Selective interaction of methylindoloquinolines with DNA
Prasun Ghosh and
Pradipta Purkayastha*
Indian Institute of Science Education and Research (IISER) Kolkata, Mohanpur Campus, Mohanpur 741252, India. E-mail: pradiptp@gmail.com; Fax: +91 33 25873121; Tel: +91 33 25873020
First published on 12th May 2014
Abstract
Interaction of three medically potent indoloquinoline drugs with model DNA has been studied. The three indoloquinoline derivatives chosen in the present work are 5-methyl-5H-indolo[3,2-c]-quinoline (MIQ), 8-chloro-5-methyl-5H-indolo[3,2-c]quinoline (CMIQ), and 2,8-dichloro-5-methyl-5H-indolo[3,2-c]quinoline (DCMIQ). The compounds are found to interact with DNA in their ground electronic states. The mode of interaction has been studied using steady state fluorescence spectroscopy coupled with circular dichroism (CD) spectroscopy. The change in fluorescence properties of the drugs confirmed their binding interaction with the DNA double helix whereas the CD signals were used to analyze the structural changes of the DNA due to the process. Fluorescence quenching studies and CD spectra indicate that the drugs intercalate into the DNA. Moreover, it has been noticed that the chloro derivatives of the indoloquinoline drugs bind more efficiently to the DNA due to hydrophobic interaction and thus enhance the DNA melting temperature. Sequenced DNA oligos have been used to confirm the selectivity in binding of the drug molecules to DNA.
Introduction
Deoxyribonucleic acid (DNA) encodes genetic instructions from one generation to the other that control the nature and function of each individual cell.1,2 This enables performance of multidimensional research on DNA and its interaction with small molecules that yield important information regarding the development of therapeutic agents in controlling gene expression, mismatch repairing, etc.3–6 The affinity, strength, and mode of interaction of small molecules with DNA are governed by various structural and electronic factors of both the candidates.7 In the present study, the model DNA that has been used is derived from calf thymus (ctDNA) having relatively low protein content with a highly polymerized skeleton. ctDNA has successfully been used in many drug–DNA interaction studies.8–10 In general there are typically three modes of binding interaction of DNA with small molecules: (i) electrostatic binding, (ii) groove binding, and (iii) intercalation. Among them, intercalative binding is the most effective and efficient for drugs targeted to DNA. In this mode the structure of DNA is directly affected by replacing the hydrogen bonding between the base pairs.11Results from interaction of small molecules with polymeric DNA provide idea on a broader aspect but fails to give any information about selectivity of drug–DNA encounter. To know about selectivity and specificity of binding in detail, particular sequence/s of oligos can be chosen.12 Information on selectivity is very important for synthesis of new drugs since consequentially detailed mechanism on the action of drugs within the cell can be understood. Keeping this sensitive aspect under consideration we chose the following two small sequenced oligos:
5′-CAGCTCGAGTTTTCTCGAGCTG-3′-primer2 (P2)
5′-CGCAATTGGCTTTTGCCAATTGCG-3′-primer1 (P1)
P2 and P1 contain the specific –GCTCG– and –AATT– sequences, respectively.13
Indoloquinoline and some of its derivatives were first extracted from the African medicinal plant Cryptolepis sanguinolenta.14,15 They are well known for their broad spectrum of biological activities including antiparasitic, antifungal, antibacterial, cytotoxicity, anti-inflammatory, and antihyperglycaemic.16,17 These compounds are known to inhibit DNA replication and transcription16 and also exhibit strong antiplasmodial activity.17 A number of 2-substituted indoloquinolines have been synthesized and evaluated in antifungal screens, and several have been shown to increase potency and expand the antifungal spectrum of cryptolepine.18 Indoloquinolines and their methyl derivatives are known to possess suitable skeleton for DNA intercalation as these molecules are planar and hydrophobic in nature resulting into dramatic changes in DNA conformation and inhibiting DNA replication and transcription.19–21
In our laboratory we have long been involved in studying the photophysical properties of the parent cryptosanguinolentine or 5-methyl-5H-indolo[3,2-c]quinoline (MIQ) and two of its chloro derivatives, 8-chloro-5-methyl-5H-indolo[3,2-c]quinoline (CMIQ) and 2,8-dichloro-5-methyl-5H-indolo[3,2-c]quinoline (DCMIQ) in homogeneous and microheterogeneous media.21,22 The molecular structures of these compounds are shown in Scheme 1. All of them exist in their neutral and zwitterionic forms in aqueous solution (Scheme 2). In the excited state, the zwitterionic species is predominant over the neutral ones. Formation of two types of solvated species is inferred from our previous solvatochromatic study.22 Scheme 2 suggests formation of intermolecular stacks between the zwitterions due to coulombic interactions. This stacking is possible in both ground and excited states.23,24 We showed that the equilibrium of dimer formation in the ground state as well as excited state can be modulated through application of cyclodextrins of different dimensions.23 The structure and potency in biological activity of these molecules encouraged us to investigate the interaction of these compounds with model and tactically sequenced DNA. In this study we have also shown how the position of chloro substituent/s in parent MIQ modulates the drug–DNA interaction in polymeric ctDNA as well as in small oligos.
 | ||
| Scheme 1 Representative structures of (A) 5-methyl-5H-indolo[3,2-c]quinoline (MIQ), (B) 8-chloro-5-methyl-5H-indolo[3,2-c]quinoline (CMIQ), and (C) 2,8-dichloro-5-methyl-5H-indolo[3,2-c]quinoline (DCMIQ). | ||
 | ||
| Scheme 2 Equilibrium between the neutral and the zwitterionic forms of MIQ. | ||
Results and discussion
Ground state interaction between the indoloquinolines and ctDNA
The absorption spectra of MIQ, CMIQ and DCMIQ in aqueous buffer show similar spectral structures in the range 220 nm to 400 nm. These bands are due to π–π* and n–π* electronic transitions in the molecules.22 Absorption of these dyes in the said spectral region overlaps with that of DNA. Thus, to eliminate the effect of the absorbance of DNA we added a calculated amount of DNA in both reference and probe solutions used in the double-beam spectrophotometer. Fig. 1 shows the absorption spectra of the dyes in Tris–buffer at pH 7.6 in presence of various concentrations of ctDNA. The concentration of MIQ, CMIQ and DCMIQ was kept at 25 μM.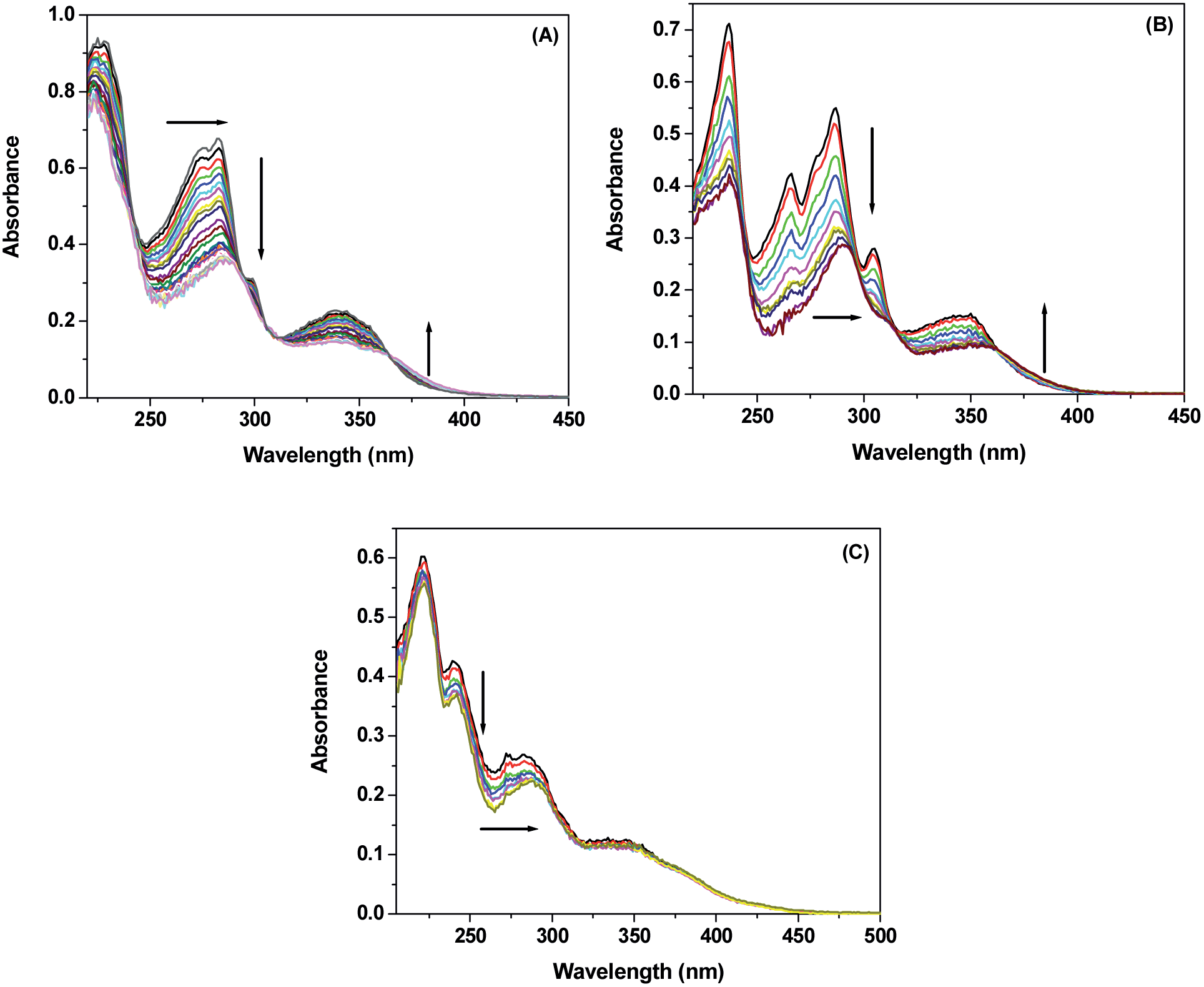 | ||
| Fig. 1 Absorption spectra of (A) MIQ, (B) CMIQ, and (C) DCMIQ in presence of different ctDNA concentrations. Arrows indicate spectral change in each case with increase in ctDNA concentration. | ||
Addition of ctDNA leads to noticeable changes in the absorption spectra of the compounds. The absorbance at ∼280 nm decreases along with structural changes and an appreciable bathochromic shift of ∼5 nm was observed. Concomitant formation of a new band at 375 nm is also observed. Formation of the new higher wavelength absorption band is most prominent in case of MIQ and least for DCMIQ. The bathochromic shift in absorption band at a particular wavelength suggests strong interaction between the DNA and the drugs.25,26 Absence of a clear isosbestic point in the spectral changes for any of the three dyes implies that 1![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :1 drug:DNA stoichiometry is probably not maintained during the binding process and/or there is more than one type of binding. This prevented us from determining the binding constants for the drug molecules with ctDNA from the absorption studies.
:1 drug:DNA stoichiometry is probably not maintained during the binding process and/or there is more than one type of binding. This prevented us from determining the binding constants for the drug molecules with ctDNA from the absorption studies.
Excited state processes for drug–ctDNA interactions
The emission spectra of MIQ, CMIQ and DCMIQ in aqueous buffer show a broad band having maximum intensity around 460 nm and a shoulder around 440 nm.22,23 Addition of ctDNA results into an appreciable quenching of fluorescence of the three compounds. For the same concentration of added ctDNA to the same amount of the fluorophores, quenching of fluorescence was in the order DCMIQ > CMIQ > MIQ. Saturation of fluorescence quenching was observed in the order MIQ > CMIQ > DCMIQ based on the amount of added ctDNA. The results show that a strong binding interaction takes place between the probe and the DNA. Relative change in fluorescence as a result of added ctDNA to the fluorophores (F/F0) has been plotted as a function of ctDNA concentration and is shown in Fig. 2A. The non-linear feature of the fit could not provide the respective binding constants but the extent of quenching inferred strong guest–host interaction. From the plots it can easily be concluded that the presence of the chloro groups in the parent MIQ increases the binding efficiency of the compounds with ctDNA. Presumably the addition of the chloro functionality to the parent MIQ makes the resultant molecules more hydrophobic resulting into a stronger binding in the hydrophobic pockets of the DNA. Thus, binding efficiency is highest in case of DCMIQ compared to the other two derivatives and hence relative fluorescence quenching is highest in this case.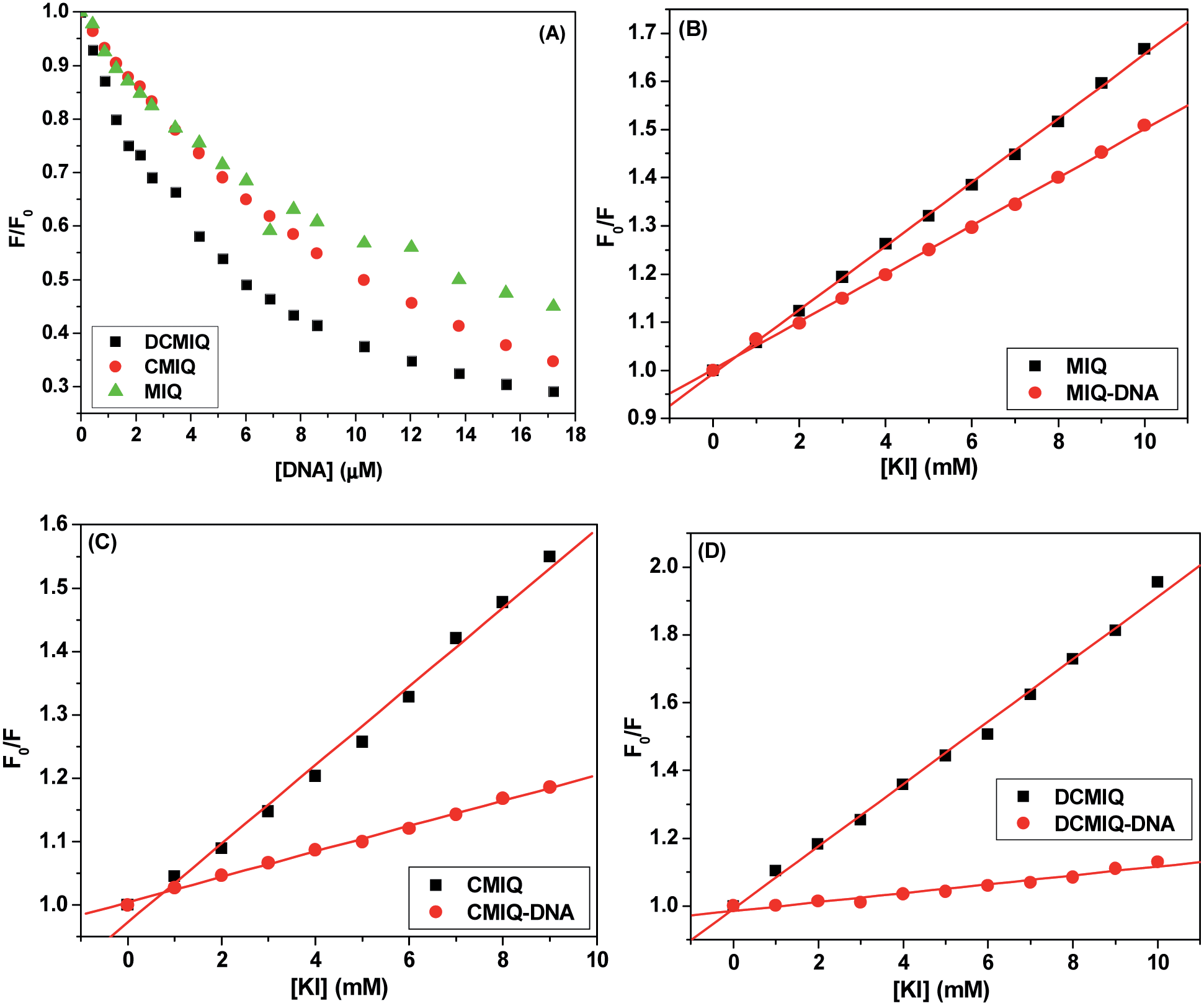 | ||
| Fig. 2 Relative extent of fluorescence quenching for DCMIQ, CMIQ, and MIQ as a function of ctDNA concentration (A), Stern–Volmer plots for fluorescence quenching of MIQ (B), CMIQ (C) and DCMIQ (D) by KI in blank aqueous and ctDNA environments. In each case concentration of the fluorophore used is 10 μM. The excitation wavelength was 290 nm. | ||
To further clarify the mode of binding of MIQ, CMIQ and DCMIQ with ctDNA, their fluorescence quenching in the DNA environment was studied using KI as a quencher. It is known that entrapped small molecules inside DNA double helix due to intercalation gets protection from externally added ionic quenchers.8,27 Possible electrostatic repulsion between the negatively charged quencher and the negative phosphate backbone of DNA reduces the possibility of interaction between the anionic quencher and intercalated drug. In contrast, electrostatic binding and groove binding of guest molecules with DNA leave them exposed to the bulk aqueous phase and does not appreciably hinder the approach of the quencher.8 Thus, the intercalative mode of binding of a fluorophore with DNA should lead to lower extent of quenching of its fluorescence compared to that without the host. Moreover, in case of groove binding the quenching efficiency of KI is higher in ctDNA environment compared to that without it because KI itself is a strong electrolyte that can release the groove-bound fluorophore.8
Table 1 provides the calculated Stern–Volmer constants for free and ctDNA bound probes on interaction with KI. The Ksv values decrease appreciably for all the three indoloquinolines. Such high degree of reduction in quenching constant indicates intercalative mode of binding of these molecules with DNA. Degree of reduction in quenching constant follows the order DCMIQ > CMIQ > MIQ indicating that DCMIQ binds most strongly and MIQ is the weakest binder. The results intuitively infer that introduction of chloro groups increases the binding capacity of the compounds.
| Fluorophore | Ksv in blank (M−1) | Ksv in drug–ctDNA complex (M−1) | % reduction in Ksv |
|---|---|---|---|
| MIQ | 66.4 | 29.3 | 55.9 |
| CMIQ | 61.9 | 20.1 | 67.5 |
| DCMIQ | 92.1 | 13.1 | 85.8 |
To provide more evidence for our findings we did steady state fluorescence anisotropy measurements with the three compounds with increase in ctDNA concentration (Fig. 3). The method is very sensitive to motional restriction offered by the immediate microenvironment of the fluorophore.28–31 Enhancement in anisotropy (r) to 0.05, 0.07 and 0.11 was recorded in case of MIQ, CMIQ and DCMIQ, respectively on addition of ctDNA. The results show that the chloro derivatives provide stronger binding to ctDNA corroborating with the ground state studies. Hence molecular hydrophobicity decides the strength of binding to DNA double helix.22
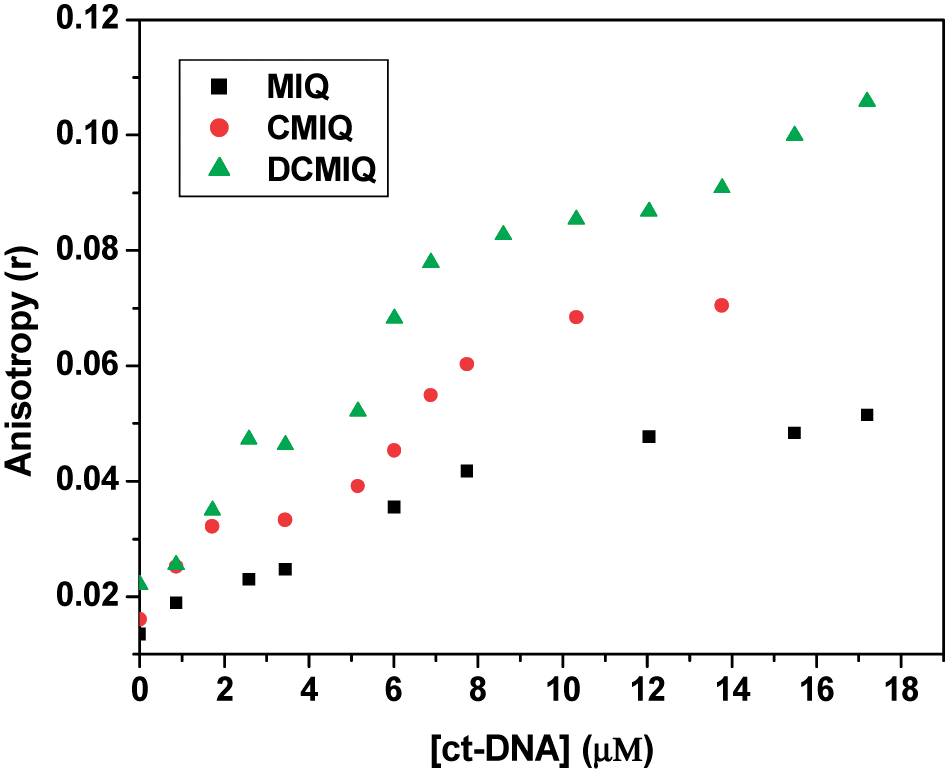 | ||
| Fig. 3 Change in steady state fluorescence anisotropy (r) of MIQ, CMIQ and DCMIQ as a function of ctDNA concentration (λex = 290 nm and λmonitored = λmaxem). | ||
It is known that intercalation leads to different degrees of structural deformation of DNA depending on the intercalating substance. Better the intercalation, more will be the structural deformation that may lead to stabilization of the DNA host. Urea is known to be a strong hydrogen bonding candidate to the DNA base pairs and thus can displace the intercalated compound from DNA helix.32,33 The degree of deformation in the DNA helix can thus be measured by applying urea to the DNA–drug intercalate. We have shown spectroscopically in Fig. 4 that urea releases the intercalated drugs in the order DCMIQ > CMIQ > MIQ.
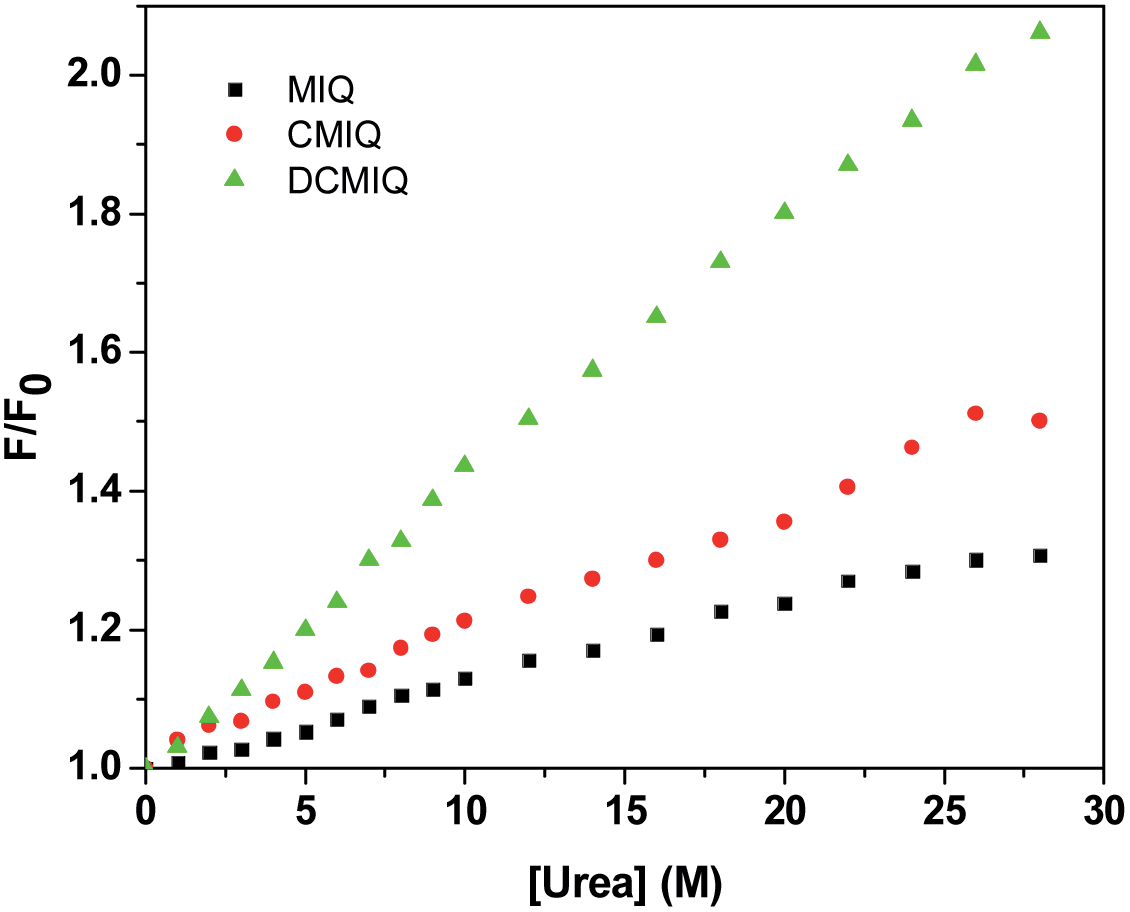 | ||
| Fig. 4 Relative emission plots for DCMIQ, CMIQ and MIQ bound to ct-DNA on application of urea. | ||
Circular dichroism spectral studies
CD spectroscopic technique is extensively used as a powerful tool in elucidating the modifications of the secondary structure of biopolymers as a result of interaction with small molecules.34–38 Perturbation in the secondary structure of ctDNA upon interaction with MIQ and its chloro derivatives was studied by observing the changes in CD signals. Fig. 5 shows the intrinsic CD spectra of ctDNA in aqueous buffer at various concentrations of the externally added probe.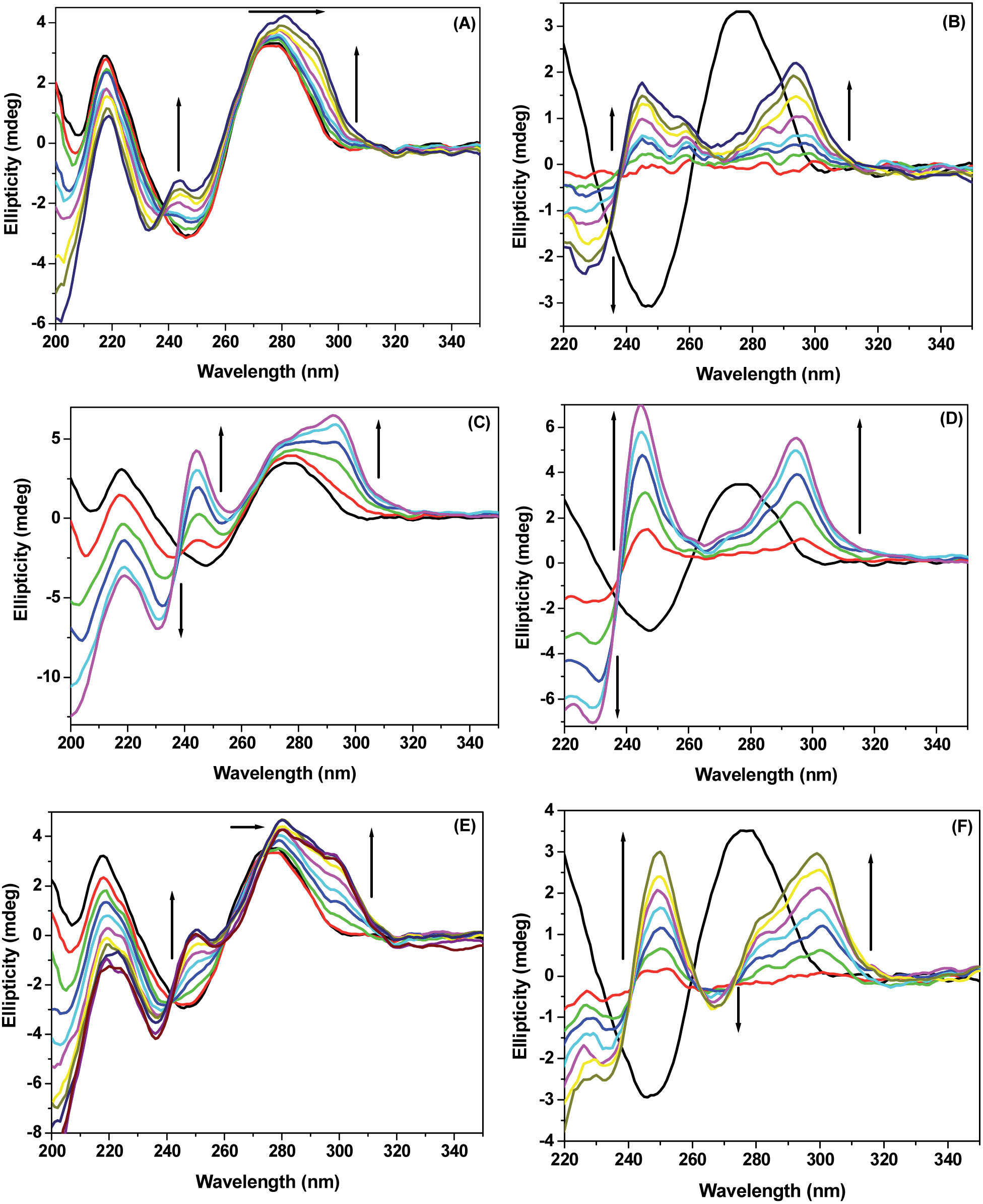 | ||
| Fig. 5 CD spectra of ctDNA with varying concentrations of (A) MIQ (up to 32 μM), (C) CMIQ (up to 40 μM), and (E) DCMIQ (up to 32 μM). (B), (D) and (F) are corresponding difference CD spectra of ctDNA in presence of MIQ, CMIQ and DCMIQ, respectively. The original CD spectrum is shown in black. 62 μM ctDNA has been used in each case. | ||
The intrinsic CD profile of DNA in the far-UV wavelength regime (220–320 nm) is characterized by a positive peak at ∼276 nm and a negative peak at ∼247 nm. These are signature bands for the right-handed B-form with 10.4 base pairs per turn, which is the normal form of ctDNA in aqueous buffer.39,8 Remarkable change in the CD signal of ctDNA is observed with increase in concentration of MIQ, CMIQ and DCMIQ. All the three molecules are planar and achiral in nature. Thus, presumably the modifications observed in the CD spectra due to addition of the three indoloquinolines are solely because of structural changes in ctDNA. On addition of MIQ, the 276 nm peak shifts to 280 nm with concomitant emergence of two positive induced CD (ICD) signals at 245 nm and 293 nm. The formation of these ICD peaks can be more clearly observed in the corresponding difference CD spectra. Similar is the trend for CMIQ and DCMIQ but in these cases the positive ICD peaks are more prominent and the shift of 276 nm peak is somewhat unclear compared to the previous case. Interestingly in the difference spectrum of DCMIQ we observed a negative ICD peak at ∼267 nm. The intensity of the positive ICD peaks follows the order CMIQ > DCMIQ > MIQ. The positive ICD signals for MIQ and CMIQ indicate that the principal axes of ctDNA binding site and the ligand, i.e., the dyad and helix axes of ctDNA are parallel. Hence, the molecules are intercalated in a parallel fashion to the ctDNA base pairs.40,41 The result is similar in case of DCMIQ except the appearance of the negative signal. This bi-signate signal in this case may be attributed to the intercalation mode of binding with the zwitterionic dimer of DCMIQ. Formation of zwitterion and zwitterionic dimer of DCMIQ in ground state is already established in one of our previous studies.23 Though intercalation with dimer is rare but such examples are known.42,43
Selectivity in probe–DNA binding
It is extremely important to know about the selectivity of binding of guests with DNA. In context to our present study, we chose two small sequenced DNA. One of them has the –GCTCG– sequence (primer 2 or P2) and the other one has the –AATT– sequence (primer 1 or P1). CD spectral changes of the oligos were analyzed to find the selective mode of binding of the three probe molecules (Fig. 6). Gradual addition of MIQ to P2 shows a decrease in ellipticity and the peak at ∼290 nm gets blue shifted by ∼5 nm whereas, in case of P1, no appreciable change was observed with MIQ. On addition of CMIQ, however, ellipticity of both the oligos gets reduced. In case of DCMIQ there was no change in P2 signal but intensity of the 290 nm band of P1 progressively decreased with a corresponding 5 nm hypsochromic shift.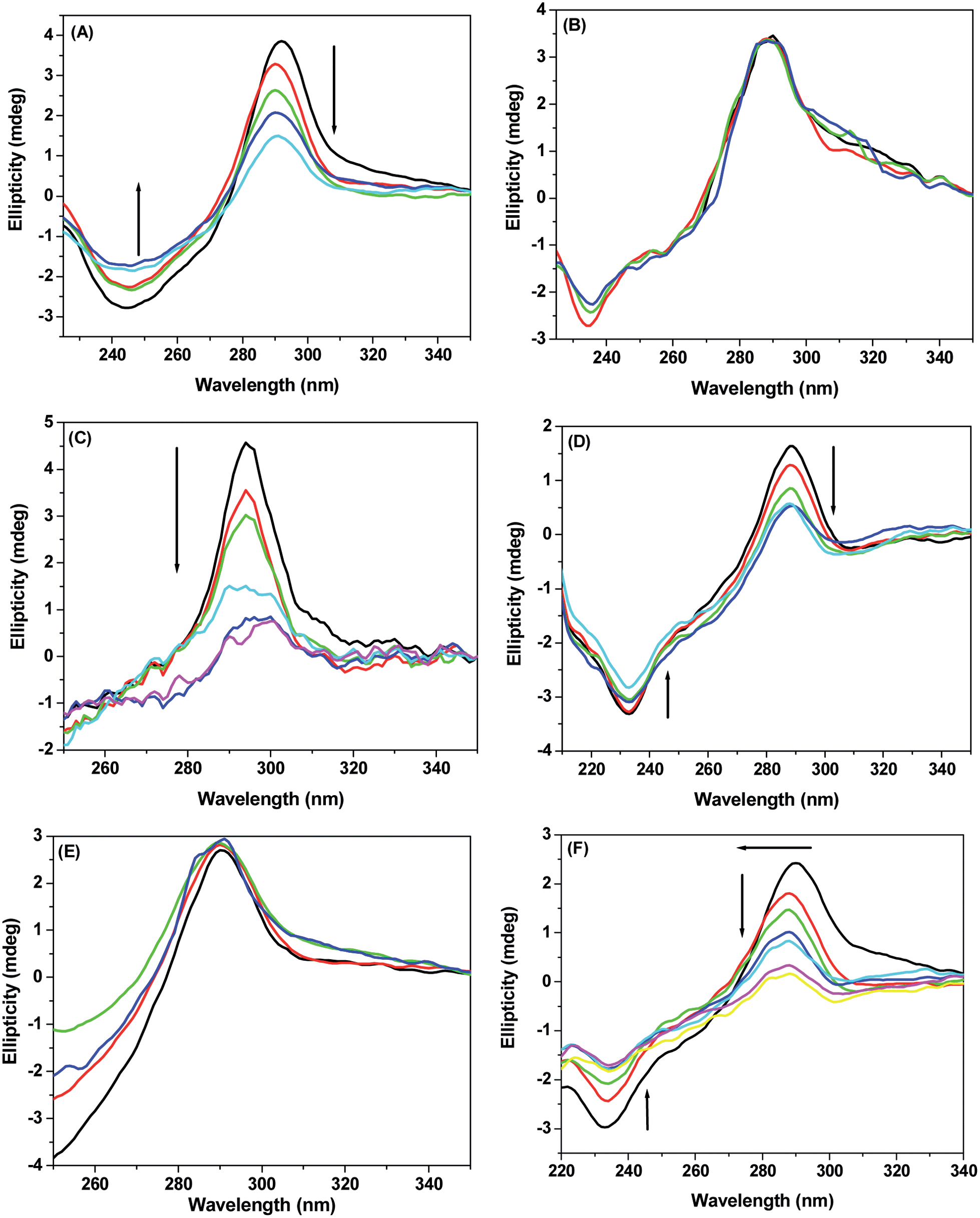 | ||
| Fig. 6 CD spectra for interaction of chosen oligos with MIQ, CMIQ and DCMIQ. (A), (C) and (E) demonstrate the changes due to interaction of the drugs with P2 and (B), (D) and (F) show the results due interaction of the probes P1. Concentration of the oligos used is 6 μM. | ||
The results dictate about the selectivity of the three compounds to bind to the specific primers. We observed that MIQ selects P2, CMIQ has no selectivity, and DCMIQ selects P1. Further, indications are obtained that the selectivity shifts from –AATT– to –GCTCG– sequences for DCMIQ and CMIQ to parent MIQ. It has been mentioned earlier that on addition of chloro functionality to the methylindoloquinolines, the molecule becomes more hydrophobic that increases the tendency toward binding to the –GCTCG– sequence. Moreover, since the chloro group/s can form weak hydrogen bonds, consequently when chloro derivatives interact with P2, stronger binding is expected to occur. Although there is no clear explanation about the interaction of CMIQ with both the oligos, but the asymmetric shape of the molecule may be a reason for such a behavior. Reduction of ellipticity of oligos is a well known signature for intercalation of drugs with DNA.
Melting of DNA helix on interaction with MIQ, CMIQ and DCMIQ
With rise in temperature hydrogen bonds between the DNA base pairs start to cleave resulting into random coil. The melting temperature (Tm) of DNA is defined as the temperature at which half of the DNA strands are in the double-helical state and half in the random-coil state. Tm depends on the length, the specific nucleotide sequence, and amount of GC base pair present in DNA. Intercalation of small molecules into the DNA double helix is known to perturb the thermal stability of the latter, which gets manifested through alteration in Tm. However, electrostatic or groove-binding interactions of small molecules with DNA lead to only insignificant modulation in Tm. This is easily understandable, since melting of the DNA helix requires rupture of the hydrogen bonds between the base pairs, but electrostatic or groove-binding of a dye to DNA can hardly affect that. It is only the intercalative mode of binding which can perturb the hydrogen bonding interaction between the DNA base pairs and thus Tm gets remarkably modified. Therefore, any significant perturbation in the Tm of the DNA helix in presence of the guest molecule is an important evidence for the intercalative mode of binding.44,45In the present study, we have monitored the change in Tm of free ctDNA and upon interaction with MIQ, CMIQ and DCMIQ to get an idea about the effects on the DNA due to binding to the different guests. Tm of pure ctDNA is 76 °C that matches well with previous reports.46 The ctDNA sample that we used consists of 160 base pairs on an average. Fig. 7 clearly reflects the ability of all the three methylindoloqinoline derivatives to enhance the thermal stability of DNA. With addition of MIQ, Tm of ctDNA shifts to 80.5 °C and addition of CMIQ and DCMIQ leads to an enhancement of the Tm to 85.5 °C and 89 °C, respectively. The results reinforce the intercalative mode of binding of all the three molecules to DNA and show that chloro functionality/functionalities make/s the parent indoloquinolines more effective DNA intercalator as reflected by the changes in the Tm of ctDNA.
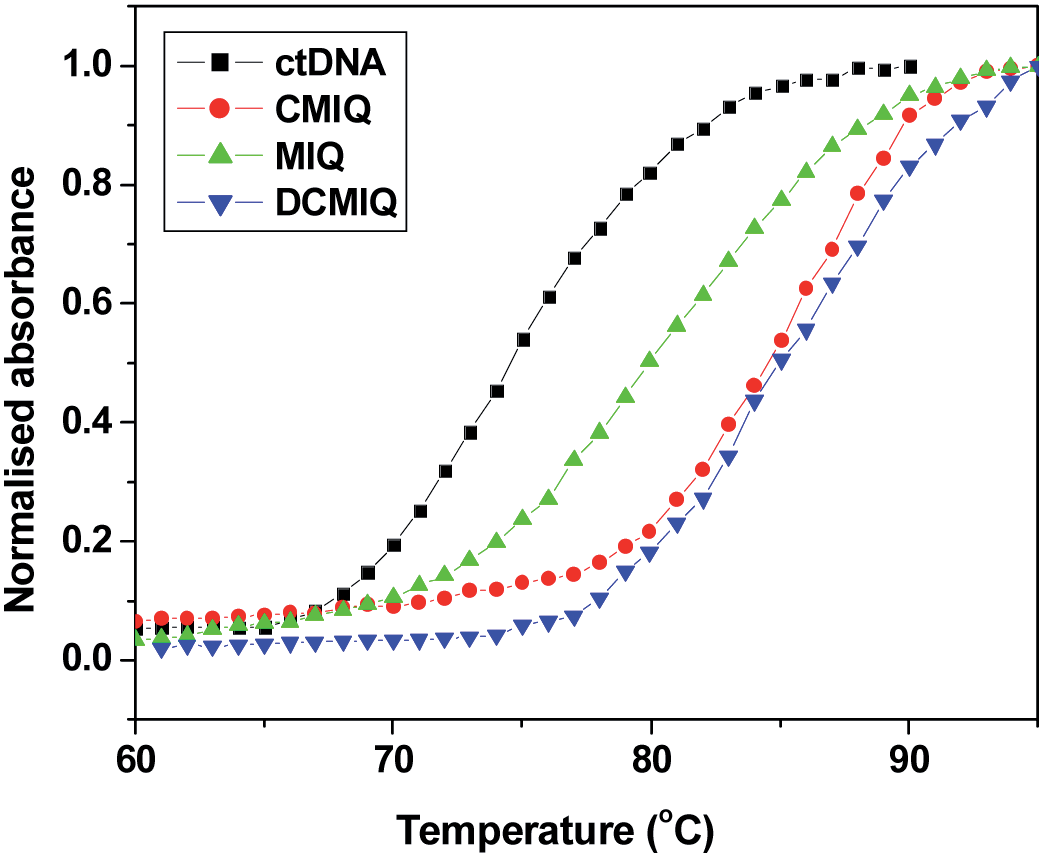 | ||
| Fig. 7 Thermal melting profiles of ctDNA, MIQ–DNA, CMIQ–DNA and DCMIQ–DNA composites as constructed by observing the relative absorbance at λabs = 260 nm as a function of temperature. | ||
Conclusions
Interaction of three indoloquinoline compounds, MIQ, CMIQ and DCMIQ, with model DNA has been studied. The findings established that these biologically potent drugs preferably intercalate to DNA. The chloro derivatives of the parent compound are better binders to DNA helix than the normal molecule. The guest–host interaction could be monitored well by observing the remarkable changes in fluorescence signals from the drug molecules themselves. The after effect of the interaction to DNA has been studied by using circular dichroism spectroscopy where we could monitor the changes occurred in the DNA double helix. The chloro derivatives of the indoloquinolines helped in enhancing the melting temperature of DNA due to intercalation. GC and AT rich DNA oligos were used to understand the selectivity of the indoloquinoline derivatives towards intercalation in the DNA double helix. These findings will help in developing methylindoloquinoline based drugs for therapeutic control over the action on cellular DNA.Experimental
Materials
MIQ, CMIQ, and DCMIQ were synthesized, purified, and analyzed in the organic chemistry laboratories of Birla Institute of Technology and Science, Pilani following procedures mentioned elsewhere.47 Stock solutions of the compounds (1.001 × 10−3 M) were prepared in pure methanol. ctDNA, oligomers, Tris–buffer tablets, urea and potassium iodide were bought from Sigma-Aldrich and used as received. All the spectroscopic grade solvents were procured from Spectrochem, India. Triple distilled water has been used throughout the experiment. Stock solutions were prepared by dissolving solid DNA in Tris–buffer (pH = 7.6) and stored at 4 °C whenever necessary. Freshly prepared DNA solutions were used for the experiments. The purity of DNA was confirmed by monitoring the ratio of absorbance at 260 nm to that at 280 nm, which was in the range 1.8–1.9. The concentration of DNA was determined spectrophotometrically, using εDNA = 13600 M−1 cm−1 at 258 nm.48 In case of the primers, solutions were prepared following supplied protocol.Methods
The absorption spectra were recorded using a Varian Cary 300 Bio UV-vis spectrophotometer equipped with DNA melting attachment. Fluorescence measurements were performed using a PerkinElmer LS 55 scanning spectrofluorimeter. The steady state anisotropy, r, can be represented as r = (IVV + GIVH)/(IVV + 2GIVH), where IVH and IVV are the intensities obtained from the excitation polarizer oriented vertically and the emission polarizer oriented in horizontal and vertical positions, respectively. The factor G is defined as G = (IHV)/(IHH). Circular dichroism (CD) spectra were recorded on a JASCO J-720 spectrometer, using a rectangular quartz cuvette of path length 1 cm. Spectra shown are average of three successive scans recorded at a scan speed of 50 nm min−1. Appropriate blank spectra were recorded using Tris–buffer (0.1 N) and subtracted from the spectra of the compounds. All the experiments were performed at ambient temperature (298 K).Acknowledgements
Financial support from DST (SR/S1/PC-35/2011) is gratefully acknowledged. P.G. acknowledges CSIR, New Delhi for his fellowship. We express our gratitude to Professor Dalip Kumar and his group from BITS, Pilani for providing the indoloquinoline derivatives used in the present work.Notes and references
- B. Alberts, A. Johnson, J. Lewis, R. Martin, K. Roberts and P. Walter, Molecular Biology of the Cell, Garland Science, New York, 4th edn, 2002 Search PubMed.
- S. K. Zaidi, D. W. Young, J. Y. Choi, J. Pratap, A. Javed, M. Montecino, J. L. Stein, J. B. Lian, A. J. van Wijnen and G. S. Stein, J. Biol. Chem., 2004, 279, 43363–43366 CrossRef CAS PubMed.
- P. B. Dervan, Bioorg. Med. Chem., 2001, 9, 2215–2235 CrossRef CAS.
- D. Fink, S. Nebel, S. Aebi, H. Zheng, B. Cenni, A. Nehmé, R. D. Christen and S. B. Howell, Cancer Res., 1996, 56, 4881–4886 CAS.
- K. I. E. McLuckie, Z. A. E. Waller, D. A. Sanders, D. Alves, R. Rodriguez, J. Dash, G. J. McKenzie, A. R. Venkitaraman and S. Balasubramanian, J. Am. Chem. Soc., 2011, 133, 2658–2663 CrossRef CAS PubMed.
- V. Heiser, E. Scherzinger, A. Boeddrich, E. Nordhoff, R. Lurz, N. Schugardt, H. Lehrach and E. E. Wanker, Proc. Natl. Acad. Sci. U. S. A., 2000, 97, 6739–6744 CrossRef CAS PubMed.
- J. K. Barton, Science, 1986, 233, 727–734 CrossRef CAS.
- D. Sarkar, P. Das, S. Basak and N. Chattopadhyay, J. Phys. Chem. B, 2008, 112, 9243–9249 CrossRef CAS PubMed.
- B. K. Paul and N. Guchhait, J. Phys. Chem. B, 2011, 115, 11938–11949 CrossRef CAS PubMed.
- M. J. Carvlin and R. J. Fiel, Nucleic Acids Res., 1998, 26, 3724–3728 CrossRef PubMed.
- S. Z. Bathaie, A. Bolhasani, R. Hoshyar, B. Ranjbar, F. Sabouni and A. A. Moosavi-Movahedi, DNA Cell Biol., 2007, 26, 533–540 CrossRef CAS PubMed.
- E. Tuite and B. Norden, J. Am. Chem. Soc., 1994, 116, 7548–7556 CrossRef CAS.
- M. Munde, M. A. Ismail, R. Arafa, P. Peixoto, C. J. Collar, Y. Liu, L. Hu, M.-H. David-Cordonnier, A. Lansiaux, C. Bailly, D. W. Boykin and W. D. Wilson, J. Am. Chem. Soc., 2007, 129, 13732–13743 CrossRef CAS PubMed.
- J.-L. Pousset, M.-T. Martin, A. Jossang and B. Bodo, Phytochemistry, 1995, 39, 735–736 CrossRef CAS.
- P. M. Fresneda, P. Molina and S. Delgado, Tetrahedron Lett., 1999, 40, 7275–7278 CrossRef CAS.
- A. Molina, J. J. Vaquero, J. L. Garcia-Navio, J. Alvarez-Builla, B. de Pascual-Teresa, F. Gago, M. M. Rodrigo and M. Ballesteros, J. Org. Chem., 1996, 61, 5587–5599 CrossRef CAS.
- P. Grellier, L. Ramiaramanana, V. Millerioux, E. Deharo, J. Schrevel, F. Frappier, F. Trigalo, B. Bodo and J. L. Pousset, Phytother. Res., 1996, 10, 317–321 CrossRef CAS.
- S. Y. Ablordeppey, P. Fan, S. Li, A. M. Clark and C. D. Hufford, Bioorg. Med. Chem., 2002, 10, 1337–1346 CrossRef CAS.
- G. S. Sundaram, C. Venkatesh, U. K. S. Kumar, H. Ila and H. Junjappa, J. Org. Chem., 2004, 69, 5760–5762 CrossRef CAS PubMed.
- W. D. Wilson and R. Jones, Intercalation Chemistry, ed. M. S. Whittinham and A. J. Jacobson, Academic Press, New York, 1981 Search PubMed.
- S. Van Miert, S. Hostyn, B. U. W. Maes, K. Cimanga, R. Brun, M. Kaiser, P. Matyus, R. Dommisse, G. Lemiere, A. Vlietinck and L. Pieters, J. Nat. Prod., 2005, 68, 674–677 CrossRef CAS PubMed.
- P. Ghosh, S. S. Jaffer, T. Das, A. Maity, M. N. Kumar, D. Kumar and P. Purkayastha, J. Lumin., 2011, 131, 147–154 CrossRef CAS PubMed.
- P. Ghosh, S. S. Jaffer and P. Purkayastha, J. Phys. Chem. B, 2011, 115, 2046–2054 CrossRef CAS PubMed.
- M. Oda and N. Sato, Chem. Phys. Lett., 1997, 275, 40–45 CrossRef CAS.
- N. K. Modukuru, K. J. Snow, B. S. Perrin, J. Thota and C. V. Kumar, J. Phys. Chem. B, 2005, 109, 11810–11818 CrossRef CAS PubMed.
- G. Zhang, Y. Pang, S. Shuang, C. Dong, M. M. F. Choi and D. Liu, J. Photochem. Photobiol., A, 2005, 169, 153–158 CrossRef CAS PubMed.
- J. R. Lackowicz, Principles of Fluorescence Spectroscopy, Springer, New York, 3rd edn, 2006 Search PubMed.
- A. Maity, S. S. Jaffer, T. Das, P. Ghosh and P. Purkayastha, Langmuir, 2011, 27, 4068–4075 CrossRef CAS PubMed.
- P. Ghosh, A. Maity, T. Das, J. Dash and P. Purkayastha, J. Phys. Chem. C, 2011, 115, 20970–20977 CAS.
- D. Dey, N. R. Pramanik and S. Basu, J. Phys. Chem. B, 2009, 113, 8689–8694 CrossRef CAS PubMed.
- S. Sen, D. Sukul, P. Dutta and K. Bhattacharyya, J. Phys. Chem. B, 2002, 106, 3763–3769 CrossRef CAS.
- H. Baruah and U. Bierbach, JBIC, J. Biol. Inorg. Chem., 2004, 9, 335–344 CrossRef CAS PubMed.
- W. Vanderlinden, M. Blunt, C. C. David, C. Moucheron, A. K. D. Mesmaeker and S. De Feyter, J. Am. Chem. Soc., 2012, 134, 10214–10221 CrossRef CAS PubMed.
- W. A. Baase and W. C. Johnson Jr, Nucleic Acids Res., 1979, 6, 797–815 CrossRef CAS PubMed.
- A. Rodger and B. Nordén, Circular Dichroism and Linear Dichroism, Oxford University Press, New York, 1997, ch. 3 Search PubMed.
- K. Harata, Bioorg. Chem., 1981, 10, 255–265 CrossRef CAS.
- W. Curtis Johnson Jr, Proteins: Struct., Funct., Genet., 1990, 7, 205–214 CrossRef PubMed.
- A. Bonincontro, M. Falivene, C. LaMesa, G. Risuleo and M. Ruiz Pena, Langmuir, 2008, 24, 1973–1978 CrossRef CAS PubMed.
- N. K. Modukuru, K. J. Snow, B. S. Perrin and C. V. Kumar, J. Phys. Chem. B, 2005, 109, 11810–11818 CrossRef CAS PubMed.
- W. Zhonga, J.-S. Yu, Y. Liang, K. Fan and L. Lai, Spectrochim. Acta, Part A, 2004, 60, 2985–2992 CrossRef PubMed.
- N. C. Garbett, P. A. Ragazzon and J. B. Chaires, Nat. Protoc., 2007, 2, 3166–3172 CrossRef CAS PubMed.
- H.-K. Kim, J.-M. Kim, S. K. Kim, A. Rodger and B. Nordén, Biochemistry, 1996, 35, 1187–1194 CrossRef CAS PubMed.
- J. L. Mergny, G. Duval-Valentin, C. H. Nguyen, L. Perrouault, B. Faucon, M. Rougee, T. Montenay Garestier, E. Bisagni and C. Helene, Science, 1992, 256, 1681–1684 CAS.
- M. T. Bjorndal and D. K. Fygenson, Biopolymers, 2002, 65, 40–44 CrossRef CAS PubMed.
- K. Sandström, S. Wärmländer, J. Bergman, R. Engqvist, M. Leijon and A. Gräslund, J. Mol. Recognit., 2004, 17, 277–285 CrossRef PubMed.
- J. G. Duguid, V. A. Bloomfield, J. M. Benevides and G. J. Thomas Jr, Biophys. J., 1996, 71, 3350–3360 CrossRef CAS.
- D. Kumar, M. N. Kumar and V. S. Rao, Chem. Lett., 2009, 38, 156–157 CrossRef CAS.
- J. L. Jhonson, Methods Microbiol., ed. G. Gottschalk, Academic Press Inc., 1985, vol. 18, pp. 1–74, ch. 2 Search PubMed.
| This journal is © The Royal Society of Chemistry 2014 |