Micro OS-ELISA: Rapid noncompetitive detection of a small biomarker peptide by open-sandwich enzyme-linked immunosorbent assay (OS-ELISA) integrated into microfluidic device†
Masaki
Ihara‡
ae,
Amane
Yoshikawa
b,
Yushu
Wu
b,
Hiroko
Takahashi
d,
Kazuma
Mawatari
cd,
Kiyohito
Shimura
ce,
Kae
Sato
ce,
Takehiko
Kitamori
cde and
Hiroshi
Ueda
*abe
aDepartment of Bioengineering, School of Engineering, The University of Tokyo, 7-3-1 Hongo, Bunkyo, Tokyo, 113-8656, Japan. Fax: +81-3-5841-7362; E-mail: hueda@chembio.t.u-tokyo.ac.jp
bDepartment of Chemistry and Biotechnology, School of Engineering, The University of Tokyo, 7-3-1 Hongo, Bunkyo, Tokyo, 113-8656, Japan
cDepartment of Applied Chemistry, School of Engineering, The University of Tokyo, 7-3-1 Hongo, Bunkyo, Tokyo, 113-8656, Japan
dMicrochemistry group, Kanagawa Academy of Science and Technology, 3-2-1 Sakado, Takatsu, Kawasaki, Kanagawa, 213-0012, Japan
eCenter for NanoBio Integration, The University of Tokyo, 7-3-1 Hongo, Bunkyo, Tokyo, 113-8656, Japan
First published on 3rd November 2009
Abstract
A novel detection system that combines the merits of open-sandwich (OS) enzyme-linked immunoadsorbent assay (ELISA) and a microfluidic sensor chip system, and which enables rapid and noncompetitive immunodetection of small antigens of less than 1000 in molecular weight, has been proposed. Equipped with a sensitive thermal lens microscope, a minute amount of the carboxyl-terminal peptide of human osteocalcin (BGP), a biomarker for bone metabolism, was quantified utilizing antigen-dependent stabilization of an antibody variable region (OS principle). In a short analysis time (∼12 min), we could attain a detection limit comparable to that of the microplate-based OS ELISA (1 μg L−1). In addition, the effects of several pretreatments for serum-derived samples were investigated: an albumin absorption resin, addition of a protease inhibitor cocktail and heat treatment. Each pretreatment was found to be effective. Consequently, intrinsic BGP and its fragments could be detected in healthy human serum with a superior detection limit and working range compared to those of the conventional competitive ELISA method.
Introduction
Peptides are involved in biological systems in many ways and some of them are known as ideal biomarkers in body fluids.1–3 Peptides that serve as biomarkers include not only biologically active peptides, such as peptide hormones and neuropeptides, but also their degradation products, which show no apparent biological activity. Proteins with functional importance are subjected to posttranslational modification and degradation to regulate their half-lives, giving rise to smaller proteins and peptides with less or no apparent biological functions. The current proteomics works have been disclosing the significance of such peptide markers.4–7To date, there are a number of ways to assay small peptide molecules, for example, high-performance liquid chromatography (LC), mass spectroscopy (MS) and enzyme-linked immunosorbent assay (ELISA). Each of these methods has its merits and demerits. Although LC/MS instrumental analyses enable sensitive and high-throughput analysis of peptides in body fluids, they require large, costly equipment. In addition, because peptides in serum often undergo proteolytic cleavage at many sites, and thereby have a wide molecular weight distribution, the total concentration of the target peptide was often difficult to determine by instrumental analyses. On the contrary, ELISA is suitable for such purposes, and is suitable for point-of-care (POC) testing. Conventionally, two-types of ELISA have been commonly used, namely sandwich (non-competitive) and competitive ELISAs. While generally preferred, sandwich ELISA has a fundamental limitation in that the antigen must be large enough to have at least two epitopes to be captured by two kinds of antibodies, each either immobilized or enzyme-labeled. Therefore, small peptides with less than 10 amino acids are generally not detectable with the sandwich ELISA. To detect smaller molecules such as peptides, competitive ELISA employing antibodies that recognize single epitopes is used, where either an enzyme-labeled antigen or antibody is used as a competitor. While competitive ELISA can determine the total concentration of peptides bearing an epitope of interest, it often suffers from high detection limits, and the difficulty to optimize the reaction conditions due to its narrow working range.
As an alternative powerful immunoassay for small antigens, we have proposed a noncompetitive open-sandwich immunoassay (OS-IA), which is based on the principle of stabilization of the antibody variable region, VH and VL, upon binding with antigen (Fig. 1).8 In an enzyme-immunosorbent assay based on the open-sandwich principle (OS-ELISA), a small antigen with a MW of around 200–1000 can be noncompetitively detected by the sandwich formed between the immobilized VL and enzyme-labeled VH, and the antigen-dependent formation of a stable ternary complex is detected by the immobilized enzyme activity. Using OS-ELISA, various molecules have been measured with a superior detection limit and a working range compared with those achieved with the corresponding competitive ELISA.
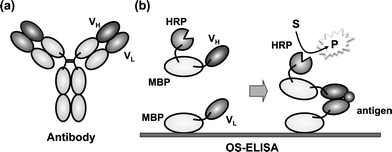 | ||
| Fig. 1 Principle of the open sandwich immunoassay. (a) Schematic structure of the whole antibody. (b) Open-Sandwich ELISA with immobilized MBP-VL and HRP-conjugated MBP-VH (MBP-VH-HRP). Antigen and MBP-VH-HRP are simultaneously added to the MBP-VL immobilized to a microplate or polystyrene beads. Antigen-dependent binding of MBP-VH-HRP is detected by peroxidase activity. | ||
As a model biomarker peptide, here we focus on the human osteocalcin C-terminal peptide, which was successfully detected by OS-ELISA with a superior sensitivity and working range compared with those of the competitive assay.9 Human osteocalcin, also known as the bone Gla (γ-carboxyl glutamic acid) protein or BGP, is the major non-collagen protein of bone composed of 49 amino acids. BGP is produced predominantly by osteoblasts in the mineralization process.10 Although the precise function of BGP in bone metabolism is not known, BGP is considered as a marker for bone metabolism based on a relationship between serum BGP and bone formation.10,11 A recent report also showed an alternative role of BGP in regulating energy metabolism.12 On the other hand, Novak et al. have reported that plasmin, a serine protease activating latent matrix metalloproteases or growth factors, specifically cleaves BGP at the C-terminal site.13,14 BGP cleavage by plasmin occurs not only in solution but also when bound to hydroxyapatite, which promotes the detachment of digested BGP products from hydroxyapatite. The plasmin-mediated lysis of the free and hydroxyapatite-bound BGP could be responsible for the abundant NH2-midterminal (1–43) and COOH-terminal (44–49) peptides in serum. The C-terminal peptide of BGP (BGP-C) was also suggested to function as a regulator for bone remodeling.15 It is assumed that plasmin cleavage of BGP plays a role in bone homeostasis, and BGP-C will provide us meaningful information that cannot be drawn from full-length BGP.
Previously, we demonstrated that OS-ELISA using conventional 96-well microplates can detect BGP-C peptides (longer than 6 AA) with a wider working range and a lower detection limit (0.1–500 μg L−1) than the corresponding competitive ELISA.9 However, the assay using bulk plates required a reasonable assay time, troublesome liquid-handling procedures, and the use of relatively large amounts (>0.2 μg/well) of each antibody reagent. In this study, to develop a POC testing device to analyze a microliter droplet of donor serum in a short measurement time, we have integrated OS-ELISA with a microfluidic sensor chip system equipped with a thermal lens microscope (TLM) detector. The microchip-based immunoassays (sandwich and competitive assays) were proven to simplify the assay procedure, reduce the assay time, and lower the consumption of samples, reagents, and energy, with the achievements of low detection limits (pM levels for proteins, while several nM for a hapten d-methamphetamine).16–20 These merits are expected to reduce the burden on patients, and be important for implementing POC testing.
In this study, we firstly attempted to detect heptameric BGP-C in bovine and human sera by OS-ELISA using bulk plates, and found the existence of some factors that inhibit the immunoreaction in the serum. To remove such inhibitory factors, several pre-treatments of serum, including an albumin absorption resin, a protease inhibitor cocktail and heating, were tested, and optimized for the reaction in the microchip.
Experimental section
Materials
Fetal bovine serum (ϕ 0.1 mm membrane filtered) was purchased from Biological Industries (Haemek, Israel). Human blood samples were collected from two healthy male donors using a Venoject II vacuum blood-collecting tube (Terumo Co., Tokyo, Japan), and allowed to clot. Intact BGP was obtained from Princeton Biomolecules (Langhorne, PA). Synthetic peptides for the C-terminal heptamer (NH2-RRFYGPV-COOH, BGP-C7) and dodecamer (NH2-GFQEAYRRFYGPV-COOH, BGP-C12) of BGP were from GenScript Co. (Piscataway, NJ).Preparation of MBP-VL and MBP-VH fusion proteins
The two variable domains (VH and VL) of the anti-BGP antibody KTM2199 were expressed as fusion proteins with E. coli maltose-binding protein (MBP). The expression of VL fused with MBP (MBP-VL) was performed by transforming E. coli BL21(DE3)pLysS with the pMAL-VL(BGP) vector as described.9 Briefly, the transformant was cultured in 10 mL of LBAG medium (10 g L−1 tryptone, 5 g L−1 yeast extract, 5 g L−1 NaCl, pH 7.5 supplemented with 100 μg mL−1 ampicillin and 1% glucose) overnight at 30 °C. Subsequently, the culture was inoculated to 800 mL of LBA medium (10 g L−1 tryptone, 5 g L−1 yeast extract, 5 g L−1 NaCl and 100 μg mL−1 ampicillin) and incubated at 20 °C until OD600 reached 0.6–0.8, when the expression was induced by the addition of isopropyl-β-d-thiogalactopyranoside (IPTG; Wako, Osaka, Japan) to a final concentration of 0.5 mM. The overnight culture at 20 °C was centrifuged, and the bacterial pellet was resuspended with 50 mL of osmotic shock solution (30 mM Tris- HCl, 20% sucrose, 1 mM EDTA, pH 8.0). The suspension was incubated at 25 °C for 10 min with frequent shaking followed by centrifugation at 8000 g, 4 °C for 10 min. Supernatant was discarded, and 15 mL of ice-cold 5 mM MgSO4 was added immediately to resuspend the cells. The suspension was incubated for 20 min at 4 °C or on ice with frequent shaking and centrifuged at 8000 g, 4 °C for 15 min. The supernatant obtained was then dialyzed against column buffer (50 mM phosphate, 500 mM NaCl, pH 7.4), applied to a column (1 cm × ϕ 1 cm) packed with TALON affinity resin (Clontech Laboratories, Inc., Mountain View, CA), washed with column buffer and eluted using elution buffer (column buffer containing 500 mM imidazole). The fractions containing target protein were pooled, and dialyzed against PBS (10 mM phosphate, 137 mM NaCl, 2.7 mM KCl, pH 7.4). The purity of the protein was confirmed by 10% SDS-PAGE, and stored in small aliquots at −80 °C.To obtain the pET-based expression vector for MBP-VH(BGP), pET-MBPp-VH(BPA)21 and pKST2(BGP)9 were used. A periplasmic expression vector pET-MBPp-VH(BPA) to secrete MBP-VH (for bisphenol A, BPA) protein was digested with NcoI and XhoI to remove the fragment encoding the VH for BPA, and ligated with the NcoI-XhoI fragment of pKST2(BGP) encoding the VH for BGP, resulted in pET-MBPp-VH(BGP). Following the confirmation of the inserted DNA sequence, the transformation of E. coli BL21(DE3)pLysS, the expression and purification of MBP-VH were performed as above.
Purified MBP-VH was labeled with horseradish peroxidase (HRP) using the peroxidase labeling kit-NH2 (Dojindo, Kumamoto, Japan) according to the manufacturer's instructions, purified using the TALON affinity resin, and buffer-exchanged with PBS. The HRP-labeled MBP-VH was concentrated, and glycerol added to a final concentration of 50%. The resulting HRP-labeled MBP-VH was stored at −20 °C prior to use.
Microfluidic device
OS-ELISA was performed in a microfluidic channel (200 μm in width) in which polystyrene microbeads (45 μm in diameter, JSR, Tokyo, Japan) were packed using the automated ELISA system.16,22 The microfluidic channel with a dam structure for retaining the microbeads was fabricated by photolithography and HF etching on a Pyrex glass chip (3 × 7 cm) (Fig. 2). For the sensitive colorimetric detection of the blue reaction product in the micro-channel, a laser-induced thermal lens microscope (TLM) was employed. The wavelengths of the excitation and probe beams were 658 nm and 785 nm, respectively. The probe beam intensity was monitored with a photodiode.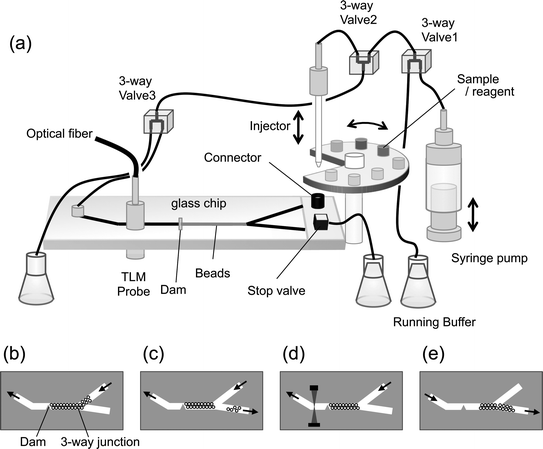 | ||
| Fig. 2 micro-ELISA system. (a) A glass chip (3 × 7 cm) integrated with microfluidic channels (200 μm × 200 μm) was used for the OS-ELISA reaction. (b) Polystyrene beads with immobilized MBP-VL were introduced from the inlet (connector) and stored between a dam structure and the three-way junction, and (c) extra beads were flushed through the stop valve. MBP-VH-HRP and antigen on the rotating stage were introduced into the microchip to form an immuno-complex on the beads. (d) After washing, the substrates were applied and dye molecules produced by the enzyme reaction were detected by a TLM downstream of the dam structure. (e) The microbeads after each assay was flushed by an inversed flow. | ||
Serum pretreatments
To remove albumin from serum, Affi-Gel BlueGel (designated BlueGel, BioRad, Hercules, CA) was used. A spin column packed with BlueGel (0.5 cm × ϕ 0.5 cm) was washed 3-times with PBS, and spun-down at 2000 g for several seconds. Subsequently, 50 μL of bovine or human serum was applied to the column and then eluted by spin-down. To inactivate proteases in the serum, Complete Mini Protease Inhibitor Cocktail Tablets (Roche Diagnostics, Indianapolis, IN) were added to the serum sample with one-seventh of the volume of the 8× Complete Mini Protease Inhibitor Cocktail working solution. In the heat pretreatment step, sera in microtubes were incubated in a DTU-1B block heater (Taitec, Saitama, Japan) at 65 °C, 10 min.To remove BGP fragments with MW of less than 3000 from serum, the serum was dialyzed against PBS overnight at 4 °C using a Slide-A-Lyzer 3 K MWCO (Thermo Fisher Scientific Inc., Waltham, MA). To prepare BGP-free serum, the serum was filtered through a spin-column packed with 10 vol. of anti-BGP antibody-agarose beads, which was prepared by reacting 0.2 mg mL−1 of KTM219 IgG with NHS-activated Sepharose 4 Fast Flow (GE Healthcare) in 0.1 M sodium carbonate buffer (pH 8.3) at 4 °C for 3 h.
OS-ELISA on a bulk plate
A 96-well flexible microplate (BD Falcon, Franklin Lakes, NJ) was incubated with 50 μL of 10 μg mL−1 MBP-VL in PBS overnight at 4 °C or at 25 °C for 1 h, and blocked with 200 μL of PBS containing 20% Immunoblock (PBS-IB, DS Pharma Biomedical, Osaka, Japan) at 25 °C for 2 h. After washing with PBS containing 0.1% Tween-20 (PBST), the immobilized MBP-VL was reacted with a mixture of 20 μL of treated or non-treated serum, 20 μL of PBS containing BGP-C7 at various concentrations, and 40 μL of 5 μg mL−1 MBP-VH-HRP in 10% PBS-IB at 25 °C for 1 h. Subsequently, the plate was washed with PBST, and added with 50 μL of 3,3′,5,5′-tetramethylbenzidine (TMB) One solution (Promega Co., Madison, MA). The enzyme reaction results in color development with intensities proportional to the amount of the ternary immuno-complex, namely, MBP-VL/BGP-C7/MBP-VH-HRP (Fig. 1). To stop the enzyme reaction, the plate was added with 50 μL of 1% H2SO4, and the absorbance at 450 nm was measured using a Model 680 plate reader (BioRad) and compared with the absorbance at 655 nm as a control.To evaluate the effect of albumin on the OS reaction, the plate immobilized with MBP-VL was incubated with the mixture of the following solutions in PBS, 20 μL of 0–16% bovine serum albumin (BSA, Fraction V, Sigma, St. Louis, MO), 20 μL of BGP-C7 at various concentrations and 40 μL of 5 μg mL−1 MBP-VH-HRP at 25 °C for 1 h. The immuno-complex was detected as above.
OS-ELISA on the microchip
A schematic illustration of the bead-based micro-ELISA system is shown in Fig. 2. First, the suspension of beads were physically-coated with MBP-VL (10 μg mL−1 in PBS for 4 °C, 16 h) was and then introduced into the dam region of the microchip via the inlet hole. Excess beads were washed away by flushing with 20% PBS-IB from one inlet to the other. After blocking by flushing with 20% PBS-IB at 30 μL min−1 for 10 s, 5 μL of the sample, 5 μL of PBS containing BGP-C7 at various concentrations, and 10 μL of 5 μg mL−1 MBP-VH-HRP conjugate in 5–15% PBS-IB were introduced at 1 μL min−1 for 5 min for the OS reaction. After washing the channel at a flow rate of 40 μL min−1 for 30 s the enzyme substrate SureBlue TMB (Kirkegaard & Perry Laboratories, Inc., Gaithersburg, MD) was pumped into the channel, and reacted without flow for 1 min. Then, the accumulated product was flowed through the channel and detected using the downstream TLM, where the peak intensity was used as a signal.Sandwich ELISA on a bulk plate
The microplate was incubated with 50 μL of 1 μg mL−1 KTM219 in PBS at 25 °C for 1 h, blocked with 20% PBS-IB at 25 °C for 2 h, washed with PBST, and then incubated with 25 μL of treated or non-treated serum and 25 μL of PBS containing 10% immunoblock. Following washing with PBST, the reaction with 50 μL of 1 μg mL−1 biotinylated anti-BGP N-terminus antibody KTM2239 in PBS was conducted for 1 h, and the immuno-complex was probed with the streptavidin-HRP conjugate (Vector Laboratories, Burlingame, CA) and TMB One solution.Results
Pretreatment of bovine serum
In our previous study, we reported the noncompetitive detection of C-terminal BGP peptides by OS-ELISA.9 To extend the assay to a practical diagnosis, we first tried to determine the BGP concentration in non-treated bovine and human sera by OS-ELISA, and found that some factors in serum inhibit the OS reaction. To clarify the inhibitory factors, firstly we focused on albumin as a potential inhibitor, because albumin is the most abundant serum constituent (40–50% w/v) and studied the effects of albumin on the viscosity and permittivity. To reduce the albumin concentration without affecting the intrinsic BGP level, we examined the effect of several commercially available albumin-absorption gels including BlueGel, which is agarose beads crosslinked with covalently Cibacron Blue F3GA dye (CB). CB is known to bind strongly to albumin mainly by hydrophobic interactions between the anthraquinone chromophore in CB and the nonpolar surfaces of albumin. In addition, a weak interaction between CB and BGP was expected, because the negatively-charged intact BGP (isoelectric point = 4.0–4.5) repels negatively-charged sulfonate groups in CB.The efficient removal of albumin from bovine serum by BlueGel was confirmed by SDS-PAGE (Fig. 3). The band corresponding to albumin was markedly reduced after the BlueGel-treatment, whereas no other significant changes were observed for other bands. In a microplate-based OS-ELISA for the heptameric BGP-C-terminal peptide BGP-C7 in 20% BlueGel-treated bovine serum, the signals were significantly increased compared with those in the same amount of non-treated serum, giving a dose-response curve overlapping with that in PBS (Fig. 4A). The detection limit (>3 × SD of the background) of OS-ELISA was estimated to be ∼5 μg L−1 BGP-C7 in BlueGel-treated bovine serum and PBS.
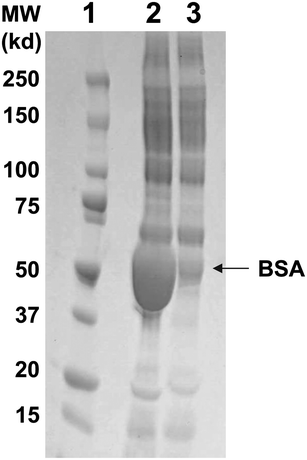 | ||
| Fig. 3 Effect of BlueGel treatment. Lane 1: Molecular weight markers, Lanes 2–3: Non-treated and BlueGel-treated bovine sera. A dominant band in lane 2 corresponds to BSA. | ||
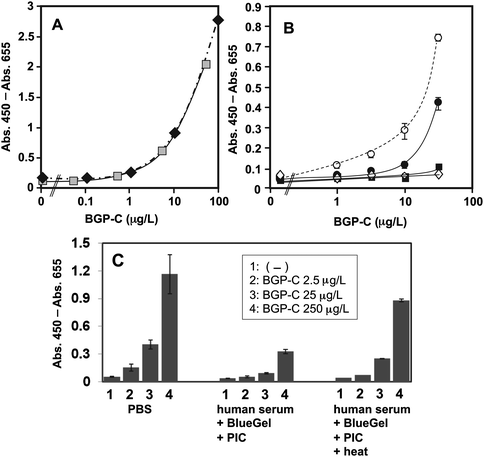 | ||
| Fig. 4 (A) Microplate OS-ELISA in PBS or in the presence of pretreated bovine serum. Black diamonds and gray squares represent BGP-C7 signals on the background of PBS and 25% BlueGel-treated bovine serum in PBS, respectively. The signal intensities were higher than those of other microplate assays due to longer incubation time in HRP reaction. (B) The albumin concentration dependency of OS-ELISA signals. Open circles, closed circles, closed squares and open diamond denote signals in the presence of 0, 1, 2 and 4% (w/v) BSA in PBS containing 5% immunoblock, respectively. Averages of three samples with 1 SD are shown. (C) Effects of pretreatments on microplate OS-ELISA in the presence of human serum. From the left to right, three sets of OS-ELISA in PBS, 25% human serum in PBS treated with BlueGel and PIC, and the same with additional heat treatment are shown. (1)-(4) Final 0, 2.5, 25, and 250 μg L−1 BGP-C7, respectively, were added to the wells immobilized with MBP-VL. An average of three samples with 1 SD is shown. | ||
To further confirm the inhibitory effect of pure albumin, OS-ELISA in the presence of commercially available BSA was performed. As shown in Fig. 4B, the signal intensities remarkably decreased when the BSA concentration was equal to or more than 1 (w/v)%. Since the BSA concentration in 20% serum is 6–10 (w/v)%, it can be a major factor for the inhibition of the OS reaction in bovine serum.
Pretreatment of human serum
We next tried to measure BGP-C7 in the BlueGel-treated human serum by OS-ELISA using a bulk microplate. However, in this case, a significant reduction in signal intensity was observed even after BlueGel treatment, indicating the existence of other inhibitory factor(s) (Data not shown). Since bovine serum had been already heat-inactivated for 56 °C, 30 min, we examined the effects of the protease inhibitor cocktail (PIC) and heat (65 °C, 10 min) on human serum. Heating is a well-established method of inactivating the complement in immunoassays, and often results in improved immunoassay signal intensity.23,24As expected, the signal intensities were significantly improved by adding these steps after BlueGel-treatment, and the combination of three treatments (BlueGel -> PIC -> heat) gave the most sensitive dose-response (Fig. 4C).
Protease activity in human serum
Serum is known to be abundant in various intrinsic proteases. To confirm the proteolytic activity in human serum and the effect of its inhibitor PIC, BGP-C7 and MBP-VL in human serum after incubation at 25 °C for various periods were analyzed by mass spectrometry and Western blotting, respectively. The digestion of BGP-C7 was confirmed in the 10-fold diluted non-treated human serum after 12 h (Fig. S1A†), whereas no significant change was observed in the PIC-treated serum (Fig. S1B†). On the other hand, the digestion of MBP-VL was observed in the 100-fold diluted non-treated serum within 1 h, whereas the shift in the PIC-treated serum sample was observed only after 10 h (Fig. S3†). These results indicated that some protease in human serum digested MBP-VL over a relatively short period, which was one of the key inhibitory factors at least in this case.Effect of the pre-treatments on BGP concentration
To examine if these treatments affect BGP-C peptides in PBS or not, OS-ELISA was performed. In addition, the change of intact BGP concentration in serum during the treatments was investigated by sandwich ELISA, whose signal is not affected by up to 30% of non-treated serum in a sample (data not shown). Intact BGP and BGP-C7 (10 μM each) were serially treated with BlueGel, PIC, and heat at 65 °C for 10 min, and the aliquots after each treatment were assayed. As a result, no significant changes in signals were observed between the treatments, and the sample loss during each treatment was within 10% (Fig. S2†).OS-ELISA on the microfluidic chip
Based on these observations, we tried to apply OS-ELISA to the microfluidic chip. We began with the optimization of the measurement conditions in PBS, and applied these conditions to that in serum. Firstly, two methods for the immobilization of MBP-VL were examined, namely, physical absorption to polystyrene beads and the immobilization of the biotinylated protein to streptavidin-coated beads. In the former case, MBP-VL in PBS was mixed with the beads solution overnight, and PBS-IB added. In the latter case, biotinylated BSA was physically absorbed to the beads, and streptavidin was added to the beads to form the biotin-streptavidin complex. Biotinylated MBP-VL was then added to the beads and these were then blocked by PBS-IB. While the result of OS-ELISA using the latter beads showed somewhat larger signals than that of the former beads, it also showed some difficulty in achieving reproducibility and usefulness, and a higher background signal. These drawbacks are probably due to its more complicated hypermolecular structure and/or difficulty to prepare biotinylated MBP-VL without affecting its activity. Given the poor results of these beads, the former physical immobilization method was chosen instead.The detection limit was influenced by the incubation/washing conditions and the blocking agent concentration in the immuno-reaction. We investigated the effect of the flow rate (20 and 40 μL min−1) and the duration (30 and 60 s) in the washing process. As a result, the highest ratio of signal to background (S/B ratio) in a lower concentration (10 μg L−1) of BGP-C7 was observed in the washing with 40 μL min−1 for 30 s. The reactions of MBP-VH-HRP, BGP-C7, and MBP-VL beads were also carried out in the presence of various concentrations of Immunoblock in PBS, and final concentration of 5% gave the highest S/B ratio. Hence we used these conditions in the following microchip experiments.
MBP-VH-HRP at the various concentrations was introduced into the microfluidic chip at various flow rates and the resulting S/B ratios were evaluated. Consequently, it was found that the reaction with 5 μg mL−1 MBP-VH-HRP at 2 μL min−1 led to the best result. By combining the above conditions, detection of 1.0 μg L−1 BGP-C7 in PBS, which was equivalent to the detection limit in the OS-ELISA on the bulk microplate, was achieved (Fig. 5A).
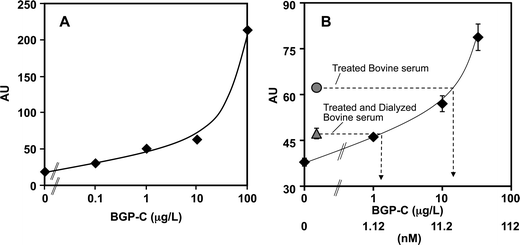 | ||
| Fig. 5 Micro OS-ELISA. (A) The mixture of MBP-VH-HRP (10 μL) and various concentrations of BGP-C7 (10 μL) in PBS were introduced to the microfluidic channel, and reacted with MBP-VL immobilized on beads. After washing and enzymatic reaction, the signals detected by TLM were plotted against BGP-C7 concentration (diamonds). (B) MBP-VH-HRP (10 μL), various concentrations of BGP-C7 (5 μL) and bovine serum treated with BlueGel and anti-BGP agarose (5 μL) were introduced and the signals obtained were plotted against the BGP-C7 concentration (diamonds) to prepare a standard curve. To estimate the concentration of BGP in bovine serum, OS-ELISA was performed in the same way, except for using PBS (5 μL) and BlueGel-treated bovine serum (5 μL) instead of BGP-C7 and serum solutions (gray circle). The BGP concentration after dialysis was estimated by using dialyzed serum and BlueGel-treated serum (gray triangle). | ||
Detection of BGP in bovine serum by OS-ELISA in a microfluidic chip
We first carried out OS-ELISA in the presence of 25% BlueGel-treated bovine serum in PBS in the microfluidic chip based on the condition optimized above. The serum mixed with BGP-C7 and MBP-VH-HRP was introduced into the microfluidic channel packed with beads pre-coated with MBP-VL, and the amount of ternary immuno-complex was detected by a SureBlue TMB substrate and TLM. As a result, the signal intensities and the detection limit (1.0 μg L−1 BGP-C7) were comparable with those in the assays using the corresponding samples in PBS.To quantify the intrinsic BGP in bovine serum, a standard curve was drawn with defined amounts of BGP-C7 diluted in 25% BGP-free serum, which was prepared by filtering the BlueGel-treated bovine serum through an anti-BGP spin column, which was packed with agarose beads immobilized with the anti-BGP antibody KTM219 (Fig.5B). To detect intrinsic BGP and its fragments, BlueGel-treated serum was measured in the same way without adding BGP-C7. Consequently, the total molar concentration of BGP and its C-terminal fragments in bovine serum BGP was calculated to be approximately 17 nM (equivalent to 15 μg L−1 BGP-C7).
Since this concentration evaluated by OS-ELISA would be the sum of both intact and fragmented BGPs, we dialyzed the serum with a dialysis membrane with a molecular weight cut-off of 3 kDa, to remove BGP fragments smaller than 3 kDa. When the dialyzed serum was assayed, the signal in OS-ELISA was significantly decreased to 1.4 nM (1.2 μg L−1 BGP-C7), suggesting that the difference between the non-dialyzed and dialyzed samples reflects the concentration of low-molecular-weight BGP fragments.
Determination of BGP in human serum by OS-ELISA on the microfluidic chip
The condition described above was applied to OS-ELISA to assay human serum. However, the detection limit was inferior to those in the bovine serum assay. We further optimized the Immunoblock concentration in the immuno-reaction, and improved the detection limit (2 μg L−1 BGP-C7) by reducing the final concentration to 2.5%. Using the BGP-free serum prepared by passing the pre-treated human serum through an anti-BGP spin column, a standard curve for BGP-C7 was made (Fig. 6A), and the signal intensities of the dialyzed and non-dialyzed pre-treated human sera were compared. Consequently, the BGP concentrations in the non-dialyzed and dialyzed human sera were estimated to be 10 and 3 nM (9 and 3 μg L−1 of BGP-C7), respectively (Fig. 6A), suggesting that the concentrations of the total BGP and the fragmented BGP-C were at 10 and 7 nM, respectively.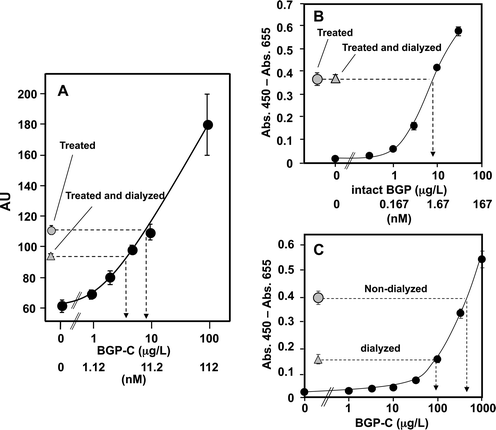 | ||
| Fig. 6 (A) Micro OS-ELISA of the treated human serum. MBP-VH-HRP (10 μL), the BGP-C7 solution (5 μL) and the human serum treated with BlueGel, PIC, heat, and KTM219-immobilized agarose (5 μL) were introduced into the microchip and the signals obtained were plotted against the BGP-C7 concentration (black circles) to prepare a standard curve. Also, the human serum (5 μL) treated with BlueGel, PIC, and heat was mixed with MBP-VH-HRP (10 μL) and PBS (5 μL), and introduced into the microchip to estimate the intrinsic BGP-C concentration including the full-length BGP in the serum (gray circle). To remove small BGP-C peptides, the pretreated serum was dialyzed and then introduced into the microchip (gray triangle). An average of the two samples with 1 SD is shown. (B) Negligible effect of dialysis on intact BGP. The triple-treated BGP-free human sera added with BGP at various concentrations were applied to conventional sandwich ELISA using microplate. The dose-response (black circles) curve is shown with the data from the triple-treated, non-dialyzed sera (gray circle) and triple-treated, dialyzed sera (gray triangle) without added BGP. An average of the three samples with 1 SD is shown. (C) Effect of dialysis on BGP-C7. OS-ELISA of the BGP-C7 solutions in PBS was performed using a microplate (black circles). The PBS containing 500 μg L−1 of BGP-C7 was dialyzed (gray triangle) or incubated (gray circle) at 4 °C for 16 h. | ||
The comparison of BGP concentrations in the pre-treated human sera before and after the dialysis was also performed on a bulk microplate. When the intact BGP concentration was compared by the conventional sandwich assay using two antibodies specific to the N-terminal and the C-terminal peptides, the estimated concentrations of the dialyzed and non-dialyzed samples were essentially identical and assayed to be approximately 1.5 nM (9 μg L−1 intact BGP) (Fig. 6B). The values were within the reported range of healthy individuals, but were significantly lower than those estimated by OS-ELISA for dialyzed samples (3 nM). We reasoned that the difference in values estimated by the two methods was partly due to the remaining BGP-C7 fragments after dialysis.
To estimate the efficiency of dialysis, BGP-C7 was added to PBS to a final concentration of 500 μg L−1. The samples were then dialyzed and the amount of remaining BGP-C7 was estimated by OS-ELISA. As shown in Fig. 6C, the signal in OS-ELISA decreased after dialysis and the remaining BGP-C7 was estimated to be ∼90 μg L−1, indicating the dialysis efficiency of 82%. BGP-C7 also was added to serum to a final concentration of 50 mg L−1, dialyzed, and the amount of remaining BGP-C7 was estimated by MALDI-TOF MS. When the signal intensities were normalized with the internal standard (BGP-C12) added after dialysis, the comparison between non-dialyzed and dialyzed samples revealed that the peak for BGP-C7 of the dialyzed sample decreased to 22% (Fig. S1C†). Taken together, the dialysis efficiency was estimated to be ∼80%, which can well explain the over-estimated concentration of the dialyzed samples in OS-ELISA. Although the amount of remaining BGP fragments was not significant for the estimation of BGP-C concentrations, the intact BGP concentration should be thus corrected.
Discussion
In conventional immunoassays, small peptides have almost always been assayed by competitive assays in a microplate format. However, their low sensitivity, relatively large sample volume and long-measurement times were considered as problems. Recently, we reported the use of OS-ELISA for BGP-C peptides and demonstrated that the detection limit was approximately 1/10 of that in the corresponding competitive ELISA.9 In this study, by integrating OS-ELISA with a microfluidic chip, we further succeeded in reducing the sample volume to 1/10 (50 μL to 5 μL) and reducing the analysis time to 1/20 (4 h to 12 min). Consequently, the detectable amount of peptide using the OS-ELISA/integrated microchip system was 1/100 of that obtained using the competitive ELISA on microplates. These improvements are of critical importance in applications to POC testing, rapid diagnosis and therapy in clinical studies.We also demonstrate that OS-ELISA enables the detection of BGP-C peptides in treated serum in a wide concentration range of 1–1000 nM, and determined the total BGP-C concentrations in bovine and human serum samples (17 and 10 nM, respectively) (Fig. 5B and 6A). The dialysis (cut off MW < 3 kDa) of the treated sera resulted in reduced signals in OS-ELISA, which corresponded to 1.4 and 3 nM in bovine and human sera, respectively, whereas no significant influence was observed for the sandwich assay of human serum (Fig. 6B). These observations support the assumption that OS-ELISA gives the total BGP (C-terminally fragmented and intact) concentration, and the sandwich assay evaluates the intact BGP concentration. The intact BGP was labile to serum proteases25–28 and the concentration can easily vary according to individual difference on protease activity or sample handling.10,29–31 On the other hand, the total BGP concentration is considered relatively steady and reflects the BGP liberation rate more faithfully. This makes the total BGP a useful diagnostic criterion.10,32 Furthermore, the difference between non-dialyzed and dialyzed sera implies the concentration of fragmented BGP, which enables the simultaneous determination of total and BGP-C fragments. Several groups suggested that BGP fragments in serum play physiological roles in bone remodeling and are novel meaningful biomarkers.14,15 Hopefully, OS-ELISA for total and fragmented BGP becomes an important diagnostic item.
In this study, we showed the successful determination of serum BGP-C peptide by micro OS-ELISA. The inhibitory factors in serum against OS-ELISA can be effectively removed by three consecutive pretreatments. BlueGel was successfully used to remove the inhibition factor, serum albumin. A possible reason is the viscosity of the solution containing a high concentration of albumin, which can prevent the efficient immuno-reaction. Another possibility is the non-specific interaction of albumin with the hydrophobic region of VH or VL, especially at their VH/VL interfaces, since albumin is known to interact with many molecules by hydrophobic interaction.33,34 However, since it was not an apparent problem in the case of the fluorescence resonance energy transfer (FRET)-based OS immunoassay using the anti-lysozyme antibody HyHEL-10,35 the inhibitory effect might depend on the nature of the antibody used.
Another inhibitory factor was protease, which can digest MBP-VH-HRP, MBP-VL, or BGP-C7 to inhibit the formation of the immuno-complex. So far, it is reported that target peptides or exogenous proteins are often rapidly degraded by proteolytic enzymes in serum.29,31,36,37 In fact, the protease activity in diluted non-treated human serum was confirmed by tracing the digestion of BGP-C7 and its inhibition with PIC by MALDI-TOF MS (Fig. S1A†), and that of MBP-VL by Western blotting (Fig. S3†). Therefore, the digestion of BGP-C7 or MBP-VL/VH should be one of the inhibition factors. Redesign of the fusion proteins that are resistant to proteases might be an effective way to overcome this problem.
Though these pre-treatments seem tedious, the removal of hemocytes from the blood sample, pre-treatments with BlueGel, PIC and/or heat can, in principle, be fully automated in the microchip system. For example, the deprivation of hemocytes and albumin, as well as heat-inactivation can be serially achieved by pumping the blood into the hemocyte filtering unit, the BlueGel-packed unit and heating unit designed in a microchip channel. Subsequently, the serum can be mixed with a reservoir containing protease inhibitors and MBP-VH-HRP in the microchip, and then sent to the MBP-VL immobilized beads unit and analyzed as done by the fully-automated ELISA system used in this study. Such total automation for the serum diagnosis would be essential for attaining high reproducibility and simple handling.38
In this study, we successfully optimized the conditions for an OS-based serum assay in a microchip device and provided a milestone toward the practical application of this assay. The analysis for the serum peptides by OS-ELISA would reveal profiles of various bioactive substances.
Acknowledgements
We thank Mr. Tatsuya Shinoda of Kyowa Medex Co. for the generous gift of anti-BGP IgGs and the genes derived from them. This study was supported in part by the Japan Science and Technology Agency (PRESTO program), and by a Grant-in-Aid for Scientific Research (B17360394 and B20360368) from MEXT, Japan. We also thank FUJIFILM Co. for financial support.References
- A. Martorella and R. Robbins, Acta Biomed., 2007, 78(Suppl. 1), 123–128 Search PubMed.
- I. Schulte, H. Tammen, H. Selle and P. Schulz-Knappe, Expert Rev. Mol. Diagn., 2005, 5, 145–157 Search PubMed.
- J. Villanueva, J. Philip, D. Entenberg, C. A. Chaparro, M. K. Tanwar, E. C. Holland and P. Tempst, Anal. Chem., 2004, 76, 1560–1570 CrossRef CAS.
- J. N. Adkins, S. M. Varnum, K. J. Auberry, R. J. Moore, N. H. Angell, R. D. Smith, D. L. Springer and J. G. Pounds, Mol. Cell. Proteomics, 2002, 1, 947–955 CrossRef CAS.
- N. L. Anderson and N. G. Anderson, Mol. Cell. Proteomics, 2002, 1, 845–867 CrossRef CAS.
- W.-C. Chang, P.-I. Hsu, Y.-Y. Chen, M. Hsiao, P.-J. Lu and C.-H. Chen, Proteomics: Clin. Appl., 2008, 2, 55–62 Search PubMed.
- R. S. Tirumalai, K. C. Chan, D. A. Prieto, H. J. Issaq, T. P. Conrads and T. D. Veenstra, Mol. Cell. Proteomics, 2003, 2, 1096–1103 CrossRef CAS.
- H. Ueda, K. Tsumoto, K. Kubota, E. Suzuki, T. Nagamune, H. Nishimura, P. A. Schueler, G. Winter, I. Kumagai and W. C. Mohoney, Nat. Biotechnol., 1996, 14, 1714–1718 CrossRef CAS.
- S. L. Lim, H. Ichinose, T. Shinoda and H. Ueda, Anal. Chem., 2007, 79, 6193–6200 CrossRef CAS.
- J. L. Allison, S. Hodges and R. Eastell, Ann. Clin. Biochem., 2000, 37, 432–446 CrossRef.
- M. S. Calvo, D. R. Eyre and C. M. Gundberg, Endocr. Rev., 1996, 17, 333–368 CAS.
- N. K. Lee, H. Sowa, E. Hinoi, M. Ferron, J. D. Ahn, C. Confavreux, R. Dacquin, P. J. Mee, M. D. McKee, D. Y. Jung, Z. Zhang, J. K. Kim, F. Mauvais-Jarvis, P. Ducy and G. Karsenty, Cell, 2007, 130, 456–469 CrossRef CAS.
- J. F. Novak, J. D. Hayes and S. K. Nishimoto, J. Bone Miner. Res., 1997, 12, 1035–1042 CrossRef CAS.
- J. F. Novak, M. B. Judkins, M. I. Chernin, P. Cassoni, G. Bussolati, J. A. Nitche and S. K. Nishimoto, Cancer Res., 2000, 60, 3470–3476 CAS.
- G. R. Mundy and J. W. Poser, Calcif. Tissue Int., 1983, 35, 164–168 CrossRef CAS.
- K. Sato, K. Mawatari and T. Kitamori, Lab Chip, 2008, 8, 1992–1998 RSC.
- K. Sato, M. Tokeshi, H. Kimura and T. Kitamori, Anal. Chem., 2001, 73, 1213–1218 CrossRef CAS.
- K. Sato, M. Tokeshi, T. Odake, H. Kimura, T. Ooi, M. Nakao and T. Kitamori, Anal. Chem., 2000, 72, 1144–1147 CrossRef CAS.
- K. Sato, M. Yamanaka, T. Hagino, M. Tokeshi, H. Kimura and T. Kitamori, Lab Chip, 2004, 4, 570–575 RSC.
- K. Sato, M. Yamanaka, H. Takahashi, M. Tokeshi, H. Kimura and T. Kitamori, Electrophoresis, 2002, 23, 734–739 CrossRef CAS.
- T. Sakata, M. Ihara, I. Makino, Y. Miyahara and H. Ueda, Anal. Chem., 2009, 81, 7532–7537 CrossRef CAS.
- T. Ohashi, K. Mawatari, K. Sato, M. Tokeshi and T. Kitamori, Lab Chip, 2009, 9, 991–995 RSC.
- R. D. Soltis, D. Hasz, M. J. Morris and I. D. Wilson, Immunology, 1979, 36, 37–45 CAS.
- P. H. van der Meide, M. C. de Labie, J. A. Wubben and A. H. Borman, J. Immunoassay Immunochem., 1991, 12, 65–82 CrossRef CAS.
- P. Garnero, M. Grimaux, B. Demiaux, C. Preaudat, P. Seguin and P. D. Delmas, J. Bone Miner. Res., 1992, 7, 1389–1398 CrossRef CAS.
- C. M. Gundberg and R. S. Weinstein, J. Clin. Invest., 1986, 77, 1762–1767 CrossRef CAS.
- C. Rosenquist, P. Qvist, N. Bjarnason and C. Christiansen, Clin. Chem., 1995, 41, 1439–1445 CAS.
- A. K. Taylor, S. Linkhart, S. Mohan, R. A. Christenson, F. R. Singer and D. J. Baylink, J. Clin. Endocrinol. Metab., 1990, 70, 467–472 CrossRef CAS.
- S. Y. Hsieh, R. K. Chen, Y. H. Pan and H. L. Lee, Proteomics, 2006, 6, 3189–3198 CrossRef CAS.
- J. Pfeilschifter, J. Erdmann, W. Schmidt, A. Naumann, H. W. Minne and R. Ziegler, Endocrinology, 1990, 126, 703–711 CrossRef CAS.
- H. Tammen, R. Hess, H. Rose, W. Wienen and M. Jost, Peptides, 2008, 29, 2188–2195 CrossRef CAS.
- E. M. Diaz Diego, R. Guerrero and C. de la Piedra, Clin. Chem., 1994, 40, 2071–2077 CAS.
- D. Essassi, R. Zini and J. P. Tillement, Prog. Clin. Biol. Res., 1989, 300, 423–426 Search PubMed.
- D. M. Togashi and A. G. Ryder, J. Fluoresc., 2008, 18, 519–526 CrossRef CAS.
- R. Arai, H. Ueda, K. Tsumoto, W. C. Mahoney, I. Kumagai and T. Nagamune, Protein Eng., Des. Sel., 2000, 13, 369–376 CrossRef CAS.
- L. H. Brinckerhoff, V. V. Kalashnikov, L. W. Thompson, G. V. Yamshchikov, R. A. Pierce, H. S. Galavotti, V. H. Engelhard and C. L. Slingluff, Jr., Int. J. Cancer, 1999, 83, 326–334 CrossRef CAS.
- S. Lien and H. B. Lowman, Trends Biotechnol., 2003, 21, 556–562 CrossRef CAS.
- A. Oki, H. Ogawa, M. Nagai, S. Shinbashi, M. Takai, A. Yokogawa and Y. Horiike, Mater. Sci. Eng., C, 2004, 24, 837–843 CrossRef.
Footnotes |
| † Electronic supplementary information (ESI) available: Additional experimental details and figures. See DOI: 10.1039/b915516c |
| ‡ Current address: Department of Bioscience and Biotechnology, Faculty of Agriculture, Shinshu University, 8304 Minawi-minowa, Kami-ina, Nagano 339–4598, Japan. |
| This journal is © The Royal Society of Chemistry 2010 |