The RNA superhighway: axonal RNA trafficking of kappa opioid receptor mRNA for neurite growth
Li-Na
Wei
Department of Pharmacology, University of Minnesota Medical School, 6-120 Jackson Hall, 321 Church St. SE, Minneapolis, MN 55455, USA. E-mail: weixx009@umn.edu; Fax: 612-6258408; Tel: 612-6259402
First published on 29th November 2010
Abstract
Neurons are highly polarized cells with extensive sub-cellular compartmentalization to accommodate diverse local needs. Information flows between the pre-synaptic (axon) and post-synaptic (dendrite) compartments, as well as between the soma and the nerve termini. It is critical that a neuron controls efficient molecular transfer/transport through its axon. But this is extremely challenging to study because of the long distance molecules must travel through axons and the apparent contextual difference in the axons' various local environments, which should not be examined in isolation. Understanding the action in neurons of drug-responsive neurotransmitter receptors such as opioid receptors has been hindered by the lack of information on the control of molecular flow between such various sub-cellular neuron compartments. Recent studies have uncovered new transport systems other than the classical vesicle transport in neurons, particularly those utilizing various granules containing certain RNAs including protein-coding mRNAs. Through integrated approaches exploiting various experimental systems, tools, and methodologies, studies have provided solid evidence for functional roles of specific RNA granules in several biological processes crucial for the survival and function of neurons. These include neurons' transport of molecules/information, stress response, and local axonal translation. By using the kappa opioid receptor (KOR) as a model, studies have also revealed a novel physiological function of KOR in mediating growth factor-stimulated neurite outgrowth during a critical period of development, which requires specific KOR mRNA untranslated sequences that direct spatially and temporally specific expression of KOR.
 Li-Na Wei | Li-Na Wei is a Distinguished McKnight University Professor at the University of Minnesota, Minneapolis, and a Professor of the Department of Pharmacology in the Medical School. Research interests in her lab focus on transcriptional and post-transcriptional regulation of protein expression in embryonic stem cells and in cellular differentiation processes of neurons and adipocytes. The ultimate goal is to understand the integration of various biological processes occurring in different sub-cellular compartments through the action of specific regulators and signaling pathways in the context of functional cells and in whole animals. |
Insight, innovation, integrationIntegration of various methodologies, including that of molecular, biochemical, cellular, genetic, imaging, and neurobiological, into a biological system that has a physiological relevance enables gaining new insights, mechanisms and biological functions of molecules in neurons that have a unique biological context. The review summarizes sequential and step-wise discoveries that used various innovative methodologies to address challenging issues in neurons that critically depend upon temporal and spatial specificity and coordination among various subcellular domains. Discussion is also provided regarding future difficulties to be addressed in order to conceptualize a functional neuronal circuitry in the whole animals, through the integration of compartmentalized subcellular events and dissected molecular details. |
Introduction
Neurons are highly specialized cells that rely on cellular polarization and sub-cellular compartmentalization to accommodate diverse needs for survival and functions. Polarization of a neuron is characterized by its axon-dendrite polarity, critical to the integration and transmission of information in the brain.1 With the establishment of this polarity early during neuronal differentiation, the unique “pre-synaptic” (axonal) and “post-synaptic” (dendritic) compartments/domains begin to form and eventually constitute most of the extra-nuclear domains in a neuron. These pre- and post-synaptic compartments grow out of its “soma”, or the cell body where the nucleus resides, and in a mature neuron, axon can elongate to thousands of times the diameter of the soma. Inside a neuron, information flows, principally, from the soma to the axon (Fig. 1). This polarity, as well as the unique features of axonal and dendritic compartments, is specified by the actions of both stochastic and asymmetric activation of intracellular signaling pathways that may be stimulated by extracellular cues. Concomitantly, growth of axons and dendrites occurs primarily through microtubule assembly and stabilization, which also provides a physiological signal to specify neuron cell polarization.2 In addition, the assembly and establishment of “synapses” on the termini of growing axons (the pre-synaptic compartment) contacting dendrites of another neuron at the receiving end (the post-synaptic compartment) eventually culminates and defines a specific circuitry through which biological/chemical signals and electric information can then be carried and transmitted.3 In general, between two neurons, information flows from the pre-synaptic compartment (axon) of a neuron to the post-synaptic compartment (dendrite) of the other neuron receiving this information input. | ||
| Fig. 1 Information flows and molecular transport from soma to axon terminal inside a neuron. EGF priming in the soma initiates nuclear export (a) of the mature KOR mRNA granule, which is used to produce KOR proteins (brown) to be deposited in the soma or dendrites (b), or to be transported into axons (yellow arrow). Axonal transport of KOR mRNA can be stimulated by neuron activity (depolarization). At the axon terminal, axon guidance cues activate FAK, which subsequently triggers a cascade of translational regulation of KOR mRNA arriving at the axon terminal to produce KOR that is then deposited in the axon terminal (pink). | ||
To ensure a smooth and efficient information flow inside a neuron, the entire axon must serve as an information superhighway, both during its growth/differentiation phase and after its maturation. Because of a neuron's compartmental features, its enormous area and distance that biological molecules must traverse during their transport, and the intimate relationship between its soma and the axon, it has been difficult to study how information is carried by various molecules, and how it flows through different compartments in a neuron. In particular, it remains extremely challenging to delineate how the soma executes its control over distant events occurring in axon termini, and how the axon termini react and give feedback to input from the soma. To address these questions, it is essential to apply highly integrated models, tools, and methodologies in a physiologically relevant experimental system that can be uniquely complex.
The axon, as an information superhighway, must manage its traffic efficiently not only to meet its spatial demands, but also to accommodate highly rigorous demands for temporal specificity. The demand for speedy transport in a neuron is enormous. To meet this need, neurons employ different kinds of transport systems, primarily the fast and slow transport, which utilize vesicle cargos.4 Classical studies have established that synaptic proteins are mainly carried by vesicles derived from the trans-Golgi network.5,6 However, while vesicles can carry a large load of different kinds of molecules needed for growth and function of axons, they are not sufficient for the axon to react in timely fashion to local cues and assemble components needed for specific local events. Because of the long distance vesicles have to travel, they are not likely to be able to replenish all the components needed at a distance in a short period of time. Therefore, certain molecules may have to be produced locally at the time of need, which would require de novo local production, i.e.protein translation. In this regard, recent studies have documented transport systems other than those viavesicles, such as transport by various RNAs.7,8 Further, local translational activation of specifically localized mRNAs has also been widely observed in various subcellular compartments including axons.8,9 Hence, it appears that RNA granules may be highly relevant to axonal transport and local translation as depicted in Fig. 1, and can be critical to neurons' survival such as at the time of stress and need for their function when stimulated. To understand how these events may be coupled and coordinately regulated in a physiological context also requires integration of different tools and methodologies applied in a relevant experimental system.
Classical pharmacology examines drug receptors including opioid receptors (ORs) to understand their pharmacological actions and mechanisms of actions, which are often quite different from their actions in a physiological context. For ORs, the principal pharmacological issues concern the efficacy of opioids as analgesics and their side effects such as the induction of tolerance and drug addiction.10,11 These are mostly neuron-specific problems. Recent molecular and genetic studies of the opioid peptides and OR reveal certain previously ignored physiological actions of the endogenous opioid peptides and ORs, which could also underlie their pharmacological actions and contribute to their pharmacological problems in the nervous system.12 But the molecular mechanisms underlying these physiological activities are entirely unclear, and require re-examination of opioid peptides and ORs in more physiologically relevant experimental systems where neurons can be dissected and monitored in real time. Among the three ORs (mu, delta and kappa), kappa OR (KOR) is best studied in terms of its regulatory events in neurons;8,13 therefore this review will use KOR as an example to illustrate how integrated studies, especially in the context of primary neurons, have helped discover new mechanisms and functions otherwise impossible to obtain.
KOR ligands have not been considered for primary analgesics because they are not as potent as those targeting mu OR; however, KOR agonists in fact provide better relief of inflammatory, visceral, and neuropathic pain, and seem to have less abuse potential.14 Together with its endogenous ligands, the dynorphin peptides, the KOR/dynorphin system is also known to modulate stress-induced and pro-addictive behaviors.15 KOR is detected in both pre- and post-synaptic compartments of neurons, and the molecular basis of its pharmacological and physiological actions involves numerous issues regarding its synthesis, regulation, trafficking and signal transduction. The difficulty in dissecting the molecular details is primarily due to the lack of tools to differentiate its pre- and post-synaptic actions in a physiological context. Another challenge is to understand how KOR differentiates itself from other OR members in mediating or modulating specific pain responses and/or animal behavior, because all the OR family members use very similar signal transduction pathways and are expressed in very similar brain areas.8 Classical pharmacological studies typically monitor and examine processes or responses in the context of whole tissue or cell preparations, which lacks the power to resolve the processes specific to certain sub-cellular compartments of a neuron. Most of these classical studies are likewise unable to address a specific time window when neurons receive inputs or are activated. To address these challenges in high-resolution studies of neurons, it is essential to employ innovative tools and methodologies that can dissect molecular details, specifically examining various sub-cellular compartments and detecting molecular events in real-time, all in a physiological context. Further integration of data gathered from these types of studies is critical to the understanding of physiological actions of drug receptors in neurons.
RNA granules connecting compartmentalized processes in neurons
1 RNA-mediated transport and spatially specific distribution of molecular complexes
One immediate task in determining the physiological activities of KOR is to learn its spatial specificity—whether the receptor protein is located in the neuron's pre- or post-synaptic compartment, and how it gets there. Initial clues that mRNA helps confer KOR spatial specificity came from the detection of multiple isoforms of its mRNA, all of these code for the same KOR protein.16 This observation prompted studies testing the hypothesis that the KOR mRNA isoforms encode specific information for the production, distribution, or regulation of KOR.16 In an animal behavior test, nerve injury triggering specific animal pain behavior was correlated with altered distribution of KOR mRNA isoforms in the injured spinal cord.17 Subsequently, using sandwich cultures where soma and fibers of neurons could be separately grown on different layers of the device, it was found that KOR mRNA isoforms were differentially distributed to the axons and the soma of dorsal root ganglia (DRG) neurons.18,19 This implies specific targeting or transport of different isoforms of KOR mRNA, as well as a relationship between their targeting or transport and certain animal behaviors or pain sensations. Ultimate proof of differential distribution and transport of KOR mRNA isoforms into axons was obtained by examining sub-cellular distribution of KOR mRNAs in primary DRG neurons using Campenot chambers, where soma and axons could be separately grown and monitored in two chambers on the same live culture.20 From a series of such Campenot chamber studies, it was concluded that KOR mRNAs containing intact 5′UTR and 3′UTR sequences are most effective for axonal transport. Further, KOR mRNA transport requires certain RNA-binding proteins such as HuR, and is mediated by microtubule- and kinesin-dependent transport processes that involve the COPI vesicle subunit Copβ1.20,21 But KOR mRNAs are not encapsulated in these vesicles. Importantly, mRNAs with defective 3′UTRs are ineffective for axonal transport in DRG neurons.21These experiments establish that neurons can engage certain RNA granules (those probably tethered to the COPI vesicles) for axonal transport (Fig. 2). These studies rely on integrated applications of various tools, including typical molecular and cellular methods, classical pharmacological agents, specific neurobiological systems, a sandwich culture method, the Campenot chamber compartment culture device, and a newly developed imaging tracking procedure allowing real-time monitoring. Through integrating these various approaches, it is possible to dissect the molecular details underlying the spatial specificity of KOR, which is mediated by granules of KOR mRNAs containing various untranslated RNA sequences.
 | ||
| Fig. 2 Signal transduction for axonal transport and translation of KOR mRNA. The mature KOR mRNA (curved blue shape)/HuR (yellow)/COP (light blue circle) granules are translationally silenced by HuR binding (with or without Grb7, red star), and transported (pink arrows) through the axon to its terminal where growth factors and axon guidance cues can activate FAK. During the transport, the granules may be sorted further if cells or axons encounter stress. When granules arrive the axon terminal, it may be stored and kept silenced, or activated by local cues such as netrin-1 or growth factors such as EGF. These local cues activate FAK, and the activated FAK initiates a cascade of protein phosphorylation, triggering the release of pGrb7 (red cross shape) from KOR mRNA to produce KOR (pink triangle). These locally produced KOR molecules are then deposited to the axon terminal at the time of need. It remains unclear whether the KOR mRNA granule during transport is bound by Grb7, and whether the translatable KOR mRNA is bound by HuR (yellow). | ||
2 RNA-mediated silencing and neuronal stress
The experiments described above conclude that KOR mRNAs can be transported from the soma to nerve fibers such as axons by tethering to COPI vesicles through the typical microtubule- and kinesin-dependent axonal fast transport machinery. The substantiation that this type of RNA transport occurs in primary neurons makes it important to address whether, how, and where these KOR mRNAs are translated. This was addressed in part through parallel studies, which first revealed a specific KOR mRNA-silencing mechanism that utilized a specific KOR RNA binding protein, Grb7.22 Secondly, this mechanism is coupled to a nuclear export process that relies on the exportin CRM tethered to another KOR RNA binding protein, HuR, which also interacts with Grb7.23 Thus, the silenced KOR mRNA can be exported out to the cytoplasm from the nucleus by forming a complex containing Grb7 binding at its 5′-UTR, HuR binding to multiple sites along the KOR mRNA, and the export machinery, CRM (Fig. 3). This provides molecular details that connect nuclear and extra-nuclear neuronal compartments in mRNA processing, transport and regulation.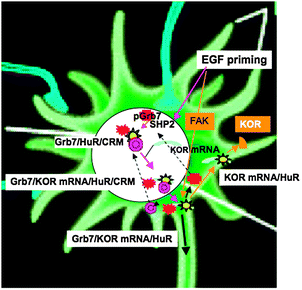 | ||
| Fig. 3 Signal transduction initiating nuclear export and silencing of KOR mRNA in the soma. EGF primes the soma to initiate, in the nucleus (large white circle), the export (pink arrows) of mature KOR mRNA (light blue curve shape) that is bound by hypo-phosphorylated Grb7 (red stars) and HuR (yellow star), and carried by exportin (CRM complex, pink circle). The KOR RNA granule to be exported from the nucleus contains, at a minimum, mature KOR mRNA, Grb7, HuR and CRM, and is translationally silenced. In the cytoplasm, the exported KOR mRNA granules can be activated by FAK (brown square) which phosphorylates Grb7 that then leaves KOR mRNA, freeing it for translation to produce KOR (brown triangle) that can be deposited in the soma surface or dendrites, or transported into the axon through classical vesicle trafficking (heavy black arrow). FAK activation in the soma may be triggered by growth factors like EGF. The phosphorylated Grb7 (red crosses) and free exportin components (pink circles) in the cytoplasm may recycle (dashed arrows) back into the nucleus. It is unclear whether cytosolic HuR (yellow) can recycle into the nucleus. It is also unclear if Grb7 binding is required for axonal transport of KOR RNA granules after they migrate out to the axon. | ||
In delineating these sequential events, several experimental systems and methodologies have proven critical. First it was the important step to identify the critical KOR RNA-binding protein, Grb7, in a yeast three-hybrid screening system.22 This protein binds to the 5′ terminal region of mature KOR mRNA, which adopts a specific stem loop structure in the 5′-UTR. Grb7, in its hypo-phosphorylated form, suppresses KOR translation by binding to a stem loop of KOR mRNA and blocking its 5′ Cap for translation factors' access. Second, among the other constituents in the complex identified using molecular and cellular methodologies, one, HuR, proved to be another key KOR mRNA-binding protein that directly interacts with Grb7.23 This expands the scope of molecular studies where functional molecular complexes can be systemically and sequentially assembled. Third, by using functional assays with the approach of real-time imaging of photo-activatable GFP fusion proteins, it is possible to establish that tethering of the nuclear export machinery CRM, viaHuR, to the Grb7/KOR mRNA complex is responsible for the nuclear export of KOR mRNA. As depicted in Fig. 3, mature KOR mRNA particles are formed in the nucleus as, at a minimum, Grb7/KORmRNA/HuR tri-molecular complexes that can be exported by tethering to the CRM export machinery, and are translationally silenced by tight Grb7 binding. Binding of KOR mRNA by these two proteins is initiated in the nucleus, probably during or immediately after KOR mRNA maturation.
Once KOR mRNA reaches the cytoplasm, its fate then depends upon the local environment. It may be locally activated if appropriate stimulatory factors are present (see next section 3), or it may be stored in the cytoplasm or further transported to nerve fibers including axons. Most cells, including neurons, can form various granules, cytoplasmic structures where specific proteins and RNAs (together as heterogeneous mRNPs) can be found. Two major granules that are relevant to the dynamics of RNAs, including mRNAs, are P-bodies (PBs) and Stress Granules (SGs). PBs are ubiquitously present and contain various mRNP complexes as well as, generally, the RNA decay machinery. They are believed to serve the function of RNA sorting and degradation.24 SGs are formed when cells encounter stress and arrest their translation initiation, and typically contain poly(A) + mRNA, 40S ribosomal subunits, various translation initiation factors and poly(A)-binding protein, etc.25 Therefore, it is believed that translation-arrested mRNAs that need to be protected for later activation are likely assembled into 48S pre-initiation complexes and stored in SGs for protection. SGs can also contain other RNAs lacking the 40S ribosomal subunit,26 and sometimes certain mRNAs can shuttle between PBs and SGs. This is believed to represent a mechanism linking sorting and protection of various mRNAs in cells under a specific condition. KOR mRNA is found mostly in SGs when cells, including neurons, encounter stresses such as heat shock or arsenite treatment.27 Further, its 5′-RNA binding protein, Grb7, is also found in SGs and is required for the formation of KOR mRNA-containing SGs, under which condition KOR translation is shut down. Finally, when the stressful condition stops within a time window and cells recover, SGs are dissolved and KOR translation resumes.27 The formation and disassembly of SGs is an ATP-dependent process, requiring dynein motors;28 therefore, the dynamics of shuttling KOR mRNPs among granules (such as PBs and SGs), or between unstructured cytosol and cytoplasmic granular structures, would also depend upon dynein motors. Conceivably, once reaching the cytoplasm in the soma of a neuron, various KOR mRNA isoforms could be sorted into pools of silenced and stored, constitutively activated, or transported species, depending upon their RNA sequences, availability of binding proteins, and local environment. Constitutively active KOR mRNA species would produce KOR to be either inserted into the membrane surrounding the soma or transported in vesicles to be delivered to nerve termini. KOR mRNA species to be transported, or silenced and stored away, would be likely to serve special purposes, because they would be delivered to different parts of the neurons by kinesin or dynein motors, and, presumably, react to various extracellular cues and/or cell-autonomous local signals arising at different stages of neuron maturation or functional specification (see next section 3).
Studying the dynamics of KOR mRNA particles exported from the nucleus, i.e. whether they are stored or transported, also depends upon the assembly of various experimental systems and methodologies as described above into an experimental system where physiologically relevant questions can be addressed.
3 RNA-mediated local translation and temporal specificity
Translationally silenced KOR mRNAs, once released from either stored or transported granules, would be activated for translation when KOR proteins are needed. Studies to address what activates these mRNAs point to locally activated signaling pathways. The first clue about local activation of KOR translation came from the studies of KOR silencing by hypo-phosphorylated Grb7 (Fig. 3). Elucidating this KOR translational repression system has been critical to learning how KOR translational silencing is maintained and, conversely, activated. A series of biochemical studies first revealed that a switch of tyrosine phosphorylation on Grb7 was critical. Hypo-phosphorylation (on two specific tyrosine residues) maintains Grb7's RNA-binding ability; conversely, hyper-phosphorylation blunts its RNA binding. Thus an important task was to identify the trigger of Grb7 phoshorylation—which proved to be focal adhesion kinase (FAK).22,29 Upon phosphorylation by FAK, Grb7 is released from KOR mRNA, which can then be accessed by translation factors. Interestingly, FAK activity can be stimulated by various growth factors and cytokines, among which the most relevant signal to growing axons is the axon guidance cue, netrin-1.22 This has been validated in DRG neurons, wherein axonally provided netrin-1 elevates KOR protein level and its ligand binding receptor activity in axons that have also received exogenous KOR mRNA locally, i.e. these KOR mRNAs are not derived from the nucleus.21,29 Another stimulus relevant to axonal behavior is local depolarization, or neuronal activity. In DRG neurons cultured in Campenot devices, potassium chloride-induced depolarization in the axonal compartment rapidly elevates translation of axon-distributed KOR mRNA—again, with this event requiring no input from the soma.20 Finally, similar to netrin-1, epidermal growth factor (EGF) is also able to activate FAK locally, thereby stimulating KOR mRNA translation.23Together, these experiments demonstrate highly coordinated and sequential processes using mRNA granules for fine control to produce locally needed proteins, with production coordinated by various extracellular signals/factors/cues. These signals regulate the behavior of certain constituents of KOR mRNA granules—primarily their RNA-binding proteins—thereby modulating the translatability of specific KOR mRNAs in a particular local context. Importantly, demonstrating the coupling of KOR mRNA export, transport, storage, and translation has depended upon integrating information gathered from studying each individual event. The molecular elements instrumental in this process include several RNA-binding proteins, specific RNA sequences, and particular signaling pathways required for nuclear export, storage/sorting, axonal transport and local translation. It is now clear that the different events occurring in various sub-cellular compartments of a neuron are coordinated to ensure the smooth flow of information from the soma to the axon, and that various sub-cellular compartments of a neuron can be regulated, autonomously, to execute local events in accordance with the specific contextual changes in the neuron. Through highly coordinated signal regulation, production of KOR proteins in a specific neuronal compartment can be effectively regulated. The response to local cues, even in a remote part of the axon, can be very efficient because it needs not to wait for commands from the soma; rather, each local event is intimately coordinated with specific upstream events. Coordination of these compartmentalized processes, as well as the ordered molecular events and signaling pathways, is collectively illustrated in Fig. 2 and 3.
Physiological action of KOR
Previously, KOR has been studied mostly using classical pharmacological approaches that address its pharmacological issues, such as its ligand binding specificity and its relationship with analgesia.10,11 More recent molecular and genetic studies, as well as parallel examination of its endogenous ligands (the dynorphin peptides) revealed a relationship between the dynorphin-KOR system and animal behavior.12 Given that KOR mRNA is expressed during early embryogenesis30 long before neuronal maturation, it is tempting to speculate that a physiological action beyond pain sensation and analgesia exists for the dynorphin-KOR system.Using the KOR gene knockout animals, it is possible to obtain primary tissues/cells that are entirely defective in the KOR system. Such tissues/cells can be used to study specific molecular and cellular defects that might reveal novel physiological actions of KOR. We first hypothesized that KOR may be involved in certain cellular processes during early developmental stages such as neurogenesis, neuronal differentiation, or neuronal growth/maturation. Because KOR-depleted animals appear grossly normal and are able to function in most aspects of their life,31 it is unlikely that KOR plays a critical role in neurogenesis or neuronal differentiation because these processes would likely impact the formation of normal brain circuitry and mutant animals would be extremely dysfunctional. Such dysfunction has not been seen in the KOR mutant animals, which narrows down possible investigation to defects in neuronal growth/maturation. In mid-gestation, neuron growth requires various growth factors and cytokines, among which the most interesting and relevant is the EGF pathway. EGF is known to modulate neurite extension,32 and its signal is first seen by membrane EGF receptors, which could elicit cytoplasmic signaling pathways. EGF receptors can also be translocated to the nucleus to directly influence nuclear events.33 We found that EGF could facilitate axon outgrowth from normal DRG neurons collected from mid- to late-gestation stage embryos, and that its effects in KOR-depleted DRG neurons at the same developmental stages are drastically blunted.34 This observation suggested that KOR proteins are involved in mediating EGF-stimulated axon outgrowth. To substantiate this view, rescue experiments were then conducted by introducing KOR-expression vectors into the KOR-null DRG neurons.
In conducting these rescue experiments, we further hypothesized that KOR mRNA isoforms contribute differentially to maintain the physiological activities of KOR. For axon outgrowth, it was predicted that KOR mRNA that can be transported to axons would likely meet that need. Thus the rescue experiments were designed to test the ability of various KOR-expression vectors conferring different combinations of the 5′UTR and 3′UTR. Interestingly, the KOR expression vector that carried the intact 5′UTR and 3′UTR most efficiently rescued the axon outgrowth defect in KOR-null DRG neurons in response to EGF stimulation.34 Consistent with that finding, we found that axonal transport of KOR mRNA likewise required the combined presence of its 5′ and 3′-UTR.21 This series of experiments uncovered an important physiological activity of KOR—which is to mediate EGF-stimulated axon outgrowth during development. Moreover, this particular KOR activity is critically dependent upon the presence of KOR mRNA-specific UTR sequences known to be required for KOR mRNA's transport to axons and translational stimulation. Amazingly, a KOR expression vector deleted in the 3’-UTR failed, entirely, to rescue the axon-growth defect seen in KOR null DRG neurons, even though this expression vector is capable of making a functional receptor that can insert into the DRG neurons' somas (as well as other cells' surfaces) and bind its specific opioid ligands.23 In other words, to mediate cued axonal outgrowth, axonal KOR must be made from mRNA trafficked, deposited and translated only at the axon terminus, and only at the right time. These experiments strongly support our hypothesis of differential functions for KOR mRNA isoforms, and provide unambiguous evidence for their roles in mediating a very specific signaling pathway responding to the growth factor EGF. In our model, EGF first primes the neurons to engage its mRNA export machinery (for nuclear export). The exported KOR mRNA particles are then transported to the axon termini in excited neurons (for axonal transport). Finally, those KOR mRNA particles reaching the termini then respond to local stimulation (in the form of transient factors like axon guidance cues) to synthesize KOR protein. Moreover, a sequential execution of these processes is important for axon outgrowth. These studies are the first to determine a physiological function of KOR in a very specific process of neuronal growth and maturation during a critical time of development. These results also provide the first evidence for functional roles of specific untranslated mRNA sequences in integrating informational inputs from different sub-cellular compartments of a neuron in order to coordinate multiple cellular processes (nuclear export, axonal transport and local translation) for a critical function of the neuron, in this case, axon outgrowth.
Again, integration using various genetic, molecular, cellular and neurobiological approaches allows this complex issue to be addressed in a physiologically relevant context, which enables the delineation of the molecular pathways and identification of the required signals.
Concluding remark
Studies of fundamental processes in neurobiology have been challenging at multiple levels, including conceptual, technical, and biological. This is largely due to the enormous complexity of the architecture of individual neurons. Only recently has it been possible to begin substantiating the concept of compartmentalization in relation to not only the function but also the fundamental cellular processes of neurons. Functionality of a neuron provides the basic unit essential to the execution of a specific brain circuit, which is an even bigger problem that requires, perhaps, revolutionary concepts and methodologies. Dissecting the fundamental process in a specific compartment of neurons is the first step needed to understand the growth/maturation of neurons, as well as the underlying mechanisms of their activities.Compartmentalization not only segregates biological processes into various spatially and temporally controllable events (local events), but also enhances an ordered integration of various locally restricted biological processes into a “pathway” specifically relevant to certain functions or activities of neurons (systemic events). Issues are how the compartments and their local constituents are maintained and how these compartments communicate with each other. While structural constraints would facilitate the formation of a certain local “community”, the use of messengers such as RNA particles to direct or target certain molecules seems to be a clever strategy that could specify not only the spatial information but also temporal control of a specific molecule that is needed. An important feature of these RNA particles is that most of them contain various adaptor molecules, such as Grb7 and HuR, for binding target mRNAs. These adaptors provide critical functions in controlling local signal propagation (i.e., restricting local events) and communication among various compartments (such as coordination of soma response to EGF priming with axonal transport and translation). It is tempting to speculate that employing such interconnected and ordered biological processes across multiple compartments may constitute the basic unit of a functional pathway in a neuron. Integration of various methodologies and experimental tools to examine individual steps is essential to the realization of how such pathways operate in a physiological context.
Future challenges will be to understand how these pathways intersect, or interact, in order to create specific circuitry, how a brain uses these pathways in order to function, how locally produced KOR proteins participate in axon outgrowth, and how these pathways may go haywire and initiate disease processes. Predictably, integration of methodologies, tools, and models will be required to address these challenges in a specific physiological or pathological context.
Acknowledgements
I would like to thank F. Burton for help in editing and comment. This work is supported by NIH grants DA11190, DA11806, DK54733, DK60521, and K02-DA13926, the Philip Morris USA Inc. and Philip Morris International and the Distinguished McKnight University Professorship to L.-N. Wei.References
- A. P. Barnes and F. Polleux, Annu. Rev. Neurosci., 2009, 32, 347–381 CrossRef CAS.
- C. Conde and A. Cáceres, Nat. Rev. Neurosci., 2009, 10, 319–332 CrossRef CAS.
- Y. Jin and C. C. Garner, Annu. Rev. Cell Dev. Biol., 2008, 24, 237–262 CrossRef CAS.
- A. B. Dahlstrom, Prog. Neurobiol., 2010, 90, 119–145 CrossRef CAS.
- S. E. Ahmari, J. Buchanan and S. J. Smith, Nat. Neurosci., 2000, 3, 445–451 CrossRef CAS.
- M. Shapira, R. G. Zhai, T. s. Dresbach, T. Bresler, V. I. Torres, E. D. Gundelfinger, N. E. Ziv and C. C. Garner, Neuron, 2003, 38, 237–252 CrossRef CAS.
- S. Yoo, E. A. van Niekerk, T. T. Merianda and J. L. Twiss, Exp. Neurol., 2010, 223, 19–27 CrossRef CAS.
- L.-N. Wei, P. Y. Law and H. H. Loh, Front. Biosci., 2004, 9, 1665–1679 CrossRef CAS.
- A. C. Lin and C. E. Holt, Curr. Opin. Neurobiol., 2008, 18, 60–68 CrossRef CAS.
- P. Lobmaier, M. l Gossop, H. Waal and J. Bramness, Eur. J. Clin. Pharmacol., 2010, 66, 537–545 CrossRef CAS.
- P. Y. Law, H. H. Loh and L.-N. Wei, Neuropharmacology, 2004, 47, 300–311 CrossRef CAS.
- B. L. Kieffer and C. Gavériaux-Ruff, Prog. Neurobiol., 2002, 66, 285–306 CrossRef CAS.
- L.-N. Wei, Epigenetics, 2008, 3, 119–121 Search PubMed.
- T. W. Vanderah, Clin. J. Pain, 2010, 26, S10–S15 CrossRef.
- M. R. Bruchasa, B. B. Landa and C. Chavkin, Brain Res., 2010, 1314, 44–55 CrossRef.
- L.-N. Wei, X. Hu, J. Bi and H. H. Loh, Mol. Pharmacol., 2000, 57, 401–408 CAS.
- B. Sung, H. H. Loh and L.-N. Wei, Neurosci. Lett., 2000, 291, 163–166 CrossRef CAS.
- J. Bi, X. Hu, H. H. Loh and L.-N. Wei, J. Neurosci., 2001, 21, 1590–1599 CAS.
- J. Bi, X. Hu, H. H. Loh and L.-N. Wei, Mol. Pharmacol., 2003, 64, 594–599 CrossRef CAS.
- J. Bi, N.-P. Tsai, Y.-P. Lin, H. H. Loh and L.-N. Wei, Proc. Natl. Acad. Sci. U. S. A., 2006, 103, 19919–19924 CrossRef CAS.
- J. Bi, N.-P. Tsai, H.-Y. Lu, H. H. Loh and L.-N. Wei, Proc. Natl. Acad. Sci. U. S. A., 2007, 104, 13810–13815 CrossRef CAS.
- N.-P. Tsai, J. Bi and L.-N. Wei, EMBO J., 2007, 26, 1522–15231 CrossRef CAS.
- N.-P. Tsai, Y.-L. Lin, Y.-C. Tsui and L.-N. Wei, J. Cell Biol., 2010, 188, 325–333 CrossRef CAS.
- P. Anderson and N. Kedersha, J. Cell Biol., 2006, 172, 803–808 CrossRef CAS.
- M. J. Moore, Science, 2005, 309, 1514–1518 CrossRef CAS.
- J. R. Buchan and R. Parker, Mol. Cell, 2009, 36, 932–941 CrossRef CAS.
- N.-P. Tsai, P.-C. Ho and L.-N. Wei, EMBO J., 2008, 27, 715–726 CrossRef CAS.
- N.-P. Tsai, Y.-C. Tsui and L.-N. Wei, Neuroscience, 2009, 159, 647–656 CrossRef CAS.
- N.-P. Tsai, J. Bi, H. H. Loh and L.-N. Wei, J. Neurosci., 2006, 26, 9743–9749 CrossRef CAS.
- H. C. Chen, L.-N. Wei and H. H. Loh, Neuroscience, 1999, 92, 1143–1155 CrossRef CAS.
- F. Simonin, O. Valverde, C. Smadja, S. Slowe, I. Kitchen, A. Dierich, M. Le Meur, B. P. Roques, R. Maldonado and B. L. Kiefer, EMBO J., 1998, 17, 886–897 CrossRef CAS.
- B. Z. Shilo, Development, 2005, 132, 4017–4027 CrossRef CAS.
- Flores, H. Duan, H. Yan, R. Nagaraj, W. Fu, Y. Zou, M. Noll and U. Banerjee, Cell, 2000, 103, 75–85 CrossRef.
- N.-P. Tsai, Y.-C. Tsui, J. E. Pintar, H. H. Loh and L.-N. Wei, Proc. Natl. Acad. Sci. U. S. A., 2010, 107, 3216–2321 CrossRef CAS.
| This journal is © The Royal Society of Chemistry 2011 |