadowing an increased catabolic capacity. Indeed, the rate of glycogen catabolism during subsequent dark auto-fermentation increased significantly (58%) vs. WT, resulting in increases in both NADH (4.0-fold) and NADPH (2.9-fold) pools, and terminal fermentation products, hydrogen (3.0-fold) D-lactate (2.3-fold) and acetate (1.4-fold). The overall energy conversion yield over four days from catabolized glycogen to hydrogen increased from 0.6 mole of hydrogen mole−1 of glucose (WT) to 1.4 (gap1::aphII) and 1.1 (gap1+) under headspace accumulation conditions (without hydrogen milking). It has demonstrated that metabolic engineering has a significant potential for redirecting carbon pathways on carbohydrate catabolism and hydrogen production in cyanobacteria.
 | ||
| Fig. 24 (A) Schematic representation of theoretical yields of NAD(P)H by glycolysis and oxidative pentose phosphate (OPP) pathways; (B) possible yields of hydrogen per mole of glucose via glycolysis and OPP pathways. Metabolites: G6P = glucose-6-phosphate; F6P = fructose-6-phosphate; FBP = fructose-1,6-bisphosphate; GAP = glyceraldehyde-3-phosphate; BPG = 1,3-bisphosphoglycerate; PEP = phosphoenolpyruvate; 6 PG = 6-phosphogluconate; Ru5P = ribulose-5-phosphate. Enzymes: GAPDH-1 = NAD+-dependent glyceraldehyde-3-phosphate dehydrogenase; TH = transhydrogenase. | ||
3.4. Biodiesel
First generation biofuels derived from agricultural edible crop oils accounting for more than 95% of biodiesel sources have a considerable impact on food security and have the potential to increase the cost of food crops (i.e., soybean, corn) resulting in biodiesel production being more expensive.397 Second generation biofuels (i.e., jatropha oil, waste cooking oil and animal fats) do not affect food security and have significant advantages over first generation oil crops but they are unsustainable. Moreover, the production of crop-derived biofuels brings on new challenges. For example, poor cold flow properties and saturated fatty acids contained in animal fats may cause production difficulties and constitute a bio-safety hazard because of their solid nature at room temperature.398 In terms of social and economic acceptability and greater energy security, microalgal oil is regarded as a third generation biofuels source. Algae can produce twenty times that of oilseed crops on a per hectare basis, and thus it is a more viable alternative.97,116,399,400 Microalgae have faster growth rates than plants and are capable of growth in highly saline waters, which are unsuitable for agriculture. They utilize a large fraction of solar energy making them effective solar to chemical energy converters.401,402 Microalgae have greater photosynthetic efficiency than terrestrial plants and require very little and simple nutrients for growth.400 Normally, a dry cellular weight basis lipid content of microalgae generally varies between 20% and 40%, while lipid contents as high as 85% have been reported for certain microalgal strains.403–405 The triglycerides productivity of microalgae could be 25–220 times higher than terrestrial plants,401 which can be readily converted to biodiesel by the transesterification process.404,406 As compared to biomass from trees and crops, microalgal oil is more economical because transportation costs are relatively low.398 Algae-to-energy systems can be either net energy positive or negative depending on the specific combination of cultivation and conversion processes used. Conversion pathways involving direct combustion for bioelectricity production generally outperformed systems involving anaerobic digestion and biodiesel production.407 Therefore, microalgae offers a significant higher yield advantage as potential feedstock for biodiesel production.58,408–410 Tang et al.411 examined the influence of light, CO2 concentration, and photoperiod on the growth of the D. tertiolecata. The results indicated that white and red LEDs, and fluorescent lights all are good light sources and a higher light intensity can significantly improve the cell growth. CO2 levels of 2 to 6% in air provided the highest growth rates. Continuous lighting also significantly increased the biomass productivity of D. tertiolecta. Differences in light source and intensity had no significant effect on the content and composition of fatty acid methyl esters (FAME, the components of biodiesel) from D. tertiolecta oil. Finally, a high content of C18:3 of D. tertiolecta biodiesel may lead to poor oxidative stability. However, the high growth rate and ability of these microalgae to grow in a brackish environment makes them a good candidate for biofuel production via other pathways.The first step for any biodiesel technology involves oil extraction from the biomass source. This step is relatively well established for edible feedstocks and more troublesome for waste oils (the presence of water and free fatty acid impurities) and algae (lack of efficient methodologies for oil extraction). Vegetable oils, which are rich in triglycerides (TGs), are subsequently treated with methanol under mild temperatures (50–80 °C) in the presence of a basic homogeneous catalyst (Fig. 25A). The process is transesterification, which allows the conversion of TGs in a mixture of FAME and glycerol (1,2,3-propanetriol). A large part of this co-produced glycerol is separated from FAME by simple decantation, although further washing/drying steps are required to remove the traces of glycerol to comply with the strict regulations for fuel grade biodiesel. This extra purification process increases production costs and generates great amounts of salts, soaps and waste water. Furthermore, the management of large amounts of residual crude glycerol produced (100 kg per ton of biofuel) represents an important challenge for the biodiesel industry. Fig. 25B shows a comparative scheme of transesterification and hydrotreating processes. Both technologies utilize TGs as feedstocks but they differ in the reactants utilized (methanol vs. hydrogen), the by-products generated (glycerol vs. propane), the final fuel product obtained (biodiesel vs. green hydrocarbons) as well as in the reaction conditions and catalysts used. Methanol and hydrogen are typically derived from fossil fuels and therefore efforts should be made to obtain these reactants from biomass sources in order to reduce the overall CO2 footprint of biofuel. While solutions in the biodiesel industry involve replacement of methanol with biomass-derived ethanol as an esterification agent, hydrotreating technologies can drastically reduce external hydrogen consumption by employing sub-products and/or residues generated during the process as sources of this gas. For example, up to 75% of H2 hydrotreating needs can be covered by steam reforming and a subsequent water gas shift (WGS) of propane co-produced during the process,413 while the lignocellulosic soybean hull wastes discarded after oil extraction can provide hydrogen for hydroprocessing by means of microbial fermentation.414 The higher cost of hydrogen compared to methanol should be a strong incentive to implement the mentioned solutions in commercial hydrotreating plants. The separation and subsequent management of the by-products generated during the process is also an important aspect determining the profitability of both technologies. In this sense, transesterification seems to be more sensitive to this parameter given the large amounts of glycerol generated and the difficulty to completely remove it from the biodiesel fuel (Fig. 25A). However, once separated, this crude glycerol can serve as a cheap feedstock for the production of a large variety of high value-added chemicals and fuels,415 thereby representing an opportunity to reduce the overall biodiesel production costs.416 On the other hand, hydrotreating generates a by-product gas stream enriched in propane, which is easily separable from the liquid hydrocarbon fuel but presents a lower chemical value compared to glycerol. Consequently, instead of decreasing the costs, this gas stream could be important to reduce the overall input of fossil fuels in the process by offering an internal source of hydrogen or heat/electricity.417
 | ||
| Fig. 25 (A) Summarized scheme of the transesterification process used for the production of biodiesel from algal biomass; (B) comparative scheme of transesterification and hydrotreating processes for the conversion of triglycerides into biodiesel and green hydrocarbons, respectively. | ||
Transesterification of lipid feedstocks requires milder temperature and pressure conditions compared to hydrotreating, so the operational costs are greater for the latter route. Nevertheless, hydrotreating conditions are similar to those used in hydrodenitrogeneration (HDN) and hydrodesulfurization (HDS) of petroleum, which provides the possibility to co-process lipids and fossil feeds in existing refinery facilities.418,419 This synergy between hydrotreating and conventional oil refineries would considerably reduce capital costs,420 and represent one of the key advantages of hydrotreating vs. conventional transesterification. However, some key points on hydrotreating still require further research studies, such as the corrosion of the hydroprocessing reactor by free fatty acids, the detrimental cold flow properties of the diesel product as a consequence of the increased content of n-alkanes,421,422 and the effect of the presence of oxygenates over intrinsic HDN and HDS activities of commercial hydroprocessing catalysts. The simplified version of the chemistry involved in transesterification and hydrotreating allows the production of biodiesel and green hydrocarbons with high yields. In this sense, both technologies benefit from the utilization of feeds with relatively low oxygen content (and thus low reactivity) like TGs to achieve the required transformations in a selective fashion, and this represents an important advantage compared with other biomass conversion routes managing more reactive feedstocks (i.e., sugars, lignocellulosic biomass). However, the latter feedstocks are more abundant and cheaper than vegetable oils. So far, the limited availability of lipids to satisfy the growing demand for both biodiesel and green hydrocarbon fuels is the most important issue facing both transesterification and hydrotreating technologies. Therefore, it is imperative to search for additional and preferable non-edible lipid sources that can ensure a sustainable supply without affecting food markets or requiring large land extensions. Hydrotreating presents higher flexibility to cope with different kinds of feeds compared to transesterification, which is more sensitive to the presence of impurities or free fatty acids. In this sense, hydrotreating is better positioned for the implementation of new, more abundant and non food-competitive feedstocks (e.g., algae) in the near future.
Transesterification of algal biomass or lipid to yield biodiesel can be performed by the following common methods such as conventional heating,71 supercritical methanol conditions,274 enzyme-catalyzed methods,423 and microwave irradiation.424 Among these methods, conventional heating requires longer reaction times with higher-energy inputs and losses to the environment. Supercritical methanol processing operates in expensive reactors at high temperatures and pressures resulting in higher-energy inputs and higher production costs. The enzymatic transesterification reaction proceeds with a slower reaction rate and there is a strong possibility of enzyme inactivation by methanol during the process. Of the four methods, microwave-assisted transesterification is the most energy-efficient, quick and reliable process to produce biodiesel from algal biomass. The two basic mechanisms of oil extraction from algal biomass observed during a microwave irradiation process are reported as the diffusion of lipids across the cell wall into the solvent due to the selectivity and solubility and disruption of the cell wall with a release of contents into the solvent.425 The direct conversion (in situ transesterification) of algal biomass under microwave irradiation conditions has proven to be an effective method for biodiesel production as this method achieves a high degree of oil-lipid removal from the dry algal biomass and efficiently converts oils and lipids to biodiesel.424,426 It also reduces the reaction time and the solvent volume as compared with the separate lipid extraction and transesterification processes. However, the application of suitable power dissipation control in microwave-assisted transesterification reactions may result in a greater benefit in terms of energy efficiency and reaction product yield. Furthermore, Patil et al.427 studied the effects of power dissipation on microwave-enhanced in situ transesterification of dry algal biomass (Nannochloropsis salina) to biodiesel fuel as well. The microwave for the transesterification reaction has twofold effects of enhancing the reaction by a thermal effect and evaporating methanol due to the strong microwave interaction of the material.428,429 The microwave interaction with the reaction compounds (triglycerides and methanol) results in a large reduction of the activation energy due to an increased dipolar polarization phenomenon. This is achieved due to molecular-level interactions of the microwaves in the reaction mixture resulting in dipolar rotation and ionic conduction.430 The reduction in activation energy is essentially dependent on the medium and reaction mechanism.431 Methanol is a strong microwave absorption material and, in general, the presence of an OH group attached to a large molecule behaves as though it were anchored to an immobile raft, and the more localized rotations dominate the microwave spectrum and result in localized superheating, which assists the reaction to complete faster.432 The microwave enhanced transesterification reaction of algal biomass to yield methyl ester is illustrated in Fig. 26. The base-catalyzed microwave transesterification mechanism is described elsewhere.426
 | ||
| Fig. 26 Microwave-enhanced in situ transesterification of algal biomass.427 | ||
 | ||
| Fig. 27 Schematic diagram of production (A) and principle (B) of emulsified biodiesel production. | ||
 | ||
| Fig. 28 Algal biofuel pipeline, showing the major stages in the process, together with the inputs and outputs that must be taken into consideration by LC.438 | ||
Although algae-based biodiesel production is still at the research and development stage, it is reasonable to expect that algal biofuel production, when commercially implemented, will resemble existing industrial processes. Hence, some process steps within the system boundary (e.g., dewatering and drying of algae) are modeled using data for other similar processes being currently practiced. Fig. 29 shows the system boundary of the biodiesel production process. The life cycle impacts were assessed for an integrated microalgal biodiesel production system that facilitates energy- and nutrient-recovery through anaerobic digestion, and utilizes glycerol generated within the facility for additional heterotrophic biodiesel production.449 Efforts to increase productivity but reduce input and cost through process engineering and the use of transgenic methods, and classical breeding aimed at developing domesticated algal crop strains, will benefit both strategies.450 The successful, large-scale generation of biodiesel from microalgal feedstocks will require viewing the algal production facilities as biologically diverse bioreactors that will obey the known rules of ecology. In the three subsections below we illustrate how the application of core concepts and principles from ecology and ecological physiology can provide important new insights into the design and operation of these systems.451,452
 | ||
| Fig. 29 System boundary of the biodiesel production process. | ||
Utilization as a potential feedstock for biodiesel production, microalgae need to overcome a few limitations, including low growth rates of photoautotrophic algae and low biomass concentrations. The algal species can grow only in a specified temperature range (15–30 °C) and fluctuation of temperature beyond the optimum range results in inhibition of growth of the microalga or its death. To achieve the desired temperature range in open ponds may be difficult as temperature of surface can go as high as 40 °C. Therefore, closed bioreactors are fabricated for the microalgae culture to minimize temperature fluctuations. However, closed bioreactors too, if operated in hot areas, may undergo an increase in temperature, which has to be controlled by using water for evaporative cooling, heat exchangers, reflection of infra-red radiation, or light dilutions. These processes make the microalgal biodiesel cost intensive. Synthesis of biodiesel from microalgae at present is low and needs further improvement in the cultivation process. Krohn et al.453 found that though the total lipids comprised 19% of algal dry weight, the synthesized biodiesel from the lipids were only 1% of dry weight. Algal lipids possess high free fatty acid content which is not saponifiable and so transesterification cannot be done with the conventional homogeneous base catalyst.453 The option available is to reduce the acid value by esterification or employing a solid acid catalyst. The deprivation of nitrogen on the accumulation of lipids in microalgae varies among the various species. A limitation of synthesis of biodiesel from microalgae is a high alcohol to oil molar ratio (up to the extent of 315:1) required during the synthesis process that enhances the production cost of biodiesel.454 Another major limitation of the oil obtained from microalgae, yeast, and fungi is lipid content (broadly classified as neutral lipids, total lipid). Only a part of the neutral lipid that comprises of triglycerides and free fatty acids can be converted to fatty acid methyl esters (i.e., biodiesel) and many of microalgae tried as feedstock for oil comprises of constituents that cannot be converted to biodiesel.455
3.5. Other biofuels production from algae
Bioethanol from algae holds significant potential due to its low percentage of lignin and hemicellulose compared to other lignocellulosic plants.456 With a low lignin content, macroalgae contain amounts of sugars (at least 50%) that could be used in fermentation for bioethanol production.457,458 However, in certain marine algae, such as red algae, the carbohydrate content is influenced by the presence of agar, a polymer of galactose and galactopyranose. Current research seeks to develop approaches of saccharification to unlock galactose from the agar and further release glucose from cellulose leading to higher ethanol yields during fermentation.458,459 Until now, only a handful of research work has been reported on bioethanol production from microalgae for several reasons. Firstly, more attention has been diverted to biodiesel production from microalgae since certain strains are capable of accumulating large quantities of lipid naturally inside their cells; secondly, through nitrogen-deficient cultivation methods (to save energy and cost), lipid content inside the microalgae cells is boosted up significantly by blocking carbohydrate synthesis pathways, while carbohydrate is the main substrate to produce bioethanol. Moreover, biodiesel has a higher calorific value than bioethanol, which are compared to be 37.3 MJ kg−1 and 26.7 MJ kg−1, respectively. Nonetheless, microalgae are found to be a superior feedstock to produce bioethanol in comparison with other first and second generation bioethanol feedstocks. First generation bioethanol is derived from food feedstock such as sugar cane and sugar beet, in which over exploitation of this feedstock creates the “food versus fuel” issues and raised several environmental problems including deforestation and ineffective land utilization. Second generation bioethanol is produced from lignocellulosic biomass such as wood, rice straw and corn stover, which should be subjected to pretreat initially to break down the complex structure of lignin and to decrease the fraction of crystalline cellulose by converting it to amorphous cellulose.460 However, most of the pre-treatment methods, i.e., steam explosion and alkali or acid pre-treatment, are energy intensive and negatively impact the environment.In contrast, microalgae cells are buoyant and do not require lignin and hemicelluloses for structural support.461 Therefore, it is expected that the overall bioethanol production process can be simplified due to the non-requirement for a chemical and enzymatic pre-treatment step. Nevertheless, it should be noted that high concentration of carbohydrates are actually entrapped within the microalgae cell wall, in which an economical physical pre-treatment process such as extrusion and mechanical shear is still required to break down the cell wall so that the carbohydrates can be released and converted to fermentable sugars for bioethanol production.461
Green algae including Spirogyra sp. and Chlorococum sp. have been shown to accumulate high levels of polysaccharides both in their complex cell walls and as starch. This starch accumulation can be used in the production of bioethanol.456,462 Harun et al.456 proved that the green algae Chlorococum sp. produces 60% higher ethanol concentrations for samples that are pre-extracted for lipids versus those that remain as dried intact cells. This indicates that microalgae can be used for both lipid-based biofuels and ethanol biofuels production from the same biomass as a means to increase their overall economic value. On the other hand, simultaneous biodiesel and bioethanol production from microalgae is also possible, in which microalgae lipid is extracted prior to a fermentation process.463,464 This concept has proven to be viable in a recent study in which lipids from Chlorococum sp. were extracted with supercritical CO2 at 60 °C and subsequently subjected to fermentation by the yeast Saccharomyces bayanus.456 From the report, microalgae with pre-extracted lipid gave 60% higher ethanol concentration for all samples than the dried microalgae without lipid extraction. This is because supercritical CO2 can act as a superior pre-treatment method to breakdown microalgae cell wall, causing the simultaneous release of lipid and carbohydrates embedded within the cell wall. A maximum bioethanol yield of 3.83 g L−1 was achieved from 10 g L−1 of lipid-extracted microalgae residue. In other words, lipid extraction from microalgae for biodiesel production and a pre-treatment step to release carbohydrates for bioethanol production occurs in just one single step, which greatly enhances the viability of microalgae biofuels production in commercial scale. Apart from supercritical CO2, other lipid extraction methods such as ultrasonication, chemical solvent, microwave and use of a bead-beater have not been studied to get a comprehensive comparison between these methods.463
Fig. 30 shows the block diagram of the superstructure for the integrated production of bioethanol and biodiesel from algae. The actual flowsheet, including all the different units, can be reconstructed using the detailed figures presented along the text. Firstly, algae are grown in ponds. After that, the oil is extracted by using an organic solvent. Finally, the starch is separated, which is saccharified and liquefied for the production of ethanol. In parallel, the oil is transesterified using the dehydrated ethanol. Two most promising alternatives were considered for the transesterification of oil using bioethanol,465 the use of a homogeneous alkali catalyst or the enzymatic catalyzed reaction. The ethanol is recovered, recycled, and mixed with part of the ethanol produced from the starch and the glycerol is separated from the product biodiesel, which in this case is fatty acid ethyl ester (FAEE). Then, Martín et al.464 also presented two alternative technologies for the biodiesel synthesis from algae oil, enzymatic or homogeneous alkali catalyzed that are coupled with bioethanol production from algae starch. To determine the optimal operating conditions, they not only couple the technologies, but also simultaneously optimize the production of both biofuels and heat integration while optimizing the water consumption. Multi-effect distillation is included to reduce the energy and cooling water consumption for ethanol dehydration. In both cases, the optimal algae composition results in 60% oil, 30% starch, and 10% protein. The best alternative for the production of biofuels corresponds to a production price of 0.35 $ gal−1, using enzymes, with energy and water consumption values of 4.00 MJ gal−1 and 0.59 gal gal−1, respectively. Even though the integrated process requires higher energy and water consumption, the simultaneous production of ethanol and biodiesel is more advantageous than the production of biodiesel using ethanol alone as it reduces the biofuel production cost around 20%, mostly because of the raw material cost reduction.
 | ||
| Fig. 30 An integrated concept of production of bioethanol and biodiesel from algae. | ||
Microalgal biomass is still not a viable choice for commercial biofuels production due to the extensive energy input required compared to current terrestrial energy crops. Nevertheless, several energy hotspots have been indicated in the overall microalgae process chain, including inorganic nitrogen source production, operation of the photobioreactor and harvesting/dewatering of microalgal biomass. It is recommended that culturing microalgae for biofuels production should be coupled with wastewater treatment and waste CO2 to minimize the heavy dependency on the inorganic nutrients and carbon sources. For the downstream processes, extraction of lipid from microalgae presents a complicated task, as there is no single method that can give optimum lipid extraction for all types of microalgae strains. Thus, breakthrough technologies such as supercritical extraction/transesterification, in situ transesterification, hydrothermal treatment and transesterfication assisted with ultrasonication or microwave radiation have a significant potential to significantly enhance the production of microalgae biodiesel. In addition, the simultaneous production of bioethanol and biodiesel is more advantageous than the biodiesel production using ethanol alone, thereby reducing the biofuel production cost by around 20%. For long-term sustainability and environmental benefits, all the processing stages of microalgae biofuels should be simplified without the involvement of an extensive energy input. In addition, the processes should be easily adopted in the existing biofuels industry and can be implemented especially in third world countries, for culturing microalgae for biofuels production is not only meant for profit making and benefiting the environment, but also to help people from the bottom billions in terms of food and energy security. The integrated process of microalgae biofuels production via combining wastewater treatment with CO2 bio-mitigation has been attracting increasing attention by researchers, which will be discussed in the following part.
4. Wastewater treatment and green algae-to-biofuel technology
Algae require considerable amounts of water to grow and thrive. The organisms themselves are typically 80–85% water466 and the photosynthetic process results in the dissociation of roughly one mole of water per mole of CO2.467 This implies that approximately 5–10 kg of water are consumed per kg of dry algae biomass produced. In addition to water incorporated within the cell, most algae grow and reproduce in aqueous suspension. When algae blooms are observed, it appears that there are copious amounts of biomass; indeed a thin suspension of Chlorella contains 2 × 1010 individual cells per liter of water.466 However, the percentage of suspended solids is actually quite low, typically less than 0.5% wet biomass (0.1% dry). Thus, for every gram of dry algae biomass generated, more than a kilogram of noncellular water is required to produce and support it. Water not only provides a physical environment in which the algae live and reproduce, it also delivers nutrients, removes waste products, and acts as a thermal regulator. Unlike natural environments, mass cultivation systems require that the water be acquired, contained, circulated, and pumped to and between desired locations. All of these activities entail inputs of energy, both direct and indirect, and the amount of energy expended is tightly coupled to the volume of water involved. The volume of water involved depends upon system geometries, losses from the system, and most importantly, the ability to reclaim and reuse water. The latter is affected by the efficiency of the separation process, the quality of the return water, and the sensitivity of the specific culture to changes and/or impurities in the return water, including waste products introduced by the algae themselves.468 Microalgae also have a significant role in wastewater treatment plants.469 As indicated in Fig. 31, the algal biomass grown to recover nitrogen and phosphorus from wastewater can be utilized in several ways such as for a fertilizer or as a food source in its own right. In order for this biomass to replace traditional phosphorus fertilizers the harvesting, transportation, stability, application techniques and the proportion of phosphorus availability to crops must be considered.470 Unlike bacterial biomass from enhanced biological phosphorus removal systems which quickly re-releases stored phosphorus under anaerobic conditions, Powell471 showed that algal biomass can retain stored phosphorus for many days. Furthermore, with regard to its fertilizer potential, Mulbry et al.472 compared seedling growth using dried algal biomass to commercial fertilizer and both showed growth at comparable levels. However, overall these issues from harvest to application are currently quite poorly covered by the literature for both algae and macrophytes. | ||
| Fig. 31 Overview of options to utilize algae to recover nitrogen phosphorus from wastewater.470 | ||
4.1. Nutrients recovery from wastewater
One promising way to make algal biofuel production more cost-effective is to couple it with wastewater treatment.440 Furthermore, many of the algal species in wastewater treatment processes form large colonies (50–200 μm), and cell aggregation might be achieved through nutrient limitation or CO2 addition,473 which will further lower the cost of algae harvesting. However, knowledge of growing algae on wastewaters such as municipal wastewater for algae harvesting by self-sedimentation is still limited. Wastewaters derived from industrial, municipal and agricultural resources (e.g., animal manure) have been studied in terms of algae growth and nutrient removal efficiency.440,472–476 However, the nutrient removal efficiencies achieved did not meet increasingly stringent regulations and limits on wastewater discharge. Therefore, further exploration is required for improved wastewater treatment and cost-effective microalgae-based biofuel feedstock production.As mentioned above, microalgae harvesting employs some typical methods such as filtration, sedimentation, centrifugation, or flocculation, which can be technically and economically challenging for larger production scales. Macroalgae are multicellular and can be more easily harvested, either manually or mechanically, which may suggest that macroalgae is a better candidate for nutrient removal from aquatic environments. However, microalgae usually have a higher lipid productivity per cultivation area contributing to a greater potential for liquid fuel production (Table 1a). As macroalgae generally do not contain lipids and have high carbohydrate contents, they are more favored for biogas or alcohol-based fuels production. Table 1b shows the levels of the nitrogen and phosphorus in different wastewaters. Compared with animal wastewater, municipal wastewater has less nitrogen and phosphorus. However, there are often considerable amounts of heavy metals such as lead, zinc, and copper in raw municipal sewage. Selection of microalgae strains with a high metal sorption capacity is crucial to achieve high metal removal efficiency. So far, only a few algal species have been studied for their metal sorption ability. Compared with typical agricultural, municipal, and industrial wastewater, anaerobic digestion (AD) effluent has relatively lower carbon levels because microbial activity during the digestion converts the carbon to methane.477 The nitrogen in AD effluent is mainly in the form of ammonium.478 Dilution of AD effluent is usually needed before feeding to algae in order to avoid the potential inhibition of algal growth due to a high ammonium concentration and turbidity.479 In addition, as there is a significant amount of bacteria in AD effluent, proper pretreatment, such as filtration and autoclaving, may be necessary to prevent the contamination of algae production systems.477
| (a) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Algae category | Representative species | Composition (%w/w) | Lipid productivity [g (m2 d−1)] | Cultivation methods | Harvesting methods | Ref.(s) | ||
| Carbohydrates | Protein | Lipid | ||||||
| Microalgae | Scenedesmus obliquus | 10–17 | 50–56 | 12–14 | 2.4–13.5 | Open ponds; PBRs | Filtration; sedimentation; centrifugation; flocculation | 480 and 481 |
| Chlorella sp. | 12–17 | 51–58 | 14–22 | 1.6–16.5 | ||||
| Euglena gracilis | 14–18 | 39–61 | 22–38 | 7.7 | ||||
| Macroalgae | Laminaria sp.(brown seaweed) | 60 | 12 | 2 | 0.7–0.9 | Natural stocks; aquaculture | Manual; mechanization | 480 and 482 |
| Ulva sp. (green seaweed) | 23–78 | 10–33 | 0–6 | 0.6 | ||||
| (b) | |||||
|---|---|---|---|---|---|
| Wastewater category | Description | TN (mg L−1) | TP (mg L−1) | N/P | Ref.(s) |
| Municipal wastewater | Sewage | 15–90 | 5–20 | 3.3 | 483 |
| Animal wastewater | Dairy | 185–2636 | 30–727 | 3.6–7.2 | 484 and 485 |
| Poultry | 802–1825 | 50–446 | 4–16 | 485 and 486 | |
| Swine | 1110–3213 | 310–987 | 3.0–7.8 | 485 and 487 | |
| Beef feedlot | 63–4165 | 14–1195 | 2.0–4.5 | 485 and 486 | |
| Piggery | 56 | 13.5 | 4.1 | 488 | |
| Industrial wastewater | Textile | 21–57 | 1.0–9.7 | 2.0–4.1 | 489 and 490 |
| Winery | 110 | 52 | 2.1 | 491 | |
| Tannery | 273 | 21 | 13 | 492 | |
| Paper mill | 1.1–10.9 | 0.6–5.8 | 3.0–4.3 | 493 | |
| Olive mill | 532 | 182 | 2.9 | 494 | |
| Anaerobic digestion effluent | Dairy manure | 125–3456 | 18–250 | 7.0–13.8 | 477 and 495 |
| Poultry manure | 1380–1580 | 370–382 | 3.6–4.3 | 496 and 497 | |
| Sewage sludge | 427–467 | 134–321 | — | 498 | |
| Food waste and dairy manure | 1640–1885 | 296–302 | — | 499 | |
| (c) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Algae category | Genus & species | Waste stream | Process type | Removal time (d) | Total nitrogen (TN) | Total phosphorus (TP) | Ref.(s) | ||
| Initial conc. (mg L−1) | Removal efficiency (%) | Initial conc. (mg L−1) | Removal efficiency (%) | ||||||
| Chlorophyte | Chlorella sp. | Digested manure | Batch | 21 | 100–240 | 76–83 | 15–30 | 63–75 | 477 |
| C. kessleri | Artificial medium | Batch | 3 | 168 | 8–19 | 10–12 | 8–20 | 500 | |
| C. pyrenoidosa | Industrial wastewater | Fed-batch | 5 | 267 | 87–89 | 56 | 70 | 504 | |
| C. sorokiniana | Municipal wastewater | Batch | 10 | — | — | 22 | 45–72 | 507 | |
| C. vulgaris | Artificial medium | Batch | 1–10 | 13–410 | 23–100 | 5–8 | 46–94 | 505 | |
| C. vulgaris | Industrial wastewater | Batch | 5–9 | 3–36 | 30–95 | 112 | 20–55 | 503 | |
| C. vulgaris | Municipal wastewater | Batch | 2–10 | 49–1550 | 55–88 | 4–42 | 12–100 | 501 and 502 | |
| C. reinhardtii | Artificial medium | Batch | 10–30 | 129 | 42–83 | 120 | 13–14 | 516 | |
| Scenedesmus sp. | Artificial medium | Batch | 0.2–4.5 | 14–44 | 30–100 | 1.4–6.0 | 30–100 | 517 | |
| S. dimorphus | Industrial wastewater | Batch | 9 | — | — | 112 | 20–55 | 503 | |
| S. obliquus | Municipal wastewater | Batch | 0.2–8 | 27 | 79–100 | 12 | 47–98 | 501 and 519 | |
| Caynobacteria | Arthrospira sp. | Animal wastewater | Semi-cont. | — | — | 84–96 | — | 72–87 | 522 |
| A. platensis | Industrial wastewater | Batch | 15 | 2–3 | 96–100 | 18–21 | 87–99 | 509 | |
| Oscillatoria sp. | Municipal wastewater | Continuous | 14 | 498 | 100 | 76 | 100 | 510 | |
| Diatom | P. tricornutum | Municipal wastewater | Continuous | 14 | 498–835 | 80–100 | 76–116 | 50–100 | 507 and 508 |
| Haptophyte | I. galbana | Artificial medium | Batch | 8 | 377 | 99 | — | — | 512 |
| (d) | ||||
|---|---|---|---|---|
| Algae category | Algal biomass productivity (g m−2 d−1) | Daily removed per reactor volume (mg L−1 d−1) | Ref.(s) | |
| N | P | |||
| Phormidium sp. | 2.71 ± 0.7 | 3.66 ± 0.17 | 0.56 ± 0.07 | 513 |
| Chlamydomonas reinhardtii | 6.06 ± 1.2 | 6.39 ± 0.20 | 0.89 ± 0.06 | 513 |
| Chlorella vulgaris | 6.28 ± 0.8 | 4.39 ± 0.06 | 0.76 ± 0.09 | 513 |
| Scenedensmus rubescens | 6.56 ± 0.8 | 4.31 ± 0.18 | 0.60 ± 0.05 | 513 |
Chlorophytes is one of the largest phyla of microalgae, with a vast array of species and a wide geographical distribution. As shown in Table 1c, Chlorella sp. has been used in numerous studies and shown to be effective in removing nitrogen and phosphorus from different wastewater streams with a wide range of initial concentrations. Nitrogen and phosphorous removal efficiencies from the growth of Chlorella sp. range from 8% to 100%. Studies in Table 1c also confirm that C. vulgaris has higher nutrient removal efficiencies than that of Chlorella kessleri when comparing their performances in artificial media. An exceptionally low nutrient removal was found in the growth of C. kessleri in which the microalgae were subjected to artificial wastewater for a relatively small amount of time.500 In other studies, Chlorella sp. nitrogen removal efficiency was 23–100%, while phosphorus removal efficiency was 20–100%.477,501–506 In addition, it has been reported that Chlorella sp. is tolerant to NH4+–N.477
To utilize simultaneously both nitrogen and phosphorus, the N/P ratio should be with in a proper range. The optimal ratio differs among cultures due to strain-varying metabolic pathways. The N/P ratio can be up to 250 for healthy freshwater environments, but in most wastewater streams ratios may be as low as 4–5.511,514 An optimal N/P ratio for C. vulgaris was reported to be 7,515 in agreement with the N/P ratio of 7.2 calculated from the Stumm empirical formula for microalgae (C106H263O110N16P). These ratios indicate that the removal rate of nitrogen would be faster than that of phosphate, since a larger proportion is required. The faster removal of nitrogen over phosphorus was observed in the growth of Chlorella pyrenoidosa in soybean processing wastewater.504 It was observed that the removed nitrogen was mainly used for algal cell synthesis, whereas 17% of the phosphorus was removed via precipitation rather than by assimilation.
Some Chlorella species are heterotrophic or mixotrophic and can consume organic forms of carbon in addition to inorganic nutrients as part of their metabolic process. This can be an advantage when using wastewater streams containing carbon residues, such as digested dairy manure.477 Acetate, found in some wastewaters, was shown to be effectively consumed during heterotrophic or mixotrophic microalgae cultivation.516 Anaerobically pretreated soybean processing wastewater was shown to improve the growth of C. pyrenoidosa by means of providing additional acetate and small organic molecules.504 Heterotrophic growth is not an advantageous strategy in wastewaters deficient in organic carbon. Under heterotrophic conditions, the addition of carbon in the form of sodium acetate or glucose was necessary to achieve ammonium removal at a level equivalent to that under autotrophic conditions for the growth of C. Vulgaris.517 Another chlorophyte widely used for nutrient removal studies is Scenedesmus sp. (small non-motile green algae) often clustered in colonies consisting of 2, 4, 8, 16 or 32 cells. The cells are equipped with spines and bristles, which make the colonies more buoyant and allow increased light and nutrient uptake while deterring predation in the water. Table 1c shows that the nitrogen and phosphorus removal efficiency of Scenedesmus sp. was 30–100%. Its nutrient uptake behavior was not remarkably different from that of some Chlorella, i.e., Scenedesmus dimorphus versus C. Vulgaris.503 However, the removal of ammonium by S. dimporphus was significantly greater than that of C. vulgaris at an incubation time of less than 9 days (220 h); while immobilized in alginate, the ammonium removal efficiency of Scenedesmus obliquus was higher than that of C. Vulgaris.501 It was reported that Scenedesmus sp. requires an N/P ratio of approximately 30 to grow without limitation by either nutrient.518 When grown in an environment with N/P ratios between 12 and 18, the microalgae had a continuous nitrogen limitation, resulting in a high internal phosphate pool.519 Thus, the subsequent nitrogen removal rates were always shown to be greater than that of phosphorus. The high N/P ratio requirement could possibly explain the low phosphorus removal of 20–55% from agricultural wastewater by S. Dimorphus.503 Other genera of green algae are also capable of effectively removing nutrients from wastewater. Sawayama et al.520 found that Botryococcus braunii grown in treated sewage from municipal wastewater was able to consume nitrate and nitrite, but did not remove ammonium. Ammonium was reported to be inhibitory to cell growth in this particular culture. Chlamydomonas reinhardtii was capable of removing 42–55% of ammonium and 13–15% of phosphorus from an artificial medium with an N/P ratio of approximately 1.506 The removal efficiency was slightly increased when scaling up the process 45- or 90-fold in a biocoil reactor.506 Non-axenic cultures, which are a mixture of different algae species, can also be used to remove nutrients from wastewater. A combination of C. vulgaris, Scenedesmus falcatus, Chlamydomonas mirabilis, and Microcystis aeruginosa showed a 58% reduction in ammonium and 34% reduction in phosphates during the algal treatment phase of a city sewage treatment process.521 Table 1d shows the algal biomass productivity, N and P removal rates of the four different unicellular microalgae species. It was clearly observed that the three green microalgae C. reinhardtii, C. vulgaris and S. rubescens were suitable for integration of wastewater treatment and algae cultivation in terms of biomass settleability, nutrient removal rate and biomass productivity.
4.2. Integrated algae systems for biofuel production
Post-hydrothermal liquefaction wastewater (PHWW) is a high-strength wastewater that can accumulate most of the feedstock nutrients (80% or so) and some of the organics up to 40%,523 which provides a significant opportunity for nutrient and carbon recycling. PHWW recycled back to the algae culturing system can allow for multiple cycles of algae growth on each aliquot of incoming nutrients, which maximizes bioenergy production per unit of nutrient inputs. This approach has been investigated in recent studies using HTL wastewater524–526 and an earlier study suggested a similar approach but used a recondensed wastewater from gasification.527 These studies showed that nutrients in wastewaters from thermochemical conversion processes can be used for algae cultivation, but that significant dilution was required (50–500 times). However, these studies did not identify a viable and sustainable source of dilution water and raised other important questions about how this nutrient recycling can be incorporated into an algae biofuel production system. In Zhou et al.'s study,528 these issues were addressed in pursuit of an optimized system integrating algal wastewater treatment and bioenergy production including original process modeling to quantify the specific benefits of nutrient recycling and analyze the national implications for sustainable biofuel production. Specifically, this study investigated a novel integrated waste-to-energy system referred to Environment-Enhancing Energy (E2-Energy) that synergistically combines algal wastewater treatment with large-scale bioenergy production via HTL as shown in Fig. 32. | ||
| Fig. 32 Simplified schematic of the Environment-Enhancing Energy (E2-Energy) process for integrated wastewater treatment and biofuel production. | ||
In the proposed E2-Energy system, wastewaters from a variety of sources (e.g., municipal, livestock, food processing) can be initially separated into a concentrated biosolids fraction and a dilute liquid fraction by common physicochemical processes (e.g., sedimentation, filtration). Then, mixed cultures of low-lipid, fast-growing algae and bacteria are cultivated in a combination of the dilute liquid wastewater fraction and recycled PHWW (Key Step 1). As the algae and bacteria grow symbiotically, the wastewater is treated by providing removal of organics and nutrients (Key Step 2). Note that the energy input for aerobic breakdown of wastewater contaminants is reduced by the oxygen provided during algal photosynthesis. Subsequently, the mixed culture biomass is harvested, and combined with the concentrated biosolids separated from the initial wastewater. This mixture is then fed into a HTL reactor for biofuel production (Key Step 3). The HTL process also generates a CO2-rich gaseous product and strong wastewater with re-released organics/nutrients (Key Step 4), which is recycled back to the algal–bacterial cultivation system for reuse. This recycling can repeat again and again over multiple cycles of algal growth, harvesting and biofuel conversion that leverages the nutrient content of wastewater into maximum bioenergy quantities, which can be many times the original wastewater energy content. The E2-Energy system elegantly has resolved several key bottlenecks in other current approaches to algal biofuel production, and provides significant environmental benefits, including carbon capture and wastewater nutrient removal. This novel approach embodies a significant paradigm change and brings together two important goals of modern society-“energy production” and “environmental protection”-into a complementary relationship, whereas historically these goals have most often been antagonistic. This harmonious combination is critically important for addressing the major challenges of a growing world population including energy security, climate change, quality of water resources and sustainable development.529
Some microalgae species can grow in a photoautotrophic mode (PM) under light, or in a heterotrophic mode (HM) in the presence of organic carbon or in a mixotrophic culture mode (MM) when supplied with both organic and inorganic carbon under light/dark conditions.530 Many different cultivation strategies have been developed to explore the potential of algae as a feedstock for biofuel, metabolites and other high-value bio-products based on these growth modes. Oyler531 developed a process of sequential photoautotrophic and heterotrophic growth (PHM) for algal biofuel production. Das et al.530 studied a phototrophic–mixotrophic two phase culture model (PMM) for algae-based biodiesel production using glycerol, glucose and sucrose as organic carbon. Xiong et al.532 developed a similar photoautotrophic-heterotrophic culture mode (PHM) for high algal cell density production. Furthermore, developing a hetero-photoautotrophic culture mode (HPM) to effectively couple treatment of organic-rich wastewater, such as concentrated municipal wastewater (CMW), with low-cost biofuel production has been also reported by Zhou et al.533 The success of such an algae-based treatment system for organic-rich wastewater relies on the ability of the algal cells to effectively assimilate both organic carbons (heterotrophic growth) and nutrients, such as N and P, from wastewater and inorganic carbon (e.g., CO2) from flue gas for maximal algal biomass and lipid production and efficient nutrient removal as well as CO2 sequestration.440,534–536 The locally isolated strain Auxenochlorella protothecoides UMN280 (ref. 476) is facultative heterotrophic and adapts well to CMW. In Zhou's study, a biological system was utilized in order to treat municipal wastewater and the sludge generated at the Metro plant are dewatered using centrifuges and then is combusted in fluid bed incinerators equipped with heat recovery boilers.533 The process requires no additional inputs of fuel and creates heat and electricity for the facility buildings. The CO2-rich flue gas during combustion could be sequestered by sparging into an algae bioreactor and the electricity produced could be used to run the bioreactor and harvest the algae as well as to convert algae to refined bio-oil directly by thermochemical processes537 or to create biodiesel by transesterification. Therefore, the integrated process could be incorporated into the typical Metro plant municipal wastewater treatment to achieve significant cost reductions of algae-based biofuel (Fig. 33).
 | ||
| Fig. 33 Integration of Pilot-scale HMP algae cultivation system into metro plant municipal wastewater process flow.532 | ||
Integrated algal systems can be employed for wastewater treatment and bioremediation to capture carbon, nitrogen and phosphorus from the specialty industrial, municipal and agriculture wastes (Fig. 34). A Green Wisdom Inc. Plans (G-WISP) in Arkansas has been developed to implement an integrated algal production system to recycle agricultural wastes (i.e., cotton plants) for biofuel. The future communities can create processing facilities that support algae production as their common core goal by developing local sustainable systems, like G-WISP. It means that using existing crops or rotational crops, which are desirable for their waste-to-energy value. Using local, multiple mix agricultural waste and alternative technologies in near closed-loop systems, these communities can create cost effectiveness and produce jobs. Algal production, combining and integrating alternative energy technologies, will foster synergistic development supporting a self-sustaining community system. This system uses anaerobically digested agricultural waste materials including catfish processing waste to feed algal cultures. It shows multiple environmental abatements of CO2 (e.g., from the anaerobic digester), and reclamation and use of other waste streams, such as water and heat (e.g., from the digester and co-generation), to support optimum sustainable algal growth. Furthermore, this system demonstrates how near-closed-loop sustainable systems can create products, algal oil and methane energy, and by-products (e.g., fertilizer), and even multiple resulting business spin-offs such as companies to help market or distribute the products and byproducts, which, in turn, create jobs for communities of the future. This sustainable community plan would insure rural revitalization and, ultimately, global economic development, while curbing the dependence of US on fossil fuels.538 Although nitrogen and phosphorous are elements key to algal growth, they are serious pollutants in many waterways. Algae can thrive in nitrogen- and phosphorus-rich conditions common to many wastewaters,381,539,540 and this feature may be harnessed to not only remove,540 but also capture these important nutrients with the aim of returning them to the terrestrial environment as agricultural fertilizer, providing a high value by-product for algae that are primarily being grown for biofuel.
 | ||
| Fig. 34 Schematics of an integrated algal culture system for bioremediation and biofuel production. | ||
From an environmental point of view, microalgae culture systems should be studied to capture CO2 and consume the nutrients in wastewaters, simultaneously. From an engineering point of view, the costs associated with all different processes should be reduced. For instance, harvesting and dewatering are processes with high energy requirements mainly because of small cell size and low cell biomass levels in microalgal cultures; thus, research efforts should be performed to achieve high cell densities. This limitation is related to the access of the cells to gas and light. Air-lift bioreactors with light distribution through optical fibers (increasing the ratio between the illumination surface and reactor volume) and membrane integrated microalgal cultivation processes may resolve these kinds of problems. Apart from the advances in PBR engineering, the application of biorefinery concepts (to exploit the full potential of commercial products derived from microalgal biomass) can make this CO2 capture process economically feasible.541,542
5. Concluding remarks and outlook
The high growth rates, reasonable growth densities and high oil contents have all been the advantages based on investing significant capital to turn algae into biofuels. The algal biofuels production chain shows that the major challenges, including strain isolation, nutrient sourcing and utilization, production management, harvesting, co-product development, fuel extraction, refining and residual biomass utilization. Improved engineering will make a significant impact on algae biofuel production. There are important challenges for engineers and biologists to either design cheap PBRs for large-scale deployment, or to combine forces to develop species that grow efficiently in low-cost open systems. PBRs have advantages over open systems as they can more easily maintain axenic cultures and controlled growth environments, which may lead to an increase in productivity and a decrease in contamination; however, contained systems are challenged by efficiencies of gas exchange and a requirement for supplemental cooling. Regardless of the growth strategy employed, substantial improvements over current technologies for the growth, harvesting and extracting oil from algae need to be made, and coordinated efforts will be needed to couple engineering advances with improved production strains. Oil extraction is another challenge. There are three major strategies (i.e., oil press/expeller, hexane extraction, and supercritical CO2 fluid extraction) for extracting oil from algae. These technologies have been successfully demonstrated but are relatively expensive, either in terms of equipment needed or energy required to extract the oil.543 Therefore, large-scale cultivation of algae for biodiesel production is still in the research and development phase. The long term potential of this technology can be improved by the following approaches:544 (1) cost saving growth technologies of oil-rich algae should be identified and developed; (2) integrated bio-refineries can be used to produce biodiesel, animal feed, biogas and electrical power thereby reducing the cost of production; (3) enhancing algal biology by genetic modification and metabolic engineering has the greatest impact on improving the economics of microalgae biodiesel; (4) area efficient techniques to capture CO2 from industrial power plants need to be identified; (5) recycling of nutrients from municipal sewage and industrial wastewaters are required to reduce the demand of fertilizers to grow algae; (6) economics of microalgae production can be improved by additional revenues from wastewater treatment and greenhouse gas emissions abatement.Algae can be grown in many ways in freshwater, saltwater or wastewater; in closed PBRs or open ponds. One key advantage of algae is that its cultivation does not require cropland. However, other resources are needed, and the amounts of these resources vary widely from one algae production pathway to another. For instance, it was reported that between 3.15 and 3650 liters of freshwater are needed to produce the algal biofuel equivalent to 1 liter of gasoline using current technologies. For comparison, 5 to 2140 liters of water are needed to produce a liter of corn ethanol and 1.9 to 6.6 liters are required to produce a liter of petroleum-based gasoline. The national research council report notes that none of the sustainability concerns will be a definitive barrier to future production of algal biofuels, significant biological and engineering innovations are needed to mitigate demands on resources.545,546 Thus, the integration of upstream production and downstream processing of microalgae, and the framing of these in the context of water savings and net energy gain is required to build up credibility and withstand scrutiny. Otherwise, microalgae biofuels could go from ‘hero to zero’ in a very short space of time in this age of advanced communications.546 The latest research indicates that biomass impregnated into seawater (saltwater, i.e. MgCl2). Then, the MgCl2 preloaded biomass can be fabricated into the mesoporous carbon stabilized MgO nanoparticles for highly efficient CO2 capture.372 Thus, if microalge are grown in seawater, it has one possibility that the solid products containing amounts of alkaline or alkaline earth metallic salts can be synthesized into the value-added mesoporous carbon materials for CO2 capture. There is an increasing emphasis on ensuring that bio-based products do not have negative effects on the natural environment and, as such, it is crucial that any issues surrounding the environmental impacts of biofuels, bioenergy and commodity chemicals production are addressed prior to the commercialization of products. Among biofuel feedstocks, algae can hold the promise to offset much or all of our fossil fuels utilization. While many of the outstanding challenges are daunting, there are many reasons to be optimistic. Investment in research and development has been steadily increasing, and new multi-stakeholder collaborations bode well for innovation. The further development of co-products for algal fuels will help increase the likelihood of success. The criteria for which chemicals are most promising as value-added algal biorefinery co-products would be scalability, demand and, most importantly, raw materials. Algal biomass serving as the feedstocks for chemical co-products is likely to have a unique and somewhat tunable chemical composition compared with traditional plants. The absence of lignin, the presence of phospholipids and the unique carbohydrate fractions of algae, as well as the variability between and within algal species, will require new product platforms and technological adaptations beyond those currently realized in conventional biorefineries. However, these challenges can easily be viewed as opportunities. The biorefinery is an ideal setting for innovation, and the creativity of the green chemistry and green engineering community with respect to biomass transformations would be critical in improving the future prospects for our energy and material economy.
Up to now, many microalgae projects can achieve maximal lipid yields only under stress conditions hindering growth and providing compositions not ideal for biofuel applications. Metabolic engineering of algal fatty acid biosynthesis promises to create strains capable of economically producing fungible and sustainable biofuels. The algal fatty acid biosynthetic pathway has been deduced by homology to bacterial and plant systems, and much of our understanding is gleaned from basic studies from these systems. However, successful engineering of lipid metabolism in algae will necessitate a thorough characterization of the algal fatty acid synthase including protein–protein interactions and regulation. Thus, many efforts have been made for improving the engineering of fatty acid biosynthesis toward optimizing microalgae as a biodiesel feedstock. Algal bioresource generation can be integrated with human communities to form a sustainable permaculture ecosystem, or an algae-based bioresource cycle. Local algae species are sourced and studied from ‘nature's culture collection’ for bioresource production. Algal farmers can utilize locally available waste resources (e.g., wastewaters, CO2 and heat) to cultivate a desired native algal biomass, which is harvested and processed at an algae-based biorefinery into consumable products. Algal cultivation integrated with algae-based biorefineries can yield a diversity of bioresources (biodiesel, green gasoline, bio-jet fuel, isolated proteins, food starches, textiles, organic fertilizers, etc.), which mitigate the cost of biofuel production. For example, the alga could be an indigenous variety of Chlorella that is grown on local nutrients from municipal wastewater treatment plant effluent and captures CO2 derived from nearby sources such as the combustion of fossil fuels, fermentation and industrial facilities, cement plants, landfill gas, or biogas from anaerobic digestion. Algal biomass produces lipids, proteins or starches that could be processed into biodiesel, nutritional supplements, and food products. The organic residuals produced during processing and after consumption can be anaerobically digested to produce biogas (methane and CO2) and solubilized mineral nutrients. The CO2 and the nutrients can be reused directly by the algal culture, avoiding the costs associated with supplying these external inputs. In addition to community use as a renewable fuel, the methane can provide energy for on-site processing, including harvesting, drying, heating, or mixing the algal culture. Utilizing the energy, nutrients and CO2 held within residual waste materials to provide all necessary inputs except for sunlight, the cultivation of algae becomes a closed-loop engineered ecosystem. Developing this biotechnology is a tangible step towards a waste-free sustainable society. Significantly, utilizing industrial wastewaters for algae cultivation, the biological effects of the emergent pollutants (i.e., engineered nanoparticles, high-concentration heavy metal) to aquatic ecosystems should be evaluated.
Acknowledgements
The authors would like to thank the Chinese Scholarship Council (CSC) for the financial support under grant no. 201206230168.References
- J. Barber, Chem. Soc. Rev., 2009, 38, 185–196 RSC.
- J. Barber and P. D. Tran, J. R. Soc., Interface, 2013, 10, 20120984 CrossRef PubMed.
- M. I. Hoffert, K. Caldeira, A. K. Jain, E. F. Haites, L. D. Harvey, S. D. Potter, M. E. Schlesinger, S. H. Schneider, R. G. Watts and T. M. Wigley, Nature, 1998, 395, 881–884 CrossRef CAS.
- International Energy Agency, World Energy Outlook 2012, Paris, France, 2012, p. 16 Search PubMed.
- N. Nakicenovic, J. Alcamo, G. Davis, B. de Vries, J. Fenhann, S. Gaffin, K. Gregory, A. Grubler, T. Y. Jung and T. Kram, Special report on emissions scenarios: a special report of Working Group III of the Intergovernmental Panel on Climate Change, Pacific Northwest National Laboratory, Environmental Molecular Sciences Laboratory (US), Richland, WA (US), 2000 Search PubMed.
- N. S. Lewis and D. G. Nocera, Proc. Natl. Acad. Sci. U. S. A., 2006, 103, 15729–15735 CrossRef CAS PubMed.
- J. Goldemberg and T. B. Johansson, World Energy Assessment: Overview: 2004 Update, United Nations Publications, 2004 Search PubMed.
- R. K. Pachauri, Climate change 2007, Synthesis report, Contribution of Working Groups I, II and III to the fourth assessment report, 2008 Search PubMed.
- B. Metz, Carbon Dioxide Capture and Storage: Special Report of the Intergovernmental Panel on Climate Change, Cambridge University Press, 2005 Search PubMed.
- N. S. Lewis and G. Crabtree, 2005, http://resolver.caltech.edu/CaltechAUTHORS:LEWsolarenergyrpt05.
- S. E. Shaheen, D. S. Ginley and G. E. Jabbour, MRS Bull., 2005, 30, 10–19 CrossRef CAS.
- D. P. Hagberg, J.-H. Yum, H. Lee, F. De Angelis, T. Marinado, K. M. Karlsson, R. Humphry-Baker, L. Sun, A. Hagfeldt and M. Grätzel, J. Am. Chem. Soc., 2008, 130, 6259–6266 CrossRef CAS PubMed.
- S. Kim, J. K. Lee, S. O. Kang, J. Ko, J.-H. Yum, S. Fantacci, F. De Angelis, D. Di Censo, M. K. Nazeeruddin and M. Grätzel, J. Am. Chem. Soc., 2006, 128, 16701–16707 CrossRef CAS PubMed.
- J.-R. Petit, J. Jouzel, D. Raynaud, N. Barkov, J.-M. Barnola, I. Basile, M. Bender, J. Chappellaz, M. Davis and G. Delaygue, Nature, 1999, 399, 429–436 CrossRef CAS.
- J. T. Houghton, Y. Ding, D. J. Griggs, M. Noguer, P. J. van der Linden, X. Dai, K. Maskell and C. Johnson, Climate change 2001: the scientific basis, Cambridge University Press Cambridge, 2001 Search PubMed.
- B. C. O'Neill and M. Oppenheimer, Science, 2002, 296, 1971–1972 CrossRef PubMed.
- T. P. Hughes, A. H. Baird, D. R. Bellwood, M. Card, S. R. Connolly, C. Folke, R. Grosberg, O. Hoegh-Guldberg, J. Jackson and J. Kleypas, Science, 2003, 301, 929–933 CrossRef CAS PubMed.
- D. A. Stainforth, T. Aina, C. Christensen, M. Collins, N. Faull, D. Frame, J. Kettleborough, S. Knight, A. Martin and J. Murphy, Nature, 2005, 433, 403–406 CrossRef CAS PubMed.
- Q. Schiermeier, Nature, 2005, 435, 732–733 CrossRef CAS PubMed.
- K. Miyamoto, Renewable biological systems for alternative sustainable energy production, Food & Agriculture Org., 1997 Search PubMed.
- K. N. Ferreira, T. M. Iverson, K. Maghlaoui, J. Barber and S. Iwata, Science, 2004, 303, 1831–1838 CrossRef CAS PubMed.
- S. Iwata and J. Barber, Curr. Opin. Struct. Biol., 2004, 14, 447–453 CrossRef CAS PubMed.
- A. Mishra, W. Wernsdorfer, K. A. Abboud and G. Christou, Chem. Commun., 2005, 54–56 RSC.
- J. Biesiadka, B. Loll, J. Kern, K.-D. Irrgang and A. Zouni, Phys. Chem. Chem. Phys., 2004, 6, 4733–4736 RSC.
- O. Kruse, J. Rupprecht, J. H. Mussgnug, G. C. Dismukes and B. Hankamer, Photochem. Photobiol. Sci., 2005, 4, 957–970 CAS.
- W. Van Camp, Curr. Opin. Biotechnol., 2005, 16, 147–153 CrossRef CAS PubMed.
- Z. P. Jing, F. Gallardo, M. B. Pascual, R. Sampalo, J. Romero, D. Navarra, A. Torres and F. M. Cánovas, New Phytol., 2004, 164, 137–145 CrossRef CAS.
- A. G. Good, A. K. Shrawat and D. G. Muench, Trends Plant Sci., 2004, 9, 597–605 CrossRef CAS PubMed.
- A. E. Farrell, R. J. Plevin, B. T. Turner, A. D. Jones, M. O'hare and D. M. Kammen, Science, 2006, 311, 506–508 CrossRef CAS PubMed.
- M. Wang, C. Saricks and D. Santini, 1999, http://www.transportation.anl.gov/pdfs/TA/58.pdf.
- M. Wang, Updated energy and greenhouse gas emission results of fuel ethanol, San Diego, 2005 Search PubMed.
- M. Isaias, M. L. V. Leal and J. A. R. da Silva, PDF, Secretariat of the Environment, Government of the State of São Paulo, http://www.eners.ch/plateforme/medias/macedo_2004.pdf, retrieved on, 2008, pp. 05–09.
- L. Di Lucia and L. J. Nilsson, Transport Pol., 2007, 14, 533–543 CrossRef.
- D. Tilman, R. Socolow, J. A. Foley, J. Hill, E. Larson, L. Lynd, S. Pacala, J. Reilly, T. Searchinger and C. Somerville, Science, 2009, 325, 270–271 CrossRef CAS PubMed.
- A. K. Agarwal, Prog. Energy Combust. Sci., 2007, 33, 233–271 CrossRef CAS.
- A. Demirbas, Prog. Energy Combust. Sci., 2007, 33, 1–18 CrossRef CAS.
- S. Atsumi, T. Hanai and J. C. Liao, Nature, 2008, 451, 86–89 CrossRef CAS PubMed.
- J. P. Scharlemann and W. F. Laurance, Science, 2008, 319, 43–44 CrossRef CAS PubMed.
- G. P. Robertson, V. H. Dale, O. C. Doering, S. P. Hamburg, J. M. Melillo, M. M. Wander and W. Parton, Science, 2008, 322, 49–50 CrossRef CAS PubMed.
- G. Stephanopoulos, Science, 2007, 315, 801–804 CrossRef CAS PubMed.
- M. E. Himmel, S.-Y. Ding, D. K. Johnson, W. S. Adney, M. R. Nimlos, J. W. Brady and T. D. Foust, Science, 2007, 315, 804–807 CrossRef CAS PubMed.
- D. Tilman, J. Hill and C. Lehman, Science, 2006, 314, 1598–1600 CrossRef CAS PubMed.
- J. Hill, E. Nelson, D. Tilman, S. Polasky and D. Tiffany, Proc. Natl. Acad. Sci. U. S. A., 2006, 103, 11206–11210 CrossRef CAS PubMed.
- A. J. Ragauskas, C. K. Williams, B. H. Davison, G. Britovsek, J. Cairney, C. A. Eckert, W. J. Frederick, J. P. Hallett, D. J. Leak and C. L. Liotta, Science, 2006, 311, 484–489 CrossRef CAS PubMed.
- J. M. Melillo, J. M. Reilly, D. W. Kicklighter, A. C. Gurgel, T. W. Cronin, S. Paltsev, B. S. Felzer, X. Wang, A. P. Sokolov and C. A. Schlosser, Science, 2009, 326, 1397–1399 CrossRef CAS PubMed.
- P. Fairley, Nature, 2011, 474, S2–S5 CrossRef CAS PubMed.
- A. Melis, Energy Environ. Sci., 2012, 5, 5531–5539 CAS.
- A. Melis, Curr. Opin. Chem. Biol., 2013, 17, 453–456 CrossRef CAS PubMed.
- S. H. Desai and S. Atsumi, Curr. Opin. Biotechnol., 2013, 14(6), 1031–1036 CrossRef PubMed.
- P. J. Crutzen, A. R. Mosier, K. A. Smith and W. Winiwarter, Atmos. Chem. Phys., 2008, 8, 389–395 CrossRef CAS.
- Y.-X. Huo, D. G. Wernick and J. C. Liao, Curr. Opin. Biotechnol., 2012, 23, 406–413 CrossRef CAS PubMed.
- J. Fargione, J. Hill, D. Tilman, S. Polasky and P. Hawthorne, Science, 2008, 319, 1235–1238 CrossRef CAS PubMed.
- T. Searchinger, R. Heimlich, R. A. Houghton, F. Dong, A. Elobeid, J. Fabiosa, S. Tokgoz, D. Hayes and T.-H. Yu, Science, 2008, 319, 1238–1240 CrossRef CAS PubMed.
- H. Michel, Angew. Chem., Int. Ed. Engl., 2012, 51, 2516–2518 CrossRef CAS PubMed.
- M. D. Archer and J. Barber, Molecular to global photosynthesis, 2004, vol. 2, pp. 1–42 Search PubMed.
- P. G. Falkowski and J. A. Raven, Aquatic photosynthesis, Blackwell Science Malden, MA, 1997 Search PubMed.
- J. Pickett, D. Anderson, D. Bowles, T. Bridgwater, P. Jarvis, N. Mortimer, M. Poliakoff and J. Woods, Sustainable biofuels: prospects and challenges, The Royal Society, London, UK, 2008 Search PubMed.
- T. M. Mata, A. A. Martins and N. S. Caetano, Renewable Sustainable Energy Rev., 2010, 14, 217–232 CrossRef CAS.
- E. Farrell, M. Bustard, S. Gough, G. McMullan, P. Singh, D. Singh and A. McHale, Bioprocess Eng., 1998, 19, 217–219 CAS.
- L. Brennan and P. Owende, Renewable Sustainable Energy Rev., 2010, 14, 557–577 CrossRef CAS.
- H. Schuhmann, D. K. Lim and P. M. Schenk, Biofuels, 2012, 3, 71–86 CrossRef CAS.
- O. Pulz and K. Scheibenbogen, in Bioprocess and algae reactor technology, apoptosis, Springer, 1998, pp. 123–152 Search PubMed.
- Y. Wang, H. Wu and M. Zong, Bioresour. Technol., 2008, 99, 7232–7237 CrossRef CAS PubMed.
- L. M. Brown, Energy Convers. Manage., 1996, 37, 1363–1367 CrossRef CAS.
- H. Hsueh, H. Chu and S. Yu, Chemosphere, 2007, 66, 878–886 CrossRef CAS PubMed.
- I. E. Huertas, B. Colman, G. S. Espie and L. M. Lubian, J. Phycol., 2000, 36, 314–320 CrossRef.
- B. Colman and C. Rotatore, Plant, Cell Environ., 1995, 18, 919–924 CrossRef CAS.
- I. S. Suh and C.-G. Lee, Biotechnol. Bioprocess Eng., 2003, 8, 313–321 CrossRef CAS.
- Q. Hu, M. Sommerfeld, E. Jarvis, M. Ghirardi, M. Posewitz, M. Seibert and A. Darzins, Plant J., 2008, 54, 621–639 CrossRef CAS PubMed.
- V. Patil, K.-Q. Tran and H. R. Giselrød, Int. J. Mol. Sci., 2008, 9, 1188–1195 CrossRef CAS PubMed.
- X. Miao and Q. Wu, Bioresour. Technol., 2006, 97, 841–846 CrossRef CAS PubMed.
- P. S. Nigam and A. Singh, Prog. Energy Combust. Sci., 2011, 37, 52–68 CrossRef CAS.
- E. Waltz, Nat. Biotechnol., 2009, 27, 15–18 CrossRef CAS PubMed.
- J. Chappell, Annu. Rev. Plant Biol., 1995, 46, 521–547 CrossRef CAS.
- D. J. McGarvey and R. Croteau, Plant Cell, 1995, 7, 1015 CrossRef CAS PubMed.
- F. Cunningham, Jr and E. Gantt, Annu. Rev. Plant Biol., 1998, 49, 557–583 CrossRef PubMed.
- P. Lindberg, S. Park and A. Melis, Metab. Eng., 2010, 12, 70–79 CrossRef CAS PubMed.
- E. W. Lusas, in Kent and Riegel's Handbook of Industrial Chemistry and Biotechnology, Springer, 2007, pp. 1549–1656 Search PubMed.
- K. Hill, Pure Appl. Chem., 2000, 72, 1255–1264 CrossRef CAS.
- K. Hill, Pure Appl. Chem., 2007, 79, 1999–2011 CrossRef CAS.
- U. Schörken and P. Kempers, Eur. J. Lipid Sci. Technol., 2009, 111, 627–645 CrossRef.
- P. M. Foley, E. S. Beach and J. B. Zimmerman, Green Chem., 2011, 13, 1399–1405 RSC.
- K. Hussain, K. Nawaz, A. Majeed and L. Feng, World Appl. Sci. J., 2010, 9, 1313–1323 CAS.
- R. W. Evans and M. Kates, Arch. Microbiol., 1984, 140, 50–56 CrossRef CAS.
- S. Renaud, D. Parry and L.-V. Thinh, J. Appl. Phycol., 1994, 6, 337–345 CrossRef CAS.
- L. Gouveia, A. E. Marques, T. L. da Silva and A. Reis, J. Ind. Microbiol. Biotechnol., 2009, 36, 821–826 CrossRef CAS PubMed.
- Z. Cohen, Chemicals from microalgae, CRC Press, 1999 Search PubMed.
- S. Tatulian and G. Cevc, Phospholipids Handbook, Marcel Dekker, Inc, New York, 1993 Search PubMed.
- H. Zoebelein, Dictionary of renewable resources, Wiley-VCH Verlag GmbH, 2001 Search PubMed.
- J. V. Gerpen, Fuel Process. Technol., 2005, 86, 1097–1107 CrossRef.
- T. L. Alleman, Lipid Technol., 2008, 20, 40–42 CrossRef CAS.
- W. van Nieuwenhuyzen and M. C. Tomás, Eur. J. Lipid Sci. Technol., 2008, 110, 472–486 CrossRef CAS.
- E. D. Larson, Sustainable bioenergy: A framework for decision makers, UN-Energy, 2007 Search PubMed.
- A. David, Energy Environ. Sci., 2009, 2, 343–346 Search PubMed.
- S. Y. Lee and E. T. Papoutsakis, Metabolic engineering, CRC Press, 1999 Search PubMed.
- J. Lü, C. Sheahan and P. Fu, Energy Environ. Sci., 2011, 4, 2451–2466 Search PubMed.
- Y. Chisti, Biotechnol. Adv., 2007, 25, 294–306 CrossRef CAS PubMed.
- Y. Chisti, Trends Biotechnol., 2008, 26, 126–131 CrossRef CAS PubMed.
- S. L. Nielsen, S. Enriquez, C. Duarte and K. Sand-Jensen, Funct. Ecol., 1996, 167–175 CrossRef.
- D. Parkhurst and T. Givnish, Internal leaf structure: a three-dimensional perspective, 1986 Search PubMed.
- Y. Chisti, Trends Biotechnol., 2008, 26, 126–131 CrossRef CAS PubMed.
- M. Gavrilescu and Y. Chisti, Biotechnol. Adv., 2005, 23, 471–499 CrossRef CAS PubMed.
- J. Gressel, Plant Sci., 2008, 174, 246–263 CrossRef CAS.
- L. R. Lynd, M. S. Laser, D. Bransby, B. E. Dale, B. Davison, R. Hamilton, M. Himmel, M. Keller, J. D. McMillan and J. Sheehan, Nat. Biotechnol., 2008, 26, 169–172 CrossRef CAS PubMed.
- H. Alper and G. Stephanopoulos, Nat. Rev. Microbiol., 2009, 7, 715–723 CrossRef CAS PubMed.
- B. Dien, M. Cotta and T. Jeffries, Appl. Microbiol. Biotechnol., 2003, 63, 258–266 CrossRef CAS PubMed.
- B. C. Chu and H. Lee, Biotechnol. Adv., 2007, 25, 425–441 CrossRef CAS PubMed.
- F. Torney, L. Moeller, A. Scarpa and K. Wang, Curr. Opin. Biotechnol., 2007, 18, 193–199 CrossRef CAS PubMed.
- E. T. Johnson and C. Schmidt-Dannert, Trends Biotechnol., 2008, 26, 682–689 CrossRef CAS PubMed.
- S. Atsumi, W. Higashide and J. C. Liao, Nat. Biotechnol., 2009, 27, 1177–1180 CrossRef CAS PubMed.
- L. L. Beer, E. S. Boyd, J. W. Peters and M. C. Posewitz, Curr. Opin. Biotechnol., 2009, 20, 264–271 CrossRef CAS PubMed.
- J. Sheehan, Nat. Biotechnol., 2009, 27, 1128–1129 CrossRef CAS PubMed.
- J. Mata-Alvarez, S. Mace and P. Llabres, Bioresour. Technol., 2000, 74, 3–16 CrossRef CAS.
- Y. Chisti, Trends Biotechnol., 2003, 21, 89–93 CrossRef CAS PubMed.
- C. E. Wyman, Trends Biotechnol., 2007, 25, 153–157 CrossRef CAS PubMed.
- D. Antoni, V. V. Zverlov and W. H. Schwarz, Appl. Microbiol. Biotechnol., 2007, 77, 23–35 CrossRef CAS PubMed.
- J. N. Reeve, Annu. Rev. Microbiol., 1992, 46, 165–191 CrossRef CAS PubMed.
- Y. Chisti, Biofuels, 2010, 1, 233–235 CrossRef CAS.
- D. R. Ort, X. Zhu and A. Melis, Plant Physiol., 2011, 155, 79–85 CrossRef CAS PubMed.
- R. E. Blankenship, D. M. Tiede, J. Barber, G. W. Brudvig, G. Fleming, M. Ghirardi, M. Gunner, W. Junge, D. M. Kramer and A. Melis, Science, 2011, 332, 805–809 CrossRef CAS PubMed.
- V. H. Work, S. D'Adamo, R. Radakovits, R. E. Jinkerson and M. C. Posewitz, Curr. Opin. Biotechnol., 2012, 23, 290–297 CrossRef CAS PubMed.
- T. T. Y. Doan, B. Sivaloganathan and J. P. Obbard, Biomass Bioenergy, 2011, 35, 2534–2544 CrossRef CAS.
- B. R. Green and W. W. Parson, Light-harvesting antennas in photosynthesis, Springer, 2003 Search PubMed.
- G. D. Scholes, Annu. Rev. Phys. Chem., 2003, 54, 57–87 CrossRef CAS PubMed.
- S. Jang, M. D. Newton and R. J. Silbey, J. Phys. Chem. B, 2007, 111, 6807–6814 CrossRef CAS PubMed.
- Y.-C. Cheng and G. R. Fleming, Annu. Rev. Phys. Chem., 2009, 60, 241–262 CrossRef CAS PubMed.
- R. van Grondelle and V. I. Novoderezhkin, Phys. Chem. Chem. Phys., 2006, 8, 793–807 RSC.
- G. S. Engel, T. R. Calhoun, E. L. Read, T.-K. Ahn, T. Mančal, Y.-C. Cheng, R. E. Blankenship and G. R. Fleming, Nature, 2007, 446, 782–786 CrossRef CAS PubMed.
- H. Lee, Y.-C. Cheng and G. R. Fleming, Science, 2007, 316, 1462–1465 CrossRef CAS PubMed.
- I. P. Mercer, Y. C. El-Taha, N. Kajumba, J. P. Marangos, J. W. Tisch, M. Gabrielsen, R. J. Cogdell, E. Springate and E. Turcu, Phys. Rev. Lett., 2009, 102, 057402 CrossRef PubMed.
- R. P. Feynman, Rev. Mod. Phys., 1948, 20, 367–387 CrossRef.
- E. Collini, C. Y. Wong, K. E. Wilk, P. M. Curmi, P. Brumer and G. D. Scholes, Nature, 2010, 463, 644–647 CrossRef CAS PubMed.
- G. Richards, K. Wilk, P. Curmi, H. Quiney and J. Davis, J. Phys. Chem. Lett., 2012, 3, 272–277 CrossRef CAS.
- D. M. Jonas, Annu. Rev. Phys. Chem., 2003, 54, 425–463 CrossRef CAS PubMed.
- T. Brixner, T. Mančal, I. V. Stiopkin and G. R. Fleming, J. Chem. Phys., 2004, 121, 4221–4236 CrossRef CAS PubMed.
- M. Cho, Chem. Rev., 2008, 108, 1331–1418 CrossRef CAS PubMed.
- D. Abramavicius, B. Palmieri, D. V. Voronine, F. Šanda and S. Mukamel, Chem Rev., 2009, 109, 2350 CrossRef CAS PubMed.
- P. J. McGinn, K. E. Dickinson, S. Bhatti, J.-C. Frigon, S. R. Guiot and S. J. O'Leary, Photosynth. Res., 2011, 109, 231–247 CrossRef CAS PubMed.
- A. Kumar, S. Ergas, X. Yuan, A. Sahu, Q. Zhang, J. Dewulf, F. X. Malcata and H. Van Langenhove, Trends Biotechnol., 2010, 28, 371–380 CrossRef CAS PubMed.
- S.-Y. Chiu, C.-Y. Kao, C.-H. Chen, T.-C. Kuan, S.-C. Ong and C.-S. Lin, Bioresour. Technol., 2008, 99, 3389–3396 CrossRef CAS PubMed.
- J. N. Rosenberg, A. Mathias, K. Korth, M. J. Betenbaugh and G. A. Oyler, Biomass Bioenergy, 2011, 35, 3865–3876 CrossRef CAS.
- E. B. Sydney, W. Sturm, J. C. de Carvalho, V. Thomaz-Soccol, C. Larroche, A. Pandey and C. R. Soccol, Bioresour. Technol., 2010, 101, 5892–5896 CrossRef CAS PubMed.
- C. Yoo, S.-Y. Jun, J.-Y. Lee, C.-Y. Ahn and H.-M. Oh, Bioresour. Technol., 2010, 101, S71–S74 CrossRef CAS PubMed.
- B. Zhao, Y. Zhang, K. Xiong, Z. Zhang, X. Hao and T. Liu, Chem. Eng. Res. Des., 2011, 89, 1758–1762 CrossRef CAS.
- F.-F. Li, Z.-H. Yang, R. Zeng, G. Yang, X. Chang, J.-B. Yan and Y.-L. Hou, Ind. Eng. Chem. Res., 2011, 50, 6496–6502 CrossRef CAS.
- D. Tang, W. Han, P. Li, X. Miao and J. Zhong, Bioresour. Technol., 2011, 102, 3071–3076 CrossRef CAS PubMed.
- M. Hannon, J. Gimpel, M. Tran, B. Rasala and S. Mayfield, Biofuels, 2010, 1, 763–784 CrossRef CAS PubMed.
- H. Znad, G. Naderi, H. Ang and M. Tade, in Advances in Chemical Engineering, 2012, vol. 1, pp. 229–244 Search PubMed.
- E. Suali and R. Sarbatly, Renewable Sustainable Energy Rev., 2012, 16, 4316–4342 CrossRef CAS.
- A. P. Carvalho, L. A. Meireles and F. X. Malcata, Biotechnol. Prog., 2006, 22, 1490–1506 CrossRef CAS PubMed.
- O. Pulz, Appl. Microbiol. Biotechnol., 2001, 57, 287–293 CrossRef CAS PubMed.
- M. Janssen, J. Tramper, L. R. Mur and R. H. Wijffels, Biotechnol. Bioeng., 2003, 81, 193–210 CrossRef CAS PubMed.
- C. Ugwu, H. Aoyagi and H. Uchiyama, Bioresour. Technol., 2008, 99, 4021–4028 CrossRef CAS PubMed.
- F. Lehr and C. Posten, Curr. Opin. Biotechnol., 2009, 20, 280–285 CrossRef CAS PubMed.
- L. Batan, J. Quinn, B. Willson and T. Bradley, Environ. Sci. Technol., 2010, 44, 7975–7980 CrossRef CAS PubMed.
- B. Willson, The solix AGS system: a low-cost photobioreactor system for production of biofuels from microalgae, 2009 Search PubMed.
- F. K. Bentley and A. Melis, Biotechnol. Bioeng., 2012, 109, 100–109 CrossRef CAS PubMed.
- D. Briassoulis, P. Panagakis, M. Chionidis, D. Tzenos, A. Lalos, C. Tsinos, K. Berberidis and A. Jacobsen, Bioresour. Technol., 2010, 101, 6768–6777 CrossRef CAS PubMed.
- A. Henrard, M. De Morais and J. Costa, Bioresour. Technol., 2011, 102, 4897–4900 CrossRef CAS PubMed.
- R. Reyna-Velarde, E. Cristiani-Urbina, D. J. Hernández-Melchor, F. Thalasso and R. O. Cañizares-Villanueva, Chem. Eng. Process., 2010, 49, 97–103 CrossRef CAS.
- International Workshop on Algae Technology, in Bioforsk FOKUS, 2012, vol. 6, ch. Book of Abstracts Search PubMed.
- R. Radakovits, R. E. Jinkerson, A. Darzins and M. C. Posewitz, Eukaryotic Cell, 2010, 9, 486–501 CrossRef CAS PubMed.
- S. Amin, Energy Convers. Manage., 2009, 50, 1834–1840 CrossRef CAS.
- J.-S. Lee and J.-P. Lee, Biotechnol. Bioprocess Eng., 2003, 8, 354–359 CrossRef CAS.
- X. Zeng, M. K. Danquah, X. D. Chen and Y. Lu, Renewable Sustainable Energy Rev., 2011, 15, 3252–3260 CrossRef CAS.
- H. Greenwell, L. Laurens, R. Shields, R. Lovitt and K. Flynn, J. R. Soc., Interface, 2010, 7, 703–726 CrossRef CAS PubMed.
- N. M. D. Courchesne, A. Parisien, B. Wang and C. Q. Lan, J. Biotechnol., 2009, 141, 31–41 CrossRef CAS PubMed.
- G. Potvin and Z. Zhang, Biotechnol. Adv., 2010, 28, 910–918 CrossRef CAS PubMed.
- M. Y. Menetrez, Environ. Sci. Technol., 2012, 46, 7073–7085 CrossRef CAS PubMed.
- C. S. Jones and S. P. Mayfield, Curr. Opin. Biotechnol., 2012, 23, 346–351 CrossRef CAS PubMed.
- F. X. Malcata, Trends Biotechnol., 2011, 29, 542–549 CrossRef CAS PubMed.
- R. Luque, Energy Environ. Sci., 2010, 3, 254–257 CAS.
- D. J. Gilmour and W. B. Zimmerman, Biofuels, 2012, 3, 511–513 CrossRef CAS.
- M. Hildebrand, R. M. Abbriano, J. E. Polle, J. C. Traller, E. M. Trentacoste, S. R. Smith and A. K. Davis, Curr. Opin. Chem. Biol., 2013, 17, 506–514 CrossRef CAS PubMed.
- J. N. Rosenberg, G. A. Oyler, L. Wilkinson and M. J. Betenbaugh, Curr. Opin. Biotechnol., 2008, 19, 430–436 CrossRef CAS PubMed.
- R. H. Wijffels and M. J. Barbosa, Science, 2010, 329, 796–799 CrossRef CAS PubMed.
- M. C. Posewitz, A. Dubini, J. E. Meuser, M. Seibert and M. L. Ghirardi, The Chlamydomonas Sourcebook: Organellar and Metabolic Processes, 2009, vol. 2, pp. 217–246 Search PubMed.
- L. Rodolfi, G. Chini Zittelli, N. Bassi, G. Padovani, N. Biondi, G. Bonini and M. R. Tredici, Biotechnol. Bioeng., 2009, 102, 100–112 CrossRef CAS PubMed.
- A. Converti, A. A. Casazza, E. Y. Ortiz, P. Perego and M. Del Borghi, Chem. Eng. Process., 2009, 48, 1146–1151 CrossRef CAS.
- P. G. Stephenson, C. M. Moore, M. J. Terry, M. V. Zubkov and T. S. Bibby, Trends Biotechnol., 2011, 29, 615–623 CrossRef CAS PubMed.
- R. Huerlimann, R. De Nys and K. Heimann, Biotechnol. Bioeng., 2010, 107, 245–257 CrossRef CAS PubMed.
- D. Luo, Z. Hu, D. G. Choi, V. M. Thomas, M. J. Realff and R. R. Chance, Environ. Sci. Technol., 2010, 44, 8670–8677 CrossRef CAS PubMed.
- A. Singh and S. I. Olsen, Appl. Energy, 2011, 88, 3548–3555 CrossRef CAS.
- R. Slade and A. Bauen, Biomass Bioenergy, 2013, 53, 39–38 CrossRef.
- E. Molina Grima, E.-H. Belarbi, F. Acién Fernández, A. Robles Medina and Y. Chisti, Biotechnol. Adv., 2003, 20, 491–515 CrossRef CAS PubMed.
- N. Uduman, Y. Qi, M. K. Danquah, G. M. Forde and A. Hoadley, J. Renewable Sustainable Energy, 2010, 2, 012701 CrossRef.
- L. A. Schlesinger and J. L. Heskett, Sloan Management Review, 1991, 32, 17–29 Search PubMed.
- D. Vandamme, I. Foubert and K. Muylaert, Trends Biotechnol., 2013, 31, 233–239 CrossRef CAS PubMed.
- M. Packer, Energy Policy, 2009, 37, 3428–3437 CrossRef.
- S.-H. Ho, C.-Y. Chen, D.-J. Lee and J.-S. Chang, Biotechnol. Adv., 2011, 29, 189–198 CrossRef CAS PubMed.
- L. B. Brentner, M. J. Eckelman and J. B. Zimmerman, Environ. Sci. Technol., 2011, 45, 7060–7067 CrossRef CAS PubMed.
- P. T. Pienkos and A. Darzins, Biofuels, Bioprod. Biorefin., 2009, 3, 431–440 CrossRef CAS.
- A. Wileman, A. Ozkan and H. Berberoglu, Bioresour. Technol., 2012, 104, 432–439 CrossRef CAS PubMed.
- D. Vandamme, I. Foubert and K. Muylaert, Trends Biotechnol., 2013, 31, 233–239 CrossRef CAS PubMed.
- A. Ben-Amotz and M. Avron, Trends Biotechnol., 1990, 8, 121–126 CrossRef CAS.
- V. M. Rwehumbiza, R. Harrison and L. Thomsen, Chem. Eng. J., 2012, 200, 168–175 CrossRef.
- R. H. Wijffels, M. J. Barbosa and M. H. Eppink, Biofuels, Bioprod. Biorefin., 2010, 4, 287–295 CrossRef CAS.
- N. B. Wyatt, L. M. Gloe, P. V. Brady, J. C. Hewson, A. M. Grillet, M. G. Hankins and P. I. Pohl, Biotechnol. Bioeng., 2012, 109, 493–501 CrossRef CAS PubMed.
- X. Zhang, P. Amendola, J. C. Hewson, M. Sommerfeld and Q. Hu, Bioresour. Technol., 2012, 116, 477–484 CrossRef CAS PubMed.
- J. Bratby, Coagulation and flocculation in water and wastewater treatment, IWA publishing, 2006 Search PubMed.
- Y.-R. Chang and D.-J. Lee, Drying Technol., 2012, 30, 1317–1322 CrossRef CAS.
- D. Vandamme, I. Foubert, B. Meesschaert and K. Muylaert, J. Appl. Phycol., 2010, 22, 525–530 CrossRef.
- H. Zheng, Z. Gao, J. Yin, X. Tang, X. Ji and H. Huang, Bioresour. Technol., 2012, 112, 212–220 CrossRef CAS PubMed.
- C. M. L. L. Teixeira, F. V. Kirsten and P. C. N. Teixeira, J. Appl. Phycol., 2012, 24, 557–563 CrossRef CAS.
- N. Rashid, S. U. Rehman and J.-I. Han, Process Biochem., 2013, 48, 1107–1110 CrossRef CAS.
- N. Rashid, M. S. U. Rehman and J.-I. Han, Chem. Eng. J., 2013, 226, 238–242 CrossRef CAS.
- J. Morales, J. De La Noüe and G. Picard, Aquacultural Engineering, 1985, 4, 257–270 CrossRef.
- A. Ahmad, N. M. Yasin, C. Derek and J. Lim, Chem. Eng. J., 2011, 173, 879–882 CrossRef CAS.
- M. S. Farid, A. Shariati, A. Badakhshan and B. Anvaripoor, Bioresour. Technol., 2013, 131, 555–559 CrossRef CAS PubMed.
- W. Farooq, Y.-C. Lee, J.-I. Han, C. H. Darpito, M. Choi and J.-W. Yang, Green Chem., 2013, 15, 749–755 RSC.
- Y.-C. Lee, B. Kim, W. Farooq, J. Chung, J.-I. Han, H.-J. Shin, S. H. Jeong, J.-Y. Park, J.-S. Lee and Y.-K. Oh, Bioresour. Technol., 2013, 132, 440–445 CrossRef CAS PubMed.
- Y.-C. Lee, Y. S. Huh, W. Farooq, J. Chung, J.-I. Han, H.-J. Shin, S. H. Jeong, J.-S. Lee, Y.-K. Oh and J.-Y. Park, Bioresour. Technol., 2013, 137, 74–81 CrossRef CAS PubMed.
- H.-K. Han, Y.-C. Lee, M.-Y. Lee, A. J. Patil and H.-J. Shin, ACS Appl. Mater. Interfaces, 2011, 3, 2564–2572 CAS.
- K. Spilling, J. Seppälä and T. Tamminen, J. Appl. Phycol., 2011, 23, 959–966 CrossRef CAS.
- L. Christenson and R. Sims, Biotechnol. Adv., 2011, 29, 686–702 CrossRef CAS PubMed.
- A. Schlesinger, D. Eisenstadt, A. Bar-Gil, H. Carmely, S. Einbinder and J. Gressel, Biotechnol. Adv., 2012, 30, 1023–1030 CrossRef CAS PubMed.
- T. J. Lundquist, I. C. Woertz, N. Quinn and J. R. Benemann, A Realistic Technology and Engineering Assessment of Algae Biofuel Production, Energy Biosciences Institute, 2010, pp. 1–178 Search PubMed.
- D. Vandamme, I. Foubert, I. Fraeye, B. Meesschaert and K. Muylaert, Bioresour. Technol., 2012, 105, 114–119 CrossRef CAS PubMed.
- Z. Wu, Y. Zhu, W. Huang, C. Zhang, T. Li, Y. Zhang and A. Li, Bioresour. Technol., 2012, 110, 496–502 CrossRef CAS PubMed.
- K.-Y. Show, D.-J. Lee and J.-S. Chang, Bioresour. Technol., 2011, 102, 8524–8533 CrossRef CAS PubMed.
- R. Bosma, W. A. van Spronsen, J. Tramper and R. H. Wijffels, J. Appl. Phycol., 2003, 15, 143–153 CrossRef.
- D. Vandamme, S. C. V. Pontes, K. Goiris, I. Foubert, L. J. J. Pinoy and K. Muylaert, Biotechnol. Bioeng., 2011, 108, 2320–2329 CrossRef CAS PubMed.
- J. Kim, B.-G. Ryu, B.-K. Kim, J.-I. Han and J.-W. Yang, Bioresour. Technol., 2012, 111, 268–275 CrossRef CAS PubMed.
- L. Gouveia, Microalgae as a Feedstock for Biofuels, Springer, 2011 Search PubMed.
- M. Cerff, M. Morweiser, R. Dillschneider, A. Michel, K. Menzel and C. Posten, Bioresour. Technol., 2012, 118, 289–295 CrossRef CAS PubMed.
- Y.-R. Hu, F. Wang, S.-K. Wang, C.-Z. Liu and C. Guo, Bioresour. Technol., 2013, 138, 387–390 CrossRef CAS PubMed.
- L. Xu, C. Guo, F. Wang, S. Zheng and C.-Z. Liu, Bioresour. Technol., 2011, 102, 10047–10051 CrossRef CAS PubMed.
- J. K. Lim, D. C. J. Chieh, S. A. Jalak, P. Y. Toh, N. H. M. Yasin, B. W. Ng and A. L. Ahmad, Small, 2012, 8, 1683–1692 CrossRef CAS PubMed.
- D. Liu, F. Li and B. Zhang, Water Sci. Technol., 2009, 59, 1085–1091 CrossRef CAS PubMed.
- E. Bejor, C. Mota, N. Ogarekpe, K. Emerson and J. Ukpata, Int. J. Sustain. Dev., 2013, 2, 1–11 Search PubMed.
- A. W. Larkum, I. L. Ross, O. Kruse and B. Hankamer, Trends Biotechnol., 2012, 30, 198–205 CrossRef CAS PubMed.
- R. Craggs, D. Sutherland and H. Campbell, J. Appl. Phycol., 2012, 24, 329–337 CrossRef CAS.
- P. M. Schenk, S. R. Thomas-Hall, E. Stephens, U. C. Marx, J. H. Mussgnug, C. Posten, O. Kruse and B. Hankamer, BioEnergy Res., 2008, 1, 20–43 CrossRef.
- R. L. Taylor, J. D. Rand and G. S. Caldwell, Mar. Biotechnol., 2012, 14, 774–781 CrossRef CAS PubMed.
- R. Eldridge, D. Hill and B. Gladman, J. Appl. Phycol., 2012, 24, 1667–1679 CrossRef CAS.
- S. Salim, M. Vermuë and R. Wijffels, Bioresour. Technol., 2012, 118, 49–55 CrossRef CAS PubMed.
- W. Zhou, Y. Cheng, Y. Li, Y. Wan, Y. Liu, X. Lin and R. Ruan, Appl. Biochem. Biotechnol., 2012, 167, 214–228 CrossRef CAS PubMed.
- J. Zhang and B. Hu, Bioresour. Technol., 2012, 114, 529–535 CrossRef CAS PubMed.
- G. Gutzeit, D. Lorch, A. Weber, M. Engels and U. Neis, Water Sci. Technol., 2005, 52, 9–18 CAS.
- A. K. Lee, D. M. Lewis and P. J. Ashman, J. Appl. Phycol., 2009, 21, 559–567 CrossRef CAS.
- S. Van Den Hende, H. Vervaeren, H. Saveyn, G. Maes and N. Boon, Biotechnol. Bioeng., 2011, 108, 549–558 CrossRef CAS PubMed.
- Y. Su, A. Mennerich and B. Urban, Water Res., 2011, 45, 3351–3358 CrossRef CAS PubMed.
- Y. Li, A. Mangott, S. Grierson and P. M. Schenk, Biofuels, 2013, 4, 263–266 CrossRef CAS.
- W. Zhou, M. Min, B. Hu, X. Ma, Y. Liu, Q. Wang, J. Shi, P. Chen and R. Ruan, Sep. Purif. Technol., 2013, 107, 158–165 CrossRef CAS.
- J. Lee, D.-H. Cho, R. Ramanan, B.-H. Kim, H.-M. Oh and H.-S. Kim, Bioresour. Technol., 2013, 131, 195–201 CrossRef CAS PubMed.
- Z. Gu, Z. Liao, M. Schulz, J. R. Davis, J. C. Baygents and J. Farrell, Ind. Eng. Chem. Res., 2009, 48, 3112–3117 CrossRef CAS.
- A. K. Lee, D. M. Lewis and P. J. Ashman, Appl. Energy, 2013, 108, 45–53 CrossRef.
- S. Babel and S. Takizawa, Desalination, 2010, 261, 46–51 CrossRef CAS.
- X. Zhang, Q. Hu, M. Sommerfeld, E. Puruhito and Y. Chen, Bioresour. Technol., 2010, 101, 5297–5304 CrossRef CAS PubMed.
- M. Hung and J. Liu, Colloids Surf., B, 2006, 51, 157–164 CrossRef CAS PubMed.
- S. Zou, Y. Gu, D. Xiao and C. Y. Tang, J. Membr. Sci., 2011, 366, 356–362 CrossRef CAS.
- C. Chow, S. Panglisch, J. House, M. Drikas, M. Burch and R. Gimbel, Aqua, 1997, 46, 324–334 CAS.
- N. Rossignol, L. Vandanjon, P. Jaouen and F. Quemeneur, Aquacultural Engineering, 1999, 20, 191–208 CrossRef.
- A. Wei, G. Zeng, G. Huang, J. Liang and X. Li, Int. J. Environ. Sci. Technol., 2009, 6, 395–406 CrossRef CAS.
- C.-H. Zhang, F.-l. Yang, W.-J. Wang and B. Chen, Sep. Purif. Technol., 2008, 61, 276–286 CrossRef CAS.
- L. Heng, Y. Yanling, G. Weijia, L. Xing and L. Guibai, Desalination, 2008, 222, 74–80 CrossRef CAS.
- J.-B. Castaing, A. Massé, M. Pontié, V. Séchet, J. Haure and P. Jaouen, Desalination, 2010, 253, 71–77 CrossRef CAS.
- S. Babel and S. Takizawa, Water Sci. Technol., 2000, 41, 327–335 Search PubMed.
- N. Rossi, I. Petit, P. Jaouen, P. Legentilhomme and M. Derouiniot, Sep. Sci. Technol., 2005, 40, 3033–3050 CrossRef CAS.
- W. Liu, Harvesting of Microalgae by Membrane Filtration, Amsterdam, 2011 Search PubMed.
- X. Sun, C. Wang, Y. Tong, W. Wang and J. Wei, Algal Res., 2013, 2, 437–444 CrossRef.
- T. Hwang, S.-J. Park, Y.-K. Oh, N. Rashid and J.-I. Han, Bioresour. Technol., 2013, 139, 379–382 CrossRef CAS PubMed.
- M. Y. Jaffrin, J. Membr. Sci., 2008, 324, 7–25 CrossRef CAS.
- M. R. Bilad, G. Mezohegyi, P. Declerck and I. F. Vankelecom, Water Res., 2012, 46, 63–72 CrossRef CAS PubMed.
- M. Frappart, A. Massé, M. Y. Jaffrin, J. Pruvost and P. Jaouen, Desalination, 2011, 265, 279–283 CrossRef CAS.
- S. D. Rios, E. Clavero, J. Salvadó, X. Farriol and C. Torras, Ind. Eng. Chem. Res., 2010, 50, 2455–2460 CrossRef.
- D. A. Ladner, D. R. Vardon and M. M. Clark, J. Membr. Sci., 2010, 356, 33–43 CrossRef CAS.
- M. R. Bilad, V. Discart, D. Vandamme, I. Foubert, K. Muylaert and I. F. Vankelecom, Bioresour. Technol., 2013, 138, 329–338 CrossRef CAS PubMed.
- M. Kröger and F. Müller-Langer, Biofuels, 2012, 3, 333–349 CrossRef.
- A. Ross, J. Jones, M. Kubacki and T. Bridgeman, Bioresour. Technol., 2008, 99, 6494–6504 CrossRef CAS PubMed.
- L. Xu, D. W. Brilman, J. A. Withag, G. Brem and S. Kersten, Bioresour. Technol., 2011, 102, 5113–5122 CrossRef CAS PubMed.
- D. A. Laird, P. Fleming, D. D. Davis, R. Horton, B. Wang and D. L. Karlen, Geoderma, 2010, 158, 443–449 CrossRef CAS.
- R. B. Levine, T. Pinnarat and P. E. Savage, Energy Fuels, 2010, 24, 5235–5243 CrossRef CAS.
- P. D. Patil, V. G. Gude, A. Mannarswamy, S. Deng, P. Cooke, S. Munson-McGee, I. Rhodes, P. Lammers and N. Nirmalakhandan, Bioresour. Technol., 2011, 102, 118–122 CrossRef CAS PubMed.
- T.-O. Matsui, A. Nishihara, C. Ueda, M. Ohtsuki, N.-O. Ikenaga and T. Suzuki, Fuel, 1997, 76, 1043–1048 CrossRef CAS.
- N.-O. Ikenaga, C. Ueda, T. Matsui, M. Ohtsuki and T. Suzuki, Energy Fuels, 2001, 15, 350–355 CrossRef CAS.
- A. A. Peterson, F. Vogel, R. P. Lachance, M. Fröling, M. J. Antal, Jr and J. W. Tester, Energy Environ. Sci., 2008, 1, 32–65 CAS.
- R. H. Perry, D. W. Green and J. O. Maloney, Perry's chemical engineers' handbook, McGraw-Hill New York, 2008 Search PubMed.
- L. Zhang, C. C. Xu and P. Champagne, Energy Convers. Manage., 2010, 51, 969–982 CrossRef CAS.
- L. Garcia Alba, C. Torri, C. Samorì, J. van der Spek, D. Fabbri, S. R. Kersten and D. W. Brilman, Energy Fuels, 2011, 26, 642–657 CrossRef.
- A. G. Chakinala, D. W. Brilman, W. P. van Swaaij and S. R. Kersten, Ind. Eng. Chem. Res., 2009, 49, 1113–1122 CrossRef.
- S. Stucki, F. Vogel, C. Ludwig, A. G. Haiduc and M. Brandenberger, Energy Environ. Sci., 2009, 2, 535–541 CAS.
- A. Kruse and E. Dinjus, J. Supercrit. Fluids, 2007, 39, 362–380 CrossRef CAS.
- W. Wagner and A. Pruss, J. Phys. Chem. Ref. Data, 2002, 31, 387–536 CrossRef CAS.
- M. Uematsu and E. Franck, Static dielectric constant of water and steam, American Chemical Society and the American Institute of Physics for the National Bureau of Standards, 1980 Search PubMed.
- A. V. Bandura and S. N. Lvov, J. Phys. Chem. Ref. Data, 2006, 35, 15–30 CrossRef CAS.
- A. Kruse and E. Dinjus, J. Supercrit. Fluids, 2007, 41, 361–379 CrossRef CAS.
- U.S. DOE 2010, National Algal Biofuels Technology Roadmap, U.S. Department of Energy, Office of Energy Efficiency and Renewable Energy, Biomass Program, http://www.biomass.energy.gov Search PubMed.
- P. Biller and A. B. Ross, Biofuels, 2012, 3, 603–623 CrossRef CAS.
- P. E. Savage, R. B. Levine and C. M. Huelsman, in Hydrothermal processing of biomass, ed. M. Crocker, Royal Soc. Chem. Pub., 2010, ch. 8, pp. 190–219 Search PubMed.
- N. Akiya and P. E. Savage, Chem. Rev., 2002, 102, 2725–2750 CrossRef CAS PubMed.
- P. E. Savage, Science, 2012, 338, 1039–1040 CrossRef CAS PubMed.
- L. Florin, A. Tsokoglou and T. Happe, J. Biol. Chem., 2001, 276, 6125–6132 CrossRef CAS PubMed.
- A. Melis and T. Happe, Plant Physiol., 2001, 127, 740–748 CrossRef CAS PubMed.
- M. L. Ghirardi, M. C. Posewitz, P.-C. Maness, A. Dubini, J. Yu and M. Seibert, Annu. Rev. Plant Biol., 2007, 58, 71–91 CrossRef CAS PubMed.
- C. E. Lubner, A. M. Applegate, P. Knörzer, A. Ganago, D. A. Bryant, T. Happe and J. H. Golbeck, Proc. Natl. Acad. Sci. U. S. A., 2011, 108, 20988–20991 CrossRef CAS PubMed.
- H. Gaffron, Nature, 1939, 143, 204–205 CrossRef CAS.
- H. Gaffron and J. Rubin, J. Gen. Physiol., 1942, 26, 219–240 CrossRef CAS PubMed.
- S. I. Allakhverdiev, V. D. Kreslavski, V. Thavasi, S. K. Zharmukhamedov, V. V. Klimov, T. Nagata, H. Nishihara and S. Ramakrishna, Photochem. Photobiol. Sci., 2009, 8, 148–156 CAS.
- S. I. Allakhverdiev, V. Thavasi, V. D. Kreslavski, S. K. Zharmukhamedov, V. V. Klimov, S. Ramakrishna, D. A. Los, M. Mimuro, H. Nishihara and R. Carpentier, J. Photochem. Photobiol., C, 2010, 11, 101–113 CrossRef CAS.
- S. T. Stripp and T. Happe, Dalton Trans., 2009, 9960–9969 RSC.
- J. E. Polle, J. R. Benemann, A. Tanaka and A. Melis, Planta, 2000, 211, 335–344 CrossRef CAS PubMed.
- J. E. Polle, S.-D. Kanakagiri and A. Melis, Planta, 2003, 217, 49–59 CAS.
- M. L. Ghirardi, R. K. Togasaki and M. Seibert, Appl. Biochem. Biotechnol., 1997, 63, 141–151 CrossRef PubMed.
- V. A. Boichenko, E. Greenbaum and M. Seibert, Photoconversion of solar energy, molecular to global photosynthesis, 2004, vol. 2, pp. 397–452 Search PubMed.
- A. Melis, L. Zhang, M. Forestier, M. L. Ghirardi and M. Seibert, Plant Physiol., 2000, 122, 127–136 CrossRef CAS PubMed.
- M. L. Ghirardi, L. Zhang, J. W. Lee, T. Flynn, M. Seibert, E. Greenbaum and A. Melis, Trends Biotechnol., 2000, 18, 506–511 CrossRef CAS PubMed.
- L. Zhang, T. Happe and A. Melis, Planta, 2002, 214, 552–561 CrossRef CAS PubMed.
- A. Melis, Planta, 2007, 226, 1075–1086 CrossRef CAS PubMed.
- A. Hemschemeier, A. Melis and T. Happe, Photosynth. Res., 2009, 102, 523–540 CrossRef CAS PubMed.
- O. Kruse and B. Hankamer, Curr. Opin. Biotechnol., 2010, 21, 238–243 CrossRef CAS PubMed.
- O. Kruse, J. Rupprecht, K.-P. Bader, S. Thomas-Hall, P. M. Schenk, G. Finazzi and B. Hankamer, J. Biol. Chem., 2005, 280, 34170–34177 CrossRef CAS PubMed.
- U. Bossel, B. Eliasson and G. Taylor, Cogeneration and Distributed Generation Journal, 2003, 18, 29–70 CrossRef.
- C. Baker and R. Shaner, Int. J. Hydrogen Energy, 1978, 3, 321–334 CrossRef CAS.
- M. Syed, S. Sherif, T. Veziroglu and J. W. Sheffield, Int. J. Hydrogen Energy, 1998, 23, 565–576 CrossRef CAS.
- M. Temporal, J. L. Cela, A. Piriz, N. Grandjouan, N. Tahir and D. Hoffmann, Laser Part. Beams, 2005, 23, 137–142 CrossRef CAS.
- Z. Dehouche, M. Savard, F. Laurencelle and J. Goyette, J. Alloys Compd., 2005, 400, 276–280 CrossRef CAS.
- J.-K. Kim, I.-S. Park, K. J. Kim and K. Gawlik, Int. J. Hydrogen Energy, 2008, 33, 870–877 CrossRef CAS.
- A. Zaluska, L. Zaluski and J. Ström-Olsen, Appl. Phys. A: Mater. Sci. Process., 2001, 72, 157–165 CrossRef CAS.
- B. Sakintuna, F. Lamari-Darkrim and M. Hirscher, Int. J. Hydrogen Energy, 2007, 32, 1121–1140 CrossRef CAS.
- L. J. Murray, M. Dincă and J. R. Long, Chem. Soc. Rev., 2009, 38, 1294–1314 RSC.
- D. Slattery and M. Hampton, Complex hydrides for hydrogen storage, Department of Energy, Office of Energy Efficiency and Renewable Energy, United States, 2003 Search PubMed.
- N. L. Rosi, J. Eckert, M. Eddaoudi, D. T. Vodak, J. Kim, M. O'Keeffe and O. M. Yaghi, Science, 2003, 300, 1127–1129 CrossRef CAS PubMed.
- H.-W. Li, Y. Yan, S.-i. Orimo, A. Züttel and C. M. Jensen, Energies, 2011, 4, 185–214 CrossRef CAS.
- M. E. Bluhm, M. G. Bradley, R. Butterick, U. Kusari and L. G. Sneddon, J. Am. Chem. Soc., 2006, 128, 7748–7749 CrossRef CAS PubMed.
- T. A. Abtew, B.-c. Shih, P. Dev, V. H. Crespi and P. Zhang, Phys. Rev. B: Condens. Matter Mater. Phys., 2011, 83, 094108 CrossRef.
- M. Felderhoff, C. Weidenthaler, R. von Helmolt and U. Eberle, Phys. Chem. Chem. Phys., 2007, 9, 2643–2653 RSC.
- Z. Li, T. Sun and J. Jia, Fuel Process. Technol., 2010, 91, 1162–1167 CrossRef CAS.
- Y. Shen, T. Sun and J. Jia, Energy Fuels, 2011, 25, 2963–2967 CrossRef CAS.
- Y. Shen, T. Sun and J. Jia, Fuel, 2012, 96, 250–256 CrossRef CAS.
- Y. Shen, X. Liu, T. Sun and J. Jia, RSC Adv., 2012, 2, 8867–8882 RSC.
- H.-S. Kim, H. Lee, K.-S. Han, J.-H. Kim, M.-S. Song, M.-S. Park, J.-Y. Lee and J.-K. Kang, J. Phys. Chem. B, 2005, 109, 8983–8986 CrossRef CAS PubMed.
- A. Reddy and S. Ramaprabhu, Int. J. Hydrogen Energy, 2007, 32, 3998–4004 CrossRef CAS.
- T. Happe, A. Hemschemeier, M. Winkler and A. Kaminski, Trends Plant Sci., 2002, 7, 246–250 CrossRef CAS PubMed.
- M. L. Ghirardi, A. Dubini, J. Yu and P.-C. Maness, Chem. Soc. Rev., 2009, 38, 52–61 RSC.
- M. Seibert, P. W. King, M. C. Posewitz, A. Melis and M. L. Ghirardi, Bioenergy, ASM Press, Washington, DC, 2008, pp. 273–291 Search PubMed.
- C. Tard and C. J. Pickett, Chem. Rev., 2009, 109, 2245–2274 CrossRef CAS PubMed.
- J. A. Cracknell, K. A. Vincent and F. A. Armstrong, Chem. Rev., 2008, 108, 2439–2461 CrossRef CAS PubMed.
- M. Hambourger, M. Gervaldo, D. Svedruzic, P. W. King, D. Gust, M. Ghirardi, A. L. Moore and T. A. Moore, J. Am. Chem. Soc., 2008, 130, 2015–2022 CrossRef CAS PubMed.
- S. Krishnan and F. A. Armstrong, Chem. Sci., 2012, 3, 1015–1023 RSC.
- A. Ciaccafava, A. De Poulpiquet, V. Techer, M. Giudici-Orticoni, S. Tingry, C. Innocent and E. Lojou, Electrochem. Commun., 2012, 23, 25–28 CrossRef CAS.
- J. Peters, Science, 1999, 283, 2102 CAS.
- Y. Nicolet, C. Piras, P. Legrand, C. E. Hatchikian and J. C. Fontecilla-Camps, Structure, 1999, 7, 13–23 CrossRef CAS PubMed.
- D. W. Mulder, E. S. Boyd, R. Sarma, R. K. Lange, J. A. Endrizzi, J. B. Broderick and J. W. Peters, Nature, 2010, 465, 248–251 CrossRef CAS PubMed.
- G. Berggren, A. Adamska, C. Lambertz, T. Simmons, J. Esselborn, M. Atta, S. Gambarelli, J.-M. Mouesca, E. Reijerse and W. Lubitz, Nature, 2013, 499, 66–69 CrossRef CAS PubMed.
- Y. Nicolet, A. L. de Lacey, X. Vernede, V. M. Fernandez, E. C. Hatchikian and J. C. Fontecilla-Camps, J. Am. Chem. Soc., 2001, 123, 1596–1601 CrossRef CAS PubMed.
- A. Silakov, B. Wenk, E. Reijerse and W. Lubitz, Phys. Chem. Chem. Phys., 2009, 11, 6592–6599 RSC.
- R. Mertens and A. Liese, Curr. Opin. Biotechnol., 2004, 15, 343–348 CrossRef CAS PubMed.
- S. Shestakov and L. Mikheeva, Russ. J. Genet., 2006, 42, 1272–1284 CrossRef CAS.
- J. Appel and R. Schulz, J. Photochem. Photobiol., B, 1998, 47, 1–11 CrossRef CAS.
- J. B. McKinlay and C. S. Harwood, Curr. Opin. Biotechnol., 2010, 21, 244–251 CrossRef CAS PubMed.
- K. Srirangan, M. E. Pyne and C. Perry Chou, Bioresour. Technol., 2011, 102, 8589–8604 CrossRef CAS PubMed.
- R. Harun, M. Singh, G. M. Forde and M. K. Danquah, Renewable Sustainable Energy Rev., 2010, 14, 1037–1047 CrossRef CAS.
- A. Demirbas, Energy Convers. Manage., 2009, 50, 14–34 CrossRef CAS.
- K.-Y. Show, D.-J. Lee and J.-S. Chang, Bioresour. Technol., 2011, 102, 8524–8533 CrossRef CAS PubMed.
- K. Nath and D. Das, Appl. Microbiol. Biotechnol., 2004, 65, 520–529 CrossRef CAS PubMed.
- A. Melis and M. R. Melnicki, Int. J. Hydrogen Energy, 2006, 31, 1563–1573 CrossRef CAS.
- Y. Guan, M. Deng, X. Yu and W. Zhang, Biochem. Eng. J., 2004, 19, 69–73 CrossRef CAS.
- N. Rashid, M. S. U. Rehman, S. Memon, Z. Ur Rahman, K. Lee and J.-I. Han, Renewable Sustainable Energy Rev., 2013, 22, 571–579 CrossRef CAS.
- B. D. James, G. N. Baum, J. Perez and K. N. Baum, Technoeconomic Boundary Analysis of Biological Pathways to Hydrogen Production, US Department of Energy, September, 2009, http://www.nrel.gov/docs/fy09osti/46674.pdf Search PubMed.
- O. Jorquera, A. Kiperstok, E. A. Sales, M. Embiruçu and M. L. Ghirardi, Bioresour. Technol., 2010, 101, 1406–1413 CrossRef CAS PubMed.
- F. Romagnoli, D. Blumberga and I. Pilicka, Int. J. Hydrogen Energy, 2011, 36, 7866–7871 CrossRef CAS.
- W. A. Amos, Updated cost analysis of photobiological hydrogen production from Chlamydomonas reinhardtii green algae, Milestone Completion Report, National Renewable Energy Laboratories, Golden, CO, NREL/MP-560-35593, January 2004 Search PubMed.
- International Energy Agency, 2009.
- M. Timmins, S. R. Thomas-Hall, A. Darling, E. Zhang, B. Hankamer, U. C. Marx and P. M. Schenk, J. Exp. Bot., 2009, 60, 1691–1702 CrossRef CAS PubMed.
- F. Mus, A. Dubini, M. Seibert, M. C. Posewitz and A. R. Grossman, J. Biol. Chem., 2007, 282, 25475–25486 CrossRef CAS PubMed.
- A. F. Ferreira, J. Ortigueira, L. Alves, L. Gouveia, P. Moura and C. Silva, Biomass Bioenergy, 2013, 49, 249–259 CrossRef CAS.
- A. V. Nguyen, S. R. Thomas-Hall, A. Malnoë, M. Timmins, J. H. Mussgnug, J. Rupprecht, O. Kruse, B. Hankamer and P. M. Schenk, Eukaryotic Cell, 2008, 7, 1965–1979 CrossRef CAS PubMed.
- L. Zhang and A. Melis, Philos. Trans. R. Soc. London, Ser. B, 2002, 357, 1499–1509 CrossRef CAS PubMed.
- J. Rupprecht, J. Biotechnol., 2009, 142, 10–20 CrossRef CAS PubMed.
- O. Jorquera, A. Kiperstok, E. A. Sales, M. Embiruçu and M. L. Ghirardi, Int. J. Hydrogen Energy, 2008, 33, 2167–2177 CrossRef CAS.
- N. Rolland, A. Atteia, P. Decottignies, J. Garin, M. Hippler, G. Kreimer, S. D. Lemaire, M. Mittag and V. Wagner, Curr. Opin. Microbiol., 2009, 12, 285–291 CrossRef CAS PubMed.
- M. C. Posewitz, S. L. Smolinski, S. Kanakagiri, A. Melis, M. Seibert and M. L. Ghirardi, Plant Cell, 2004, 16, 2151–2163 CrossRef CAS PubMed.
- M. Posewitz, P. King, S. Smolinski, R. D. Smith, A. Ginley, M. Ghirardi and M. Seibert, Biochem. Soc. Trans., 2005, 33, 102–104 CrossRef CAS PubMed.
- J. D. Bloom, M. M. Meyer, P. Meinhold, C. R. Otey, D. MacMillan and F. H. Arnold, Curr. Opin. Struct. Biol., 2005, 15, 447–452 CrossRef CAS PubMed.
- O. Lenz, A. Gleiche, A. Strack and B. Friedrich, J. Bacteriol., 2005, 187, 6590–6595 CrossRef CAS PubMed.
- M. L. Ghirardi, P. W. King, M. C. Posewitz, P. C. Maness, A. Fedorov, K. Kim, J. Cohen, K. Schulten and M. Seibert, Biochem. Soc. Trans., 2005, 33, 70–72 CrossRef CAS PubMed.
- B. Esper, A. Badura and M. Rögner, Trends Plant Sci., 2006, 11, 543–549 CrossRef CAS PubMed.
- S. Manish and R. Banerjee, Int. J. Hydrogen Energy, 2008, 33, 279–286 CrossRef CAS.
- B. E. Logan and J. M. Regan, Trends Microbiol., 2006, 14, 512–518 CrossRef CAS PubMed.
- J. K. Pittman, A. P. Dean and O. Osundeko, Bioresour. Technol., 2011, 102, 17–25 CrossRef CAS PubMed.
- H. Muhammad, K. Z. Muhammad, S. K. Muhammad and A. Javaid, Journal of the Pakistan Institute of Chemical Engineers, 2012, 40, 83–86 Search PubMed.
- A. J. Usman, T. J. Muhammad and R. C. Imran, Journal of the Pakistan Institute of Chemical Engineers, 2011, 39, 23–27 Search PubMed.
- C.-H. Liu, C.-Y. Chang, Q. Liao, X. Zhu, C.-F. Liao and J.-S. Chang, Int. J. Hydrogen Energy, 2013, 38, 15807–15814 CrossRef CAS.
- K. Kumar, S. Roy and D. Das, Bioresour. Technol., 2013, 145, 116–122 CrossRef CAS PubMed.
- Y.-M. Yun, K.-W. Jung, D.-H. Kim, Y.-K. Oh, S.-K. Cho and H.-S. Shin, Bioresour. Technol., 2013, 141, 220–226 CrossRef CAS PubMed.
- J.-H. Park, H.-C. Cheon, J.-J. Yoon, H.-D. Park and S.-H. Kim, Int. J. Hydrogen Energy, 2013, 38, 6130–6136 CrossRef CAS.
- A. Xia, J. Cheng, R. Lin, H. Lu, J. Zhou and K. Cen, Bioresour. Technol., 2013, 138, 204–213 CrossRef CAS PubMed.
- A. Xia, J. Cheng, L. Ding, R. Lin, R. Huang, J. Zhou and K. Cen, Bioresour. Technol., 2013, 146, 436–443 CrossRef CAS PubMed.
- A. F. Ferreira, J. Ortigueira, L. Alves, L. Gouveia, P. Moura and C. M. Silva, Biomass Bioenergy, 2013, 49, 249–259 CrossRef CAS.
- J. Mathews and G. Wang, Int. J. Hydrogen Energy, 2009, 34, 7404–7416 CrossRef CAS.
- S. Kosourov, M. Seibert and M. L. Ghirardi, Plant Cell Physiol., 2003, 44, 146–155 CrossRef CAS PubMed.
- A. A. Tsygankov, S. N. Kosourov, I. V. Tolstygina, M. L. Ghirardi and M. Seibert, Int. J. Hydrogen Energy, 2006, 31, 1574–1584 CrossRef CAS.
- M. J. Barbosa, J. Rocha, J. Tramper and R. H. Wijffels, J. Biotechnol., 2001, 85, 25–33 CrossRef CAS PubMed.
- H.-D. Lin, B.-H. Liu, T.-T. Kuo, H.-C. Tsai, T.-Y. Feng, C.-C. Huang and L.-F. Chien, Bioresour. Technol., 2013, 143, 154–162 CrossRef CAS PubMed.
- K. G. Kenchappa, T. Guerra, X. Qian, S. Zhang, D. Bryant and G. C. Dismukes, Energy Environ. Sci., 2013, 6, 3722–3731 Search PubMed.
- M. F. Demirbas, Appl. Energy, 2011, 88, 3473–3480 CrossRef CAS.
- A. Ahmad, N. Yasin, C. Derek and J. Lim, Renewable Sustainable Energy Rev., 2011, 15, 584–593 CrossRef CAS.
- Y. Feng, C. Li and D. Zhang, Bioresour. Technol., 2011, 102, 101–105 CrossRef CAS PubMed.
- J. M. Gordon and J. E. Polle, Appl. Microbiol. Biotechnol., 2007, 76, 969–975 CrossRef CAS PubMed.
- G. W. Huber, S. Iborra and A. Corma, Chem. Rev., 2006, 106, 4044–4098 CrossRef CAS PubMed.
- B. H. Gebreslassie, R. Waymire and F. You, AIChE J., 2013, 59, 1599–1621 CrossRef CAS.
- R. Luque, J. C. Lovett, B. Datta, J. Clancy, J. M. Campelo and A. A. Romero, Energy Environ. Sci., 2010, 3, 1706–1721 CAS.
- F. Ma and M. A. Hanna, Bioresour. Technol., 1999, 70, 1–15 CrossRef CAS.
- F. Mairet, O. Bernard, P. Masci, T. Lacour and A. Sciandra, Bioresour. Technol., 2011, 102, 142–149 CrossRef CAS PubMed.
- A. Demirbas and M. Fatih Demirbas, Energy Convers. Manage., 2011, 52, 163–170 CrossRef.
- A. F. Clarens, H. Nassau, E. P. Resurreccion, M. A. White and L. M. Colosi, Environ. Sci. Technol., 2011, 45, 7554–7560 CrossRef CAS PubMed.
- I. Rawat, R. Ranjith Kumar, T. Mutanda and F. Bux, Appl. Energy, 2013, 103, 444–467 CrossRef CAS.
- T. Shirvani, X. Yan, O. R. Inderwildi, P. P. Edwards and D. A. King, Energy Environ. Sci., 2011, 4, 3773–3778 CAS.
- N. Pragya, K. K. Pandey and P. Sahoo, Renewable Sustainable Energy Rev., 2013, 24, 159–171 CrossRef CAS.
- H. Tang, N. Abunasser, M. Garcia, M. Chen, K. Simon Ng and S. O. Salley, Appl. Energy, 2011, 88, 3324–3330 CrossRef CAS.
- C. J. Rhodes, Sci. Prog., 2009, 92, 39–90 CrossRef CAS PubMed.
- D. B. Ghonasgi, E. L. Sughrue, J. Yao and X. Xu, US Pat., 7, 626, 063, 2009.
- L. Li, E. Coppola, J. Rine, J. L. Miller and D. Walker, Energy Fuels, 2010, 24, 1305–1315 CrossRef CAS.
- J. C. Serrano-Ruiz, R. Luque and A. Sepúlveda-Escribano, Chem. Soc. Rev., 2011, 40, 5266–5281 RSC.
- M. J. Haas, A. J. McAloon, W. C. Yee and T. A. Foglia, Bioresour. Technol., 2006, 97, 671–678 CrossRef CAS PubMed.
- J. C. Serrano-Ruiz, E. V. Ramos-Fernández and A. Sepúlveda-Escribano, Energy Environ. Sci., 2012, 5, 5638–5652 CAS.
- G. W. Huber, P. O'Connor and A. Corma, Appl. Catal., A, 2007, 329, 120–129 CrossRef CAS.
- G. W. Huber and A. Corma, Angew. Chem., Int. Ed., 2007, 46, 7184–7201 CrossRef CAS PubMed.
- K. Sunde, A. Brekke and B. Solberg, Energies, 2011, 4, 845–877 CrossRef CAS.
- B. Donnis, R. G. Egeberg, P. Blom and K. G. Knudsen, Top. Catal., 2009, 52, 229–240 CrossRef CAS.
- J. Walendziewski, M. Stolarski, R. Łużny and B. Klimek, Fuel Process. Technol., 2009, 90, 686–691 CrossRef CAS.
- H. Taher, S. Al-Zuhair, A. H. Al-Marzouqi, Y. Haik and M. M. Farid, Enzyme Res., 2011, 468292, DOI:10.4061/2011/468292.
- M. Koberg, M. Cohen, A. Ben-Amotz and A. Gedanken, Bioresour. Technol., 2011, 102, 4265–4269 CrossRef CAS PubMed.
- A. Ranjan, C. Patil and V. S. Moholkar, Ind. Eng. Chem. Res., 2010, 49, 2979–2985 CrossRef CAS.
- P. D. Patil, V. G. Gude, A. Mannarswamy, P. Cooke, S. Munson-McGee, N. Nirmalakhandan, P. Lammers and S. Deng, Bioresour. Technol., 2011, 102, 1399–1405 CrossRef CAS PubMed.
- P. D. Patil, H. Reddy, T. Muppaneni, A. Mannarswamy, T. Schuab, F. O. Holguin, P. Lammers, N. Nirmalakhandan, P. Cooke and S. Deng, Green Chem., 2012, 14, 809–818 RSC.
- A. Loupy, A. Petit, M. Ramdani, C. Yvanaeff, M. Majdoub, B. Labiad and D. Villemin, Can. J. Chem., 1993, 71, 90–95 CrossRef CAS.
- H. Yuan, B. Yang and G. Zhu, Energy Fuels, 2008, 23, 548–552 CrossRef.
- P. Lidström, J. Tierney, B. Wathey and J. Westman, Tetrahedron, 2001, 57, 9225–9283 CrossRef.
- L. Perreux and A. Loupy, Tetrahedron, 2001, 57, 9199–9223 CrossRef CAS.
- J. Tierney and P. Lidström, Microwave assisted organic synthesis, Wiley-Blackwell, 2009 Search PubMed.
- O. Armas, R. Ballesteros, F. Martos and J. Agudelo, Fuel, 2005, 84, 1011–1018 CrossRef CAS.
- M. Y. Selim and M. T. Ghannam, SAE Paper, 2007, 0132 Search PubMed.
- T. Kannan and R. Marappan, J. Appl. Sci., 2011, 11, 2961–2967 CrossRef CAS.
- T. Namioka, K. Yoshikawa, M. Takeshita and K. Fujiwara, Appl. Energy, 2012, 93, 517–522 CrossRef.
- P. Sungwornpatansakul, J. Hiroi, Y. Nigahara, T. K. Jayasinghe and K. Yoshikawa, Fuel Process. Technol., 2013, 116, 1–8 CrossRef CAS.
- S. A. Scott, M. P. Davey, J. S. Dennis, I. Horst, C. J. Howe, D. J. Lea-Smith and A. G. Smith, Curr. Opin. Biotechnol., 2010, 21, 277–286 CrossRef CAS PubMed.
- L. Lardon, A. Hélias, B. Sialve, J.-P. Steyer and O. Bernard, Environ. Sci. Technol., 2009, 43, 6475–6481 CrossRef CAS PubMed.
- A. F. Clarens, E. P. Resurreccion, M. A. White and L. M. Colosi, Environ. Sci. Technol., 2010, 44, 1813–1819 CrossRef CAS PubMed.
- J. Yang, M. Xu, X. Zhang, Q. Hu, M. Sommerfeld and Y. Chen, Bioresour. Technol., 2011, 102, 159–165 CrossRef CAS PubMed.
- K. Sander and G. S. Murthy, Int. J. Life Cycle Assess., 2010, 15, 704–714 CrossRef CAS.
- P. K. Campbell, T. Beer and D. Batten, Bioresour. Technol., 2011, 102, 50–56 CrossRef CAS PubMed.
- K. Soratana and A. E. Landis, Bioresour. Technol., 2011, 102, 6892–6901 CrossRef CAS PubMed.
- B. S. Sturm and S. L. Lamer, Appl. Energy, 2011, 88, 3499–3506 CrossRef CAS.
- L. F. Wu, P. C. Chen, A. P. Huang and C. M. Lee, Bioresour. Technol., 2012, 113, 14–18 CrossRef CAS PubMed.
- P. Collet, A. Hélias, L. Lardon, M. Ras, R.-A. Goy and J.-P. Steyer, Bioresour. Technol., 2011, 102, 207–214 CrossRef CAS PubMed.
- A. L. Stephenson, E. Kazamia, J. S. Dennis, C. J. Howe, S. A. Scott and A. G. Smith, Energy Fuels, 2010, 24, 4062–4077 CrossRef CAS.
- R. Chowdhury, S. Viamajala and R. Gerlach, Bioresour. Technol., 2012, 108, 102–111 CrossRef CAS PubMed.
- D. R. Georgianna and S. P. Mayfield, Nature, 2012, 488, 329–335 CrossRef CAS PubMed.
- V. H. Smith, B. S. Sturm, F. J. Denoyelles and S. A. Billings, Trends Ecol. Evol., 2010, 25, 301–309 CrossRef PubMed.
- N. Moazami, A. Ashori, R. Ranjbar, M. Tangestani, R. Eghtesadi and A. S. Nejad, Biomass Bioenergy, 2012, 39, 449–453 CrossRef CAS.
- B. J. Krohn, C. V. McNeff, B. Yan and D. Nowlan, Bioresour. Technol., 2011, 102, 94–100 CrossRef CAS PubMed.
- V. Jordan and B. Gutsche, Chemosphere, 2001, 43, 99–105 CrossRef CAS PubMed.
- Y. C. Sharma, B. Singh and J. Korstad, Green Chem., 2011, 13, 2993–3006 RSC.
- R. Harun, M. K. Danquah and G. M. Forde, J. Chem. Technol. Biotechnol., 2010, 85, 199–203 CAS.
- S.-H. Ho, S.-W. Huang, C.-Y. Chen, T. Hasunuma, A. Kondo and J.-S. Chang, Bioresour. Technol., 2013, 135, 191–198 CrossRef CAS PubMed.
- S. G. Wi, H. J. Kim, S. A. Mahadevan, D.-J. Yang and H.-J. Bae, Bioresour. Technol., 2009, 100, 6658–6660 CrossRef CAS PubMed.
- J. J. Yoon, Y. J. Kim, S. H. Kim, H. J. Ryu, J. Y. Choi, G. S. Kim and M. K. Shin, Adv. Mater. Res., 2010, 93, 463–466 CrossRef.
- C. A. Cardona and Ó. J. Sánchez, Bioresour. Technol., 2007, 98, 2415–2457 CrossRef CAS PubMed.
- R. P. John, G. Anisha, K. M. Nampoothiri and A. Pandey, Bioresour. Technol., 2011, 102, 186–193 CrossRef CAS PubMed.
- F. S. Eshaq, M. N. Ali and M. K. Mohd, Int. J. Eng. Sci. Tech., 2011, 3, 1749–1755 Search PubMed.
- M. K. Lam and K. T. Lee, Biotechnol. Adv., 2012, 30, 673–690 CrossRef CAS PubMed.
- M. Martín and I. E. Grossmann, AIChE J., 2013, 59, 2872–2883 CrossRef.
- K. Severson, M. Martín and I. E. Grossmann, AIChE J., 2012, 59, 834–844 CrossRef.
- J. S. Burlew, Algal culture from laboratory to pilot plant, 1953 Search PubMed.
- P. J. l. B. Williams and L. M. Laurens, Energy Environ. Sci., 2010, 3, 554–590 CAS.
- C. F. Murphy and D. T. Allen, Environ. Sci. Technol., 2011, 45, 5861–5868 CrossRef CAS PubMed.
- J. R. Benemann and W. J. Oswald, Systems and economic analysis of microalgae ponds for conversion of CO2 to biomass, Final report, California Univ., Dept. of Civil Engineering, Berkeley, CA (United States), 1996 Search PubMed.
- A. Shilton, N. Powell and B. Guieysse, Curr. Opin. Biotechnol., 2012, 23, 884–889 CrossRef CAS PubMed.
- N. Powell, A. Shilton, S. Pratt and Y. Chisti, Water Sci. Technol., 2011, 63, 1689–1694 CrossRef CAS PubMed.
- W. Mulbry, E. K. Westhead, C. Pizarro and L. Sikora, Bioresour. Technol., 2005, 96, 451–458 CrossRef CAS PubMed.
- J. Park, R. Craggs and A. Shilton, Bioresour. Technol., 2011, 102, 35–42 CrossRef CAS PubMed.
- L. Wang, M. Min, Y. Li, P. Chen, Y. Chen, Y. Liu, Y. Wang and R. Ruan, Appl. Biochem. Biotechnol., 2010, 162, 1174–1186 CrossRef CAS PubMed.
- L. Wang, Y. Li, P. Chen, M. Min, Y. Chen, J. Zhu and R. R. Ruan, Bioresour. Technol., 2010, 101, 2623–2628 CrossRef CAS PubMed.
- W. Zhou, Y. Li, M. Min, B. Hu, P. Chen and R. Ruan, Bioresour. Technol., 2011, 102, 6909–6919 CrossRef CAS PubMed.
- L. Wang, Y. C. Li, P. Chen, M. Min, Y. F. Chen, J. Zhu and R. Ruan, Bioresour. Technol., 2010, 101, 2623–2628 CrossRef CAS PubMed.
- M. Singh, D. L. Reynolds and K. C. Das, Bioresour. Technol., 2011, 102, 10841–10848 CrossRef CAS PubMed.
- L. Wang, Y. K. Wang, P. Chen and R. Ruan, Appl. Biochem. Biotechnol., 2010, 162, 2324–2332 CrossRef CAS PubMed.
- T. Bruton, H. Lyons, T. Lerat, M. Stanley and M. B. Rasmussen, Sustainable Energy, Glasnevin, Ireland, 2009 Search PubMed.
- T. M. Mata, A. A. Martins and N. S. Caetano, Renewable Sustainable Energy Rev., 2010, 14, 217–232 CrossRef CAS.
- S. Yokoyama, K. Jonouchi and K. Imou, Proc. World Acad. Sci. Eng. Tech., 2007, 4, 320–323 Search PubMed.
- B. D. Burks and M. M. Minnis, Onsite Wastewater and Treatment Systems, Madison, Hogarth House, 1994 Search PubMed.
- S. A. Bradford, E. Segal, W. Zheng, Q. Q. Wang and S. R. Hutchins, J. Environ. Qual., 2008, 37, S97–S115 CrossRef PubMed.
- J. C. Barker, J. P. Zublena and F. R. Walls, Livestock and poultry manure characteristics, 2001 Search PubMed.
- K. Yetilmezsoy and S. Sakar, J. Hazard. Mater., 2008, 153, 532–543 CrossRef CAS PubMed.
- A. Millmier, J. Lorimor, C. Hurburgh, C. Fulhage, J. Hattey and H. Zhang, Trans. ASABE, 2000, 43, 903–908 CrossRef CAS.
- R. Abou-Shanab, M.-K. Ji, H.-C. Kim, K.-J. Paeng and B.-H. Jeon, J. Environ. Manage., 2013, 115, 257–264 CrossRef CAS PubMed.
- S. Chinnasamy, A. Bhatnagar, R. W. Hunt and K. C. Das, Bioresour. Technol., 2010, 101, 3097–3105 CrossRef CAS PubMed.
- S. Sen and G. N. Demirer, Water Res., 2003, 37, 1868–1878 CrossRef CAS PubMed.
- K. P. M. Mosse, A. F. Patti, E. W. Christen and T. R. Cavagnaro, Aust. J. Grape Wine Res., 2011, 17, 111–122 CrossRef CAS.
- G. Durai and M. Rajasimman, J. Environ. Sci. Technol., 2011, 4, 1–17 CrossRef CAS.
- A. H. Slade, D. J. Gapes, T. R. Stuthridge, S. M. Anderson, P. H. Dare, H. G. W. Pearson and M. Dennis, Water Sci. Technol., 2004, 50, 131–139 CAS.
- B. Y. Ammary, Afr. J. Biotechnol., 2004, 3, 236–238 CrossRef CAS.
- S. Cho, T. T. Luong, D. Lee, Y. K. Oh and T. Lee, Bioresour. Technol., 2011, 102, 8639–8645 CrossRef CAS PubMed.
- K. Yetilmezsoy and S. Sakar, J. Hazard. Mater., 2008, 151, 547–558 CrossRef CAS PubMed.
- K. Yetilmezsoy and Z. Sapci-Zengin, J. Hazard. Mater., 2009, 166, 260–269 CrossRef CAS PubMed.
- A. Montusiewicz, M. Lebiocka, A. Rozej, E. Zacharska and L. Pawlowski, Bioresour. Technol., 2010, 101, 3466–3473 CrossRef CAS PubMed.
- H. M. El-Mashad and R. H. Zhang, Trans. ASABE, 2007, 50, 1815–1822 CrossRef.
- K. Lee and C. G. Lee, Biotechnol. Bioprocess Eng., 2001, 6, 194–199 CrossRef CAS.
- A. Ruiz-Marin, L. G. Mendoza-Espinosa and T. Stephenson, Bioresour. Technol., 2010, 101, 58–64 CrossRef CAS PubMed.
- M. Khan and N. Yoshida, Bioresour. Technol., 2008, 99, 575–582 CrossRef CAS PubMed.
- L. E. Gonzalez, R. O. Canizares and S. Baena, Bioresour. Technol., 1997, 60, 259–262 CrossRef CAS.
- H. Su, Y. Zhang, C. Zhang, X. Zhou and J. Li, Bioresour. Technol., 2011, 102, 9884–9890 CrossRef CAS PubMed.
- S. Aslan and I. K. Kapdan, Ecol. Eng., 2006, 28, 64–70 CrossRef.
- Q. X. Kong, L. Li, B. Martinez, P. Chen and R. Ruan, Appl. Biochem. Biotechnol., 2010, 160, 9–18 CrossRef CAS PubMed.
- J. P. Hernandez, L. E. de-Bashan and Y. Bashan, Enzyme Microb. Technol., 2006, 38, 190–198 CrossRef CAS.
- E. D. Zhang, B. Wang, Q. H. Wang, S. B. Zhang and B. D. Zhao, Bioresour. Technol., 2008, 99, 3787–3793 CrossRef CAS PubMed.
- S. M. Phang, M. S. Miah, B. G. Yeoh and M. A. Hashim, J. Appl. Phycol., 2000, 12, 395–400 CrossRef.
- R. J. Craggs, P. J. McAuley and V. J. Smith, Water Res., 1997, 31, 1701–1707 CrossRef CAS.
- R. J. Craggs, V. J. Smith and P. J. McAuley, Water Sci. Technol., 1995, 31, 151–160 CrossRef CAS.
- E. Valenzuela-Espinoza, R. Millan-Nunez and F. Nunez-Cebrero, Aquacultural Engineering, 1999, 20, 135–147 CrossRef.
- Y. Su, A. Mennerich and B. Urban, Bioresour. Technol., 2012, 124, 157–162 CrossRef CAS PubMed.
- R. Quiros, The nitrogen to phosphorus ratio for lakes: a cause or a consequence of aquatic biology?, ed. A. F. Cirelli and G. C. Marquisa, El Agua en Iberoamerica: De la Limnologia a la Gestion en Sudamerica, CYTED XVII, Centro de Estudios Transdiciplinarios del Agua, Facultad de Veterinaria, Universidad de Buenos Aires, Buenos Aires, 2002, pp. 11–26 Search PubMed.
- J. Shi, B. Podola and M. Melkonian, J. Appl. Phycol., 2007, 19, 417–423 CrossRef CAS.
- A. Bhatnagar, M. Bhatnagar, S. Chinnasamy and K. C. Das, Appl. Biochem. Biotechnol., 2010, 161, 523–536 CrossRef CAS PubMed.
- O. Perez-Garcia, Y. Bashan and M. E. Puente, J. Phycol., 2011, 47, 190–199 CrossRef.
- G. Y. Rhee, Limnol. Oceanogr., 1978, 23, 10–25 CrossRef CAS.
- A. Lavoie and J. de la Noue, Water Res., 1985, 19, 1437–1442 CrossRef CAS.
- S. Sawayama, T. Minowa, Y. Dote and S. Yokoyama, Appl. Biochem. Biotechnol., 1992, 29, 145–148 Search PubMed.
- B. D. Tripathi and S. C. Shukla, Environ. Pollut., 1991, 69, 69–78 CrossRef CAS PubMed.
- E. L. Olguin, S. Galicia, G. Mercado and T. Perez, J. Appl. Phycol., 2003, 15, 249–257 CrossRef CAS.
- G. Yu, Y. Zhang, L. Schideman, T. Funk and Z. Wang, Trans. ASABE, 2011, 54, 239–246 CrossRef.
- U. Jena, N. Vaidyanathan, S. Chinnasamy and K. Das, Bioresour. Technol., 2011, 102, 3380–3387 CrossRef CAS PubMed.
- P. Biller, A. B. Ross, S. Skill, A. Lea-Langton, B. Balasundaram, C. Hall, R. Riley and C. Llewellyn, Algal Res., 2012, 1, 70–76 CrossRef CAS.
- Z. Du, B. Hu, X. Ma, Y. Cheng, Y. Liu, X. Lin, Y. Wan, H. Lei, P. Chen and R. Ruan, Bioresour. Technol., 2013, 130, 777–782 CrossRef CAS PubMed.
- T. Minowa and S. Sawayama, Fuel, 1999, 78, 1213–1215 CrossRef CAS.
- Y. Zhou, L. Schideman, G. Yu and Y. Zhang, Energy Environ. Sci., 2013, 6, 3765–3779 CAS.
- R. Porra, W. Thompson and P. Kriedemann, Biochim. Biophys. Acta, Bioenerg., 1989, 975, 384–394 CrossRef CAS.
- P. Das, S. S. Aziz and J. P. Obbard, Renewable Energy, 2011, 36, 2524–2528 CrossRef CAS.
- J. R. Oyler, US Pat., 7905930 2011.
- W. Xiong, C. Gao, D. Yan, C. Wu and Q. Wu, Bioresour. Technol., 2010, 101, 2287–2293 CrossRef CAS PubMed.
- W. Zhou, M. Min, Y. Li, B. Hu, X. Ma, Y. Cheng, Y. Liu, P. Chen and R. Ruan, Bioresour. Technol., 2012, 110, 448–455 CrossRef CAS PubMed.
- P. Lau, N. Tam and Y. Wong, Environ. Pollut., 1995, 89, 59–66 CrossRef CAS.
- M.-K. Ji, R. Abou-Shanab, S.-H. Kim, E.-S. Salama, S.-H. Lee, A. N. Kabra, Y.-S. Lee, S. Hong and B.-H. Jeon, Ecol. Eng., 2013, 58, 142–148 CrossRef.
- R. Abou-Shanab, S.-H. Kim, M.-K. Ji, S.-H. Lee, H.-S. Roh and B.-H. Jeon, J. Renewable Sustainable Energy, 2013, 5, 052006 CrossRef.
- Z. Du, Y. Li, X. Wang, Y. Wan, Q. Chen, C. Wang, X. Lin, Y. Liu, P. Chen and R. Ruan, Bioresour. Technol., 2011, 102, 4890–4896 CrossRef CAS PubMed.
- G. Sivakumar, J. Xu, R. W. Thompson, Y. Yang, P. Randol-Smith and P. J. Weathers, Bioresour. Technol., 2012, 107, 1–9 CrossRef CAS PubMed.
- M.-K. Ji, R. Abou-Shanab, J.-H. Huang, T. C. Timmes, H.-C. Kim, Y.-K. Oh and B.-H. Jeon, J. Environ. Eng., 2013, 139, 1198–1205 CrossRef CAS.
- M.-K. Ji, H.-C. Kim, V. R. Sapireddy, H.-S. Yun, R. Abou-Shanab, J. Choi, W. Lee, T. C. Timmes and B.-H. Jeon Inamuddin, Appl. Biochem. Biotechnol., 2013, 97, 2701–2710 CAS.
- M. S. A. Rahaman, L.-H. Cheng, X.-H. Xu, L. Zhang and H.-L. Chen, Renewable Sustainable Energy Rev., 2011, 15, 4002–4012 CrossRef.
- J. Pires, M. Alvim-Ferraz, F. Martins and M. Simoes, Renewable Sustainable Energy Rev., 2012, 16, 3043–3053 CrossRef CAS.
- A. Kirrolia, N. R. Bishnoi and R. Singh, Renewable Sustainable Energy Rev., 2013, 20, 642–656 CrossRef CAS.
- K. Sudhakar and M. Premalatha, Journal of Sustainable Energy Environment, 2012, 3, 59–62 Search PubMed.
- E. Waltz, Nat. Biotechnol., 2013, 31, 12 CrossRef.
- R. K. W. Tham and W. Zhang, Biofuels, 2012, 3, 5–8 CrossRef CAS.
| This journal is © The Royal Society of Chemistry 2014 |