Advances and challenges in the identification of volatiles that mediate interactions among plants and arthropods
Marco
D'Alessandro
and
Ted C. J.
Turlings
*
University of Neuchâtel, Institute of Zoology, Laboratory of Evolutionary Entomology, Case Postale 2, CH-2007 Neuchâtel, Switzerland. E-mail: ted.turlings@unine.ch; Fax: +41 32 718 30 01; Tel: +41 32 718 31 85
First published on 2nd December 2005
Abstract
The relatively new research field of Chemical Ecology has, over the last two decades, revealed an important role of plant-produced volatile organic compounds (VOCs) in mediating interactions between plants and other organisms. Of particular interest are the volatile blends that plants actively emit in response to herbivore damage. Various efforts are underway to pinpoint the bioactive compounds in these complex blends, but this has proven to be exceedingly difficult. Here we give a short overview on the role of herbivore-induced plant volatiles in interactions between plants and other organisms and we review methods that are currently employed to collect and identify key volatile compounds mediating these interactions. Our perspective on future directions of this fascinating research field places special emphasis on the need for an interdisciplinary approach. Joint efforts by chemists and biologists should not only facilitate the elucidation of crucial compounds, but can also be expected to lead to an exploitation of this knowledge, whereby ecological interactions may be chemically manipulated in order to protect crops and the environment.
 Marco D'Alessandro  Ted Turlings | Marco D’Alessandro is currently finishing his PhD thesis in Chemical Ecology at the University of Neuchâtel under the supervision of his co-author Ted Turlings. During his research Marco has developed and employed different methods to identify key volatile organic compounds that attract parasitic wasps. He previously studied at the University of Zurich and at the Swiss Federal Institute of Technology Zurich, and worked as a research assistant at the USDA research station, Tifton, Georgia, USA and at the Swiss federal research station in Reckenholz (FAL), where he studied physiological and ecological aspects of the interactions between plants and insects. Currently he is also working on belowground interactions between plants and microbes and on the effects of these interactions on other organisms aboveground. |
Introduction
Until recently, it was not common knowledge that plants have a way to express themselves. They mainly do so by emitting odours and the chemical composition and intensity of these odours can carry information on the plants' physiological status and on the stresses that they are being subjected to. In fact, plants emit an enormous spectrum of volatile organic compounds (VOCs). At present, more than 1000 of these low molecular weight organic compounds are known, ranging from terpenoids, fatty acid derivatives, benzenoids and phenylpropanoids, C5-branched compounds and various nitrogen and sulfur containing compounds.1 Some of these VOCs are constitutively emitted by undamaged, healthy plants, but herbivore damage commonly induces plants to emit much larger amounts and may even cause several VOCs to be synthesised de novo2–4 (Fig. 1). These herbivore-induced plant volatiles (HIPVs) are known to be emitted by various parts of the plant, including leaves4–7 and flower buds8 as well as roots.9 Not only feeding by a herbivorous insect induces the release of HIPVs; even deposition of eggs by herbivorous insects has been shown to induce the plant to emit HIPVs.10–12 In addition, HIPV emission is not limited to the site of damage but occurs systemically throughout the plant, also from undamaged leaves.13–17 Many insects are well aware of this fragrant lingo of plants and have evolved ways to exploit it (Fig. 2). Recently, researchers have also become aware of this and now HIPV emissions have been the subject of an increasing number of investigations that have revealed an important role of HIPVs in communication and self-protection. Although molecular and genetic approaches are now frequently applied to reveal the intricacies of plant-mediated interactions, the chemical analyses of HIPVs remain an integral part of virtually all studies.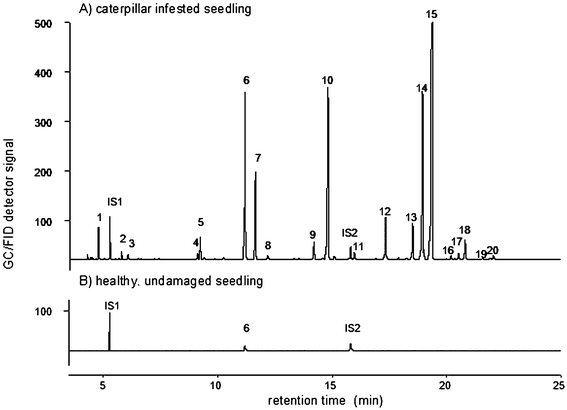 | ||
| Fig. 1 Example of GC-FID chromatograms obtained with the collected volatiles of 10-day old maize seedlings, (A) seedlings infested with Spodoptera littoralis larvae (B) undamaged seedlings. Further details on material and methods are given by D'Alessandro and Turlings.103 Compounds are: 1 = (Z)-3-hexenal, 2 = (E)-2-hexenal, 3 = (E)-3-hexen-1-ol, 4 = β-myrcene, 5 = (Z)-3-hexen-1-ol acetate, 6 = linalool, 7 = (3E)-4,8-dimethyl-1,3,7-nonatriene, 8 = benzyl acetate, 9 = phenethyl acetate, 10 = indole, 11 = methyl anthranilate, 12 = geranyl acetate, 13 = (E)-β-caryophyllene, 14 = (E)-α-bergamotene, 15 = (E)-β-farnesene, 16 = unknown sesquiterpenoid, 17 = unknown sesquiterpenoid, 18 = β-sesquiphellandrene, 19 = (E)-nerolidol, 20 = (3E,7E)-4,8,12-trimethyl-1,3,7,11-tridecatetraene. IS1 and IS2 = internal standards. | ||
 | ||
| Fig. 2 A female of the parasitic wasp Cotesia marginiventris lays an egg in a caterpillar. The wasp found its victim with the use of volatiles emitted by the plant in response to being damaged by the caterpillar. Picture by Matthias Held. | ||
Below we provide a short overview of what is known about the physiological and ecological role of HIPVs in interactions between plants and other organisms (for recent reviews see ref. 18–21), we further review methods commonly used by biologists to collect and analyse HIPVs and highlight the challenges that remain to be tackled in this area of research. The focus is on techniques used to study the importance of such HIPVs for the attraction of arthropods aboveground, and we propose some methods that could be useful in future studies, including the analysis of HIPVs belowground. Further details on these techniques and on techniques used to analyse plant VOCs in general have been described and reviewed elsewhere (e.g.ref. 22–26). Finally, we discuss and stress the importance of testing the biological relevance of HIPVs with appropriate behavioural bioassays and we propose that the remarkable sensitivity of insect chemoreceptors to VOCs make them exploitable as tools for future research on plant odours.
Physiology, ecology and application of biologically relevant HIPVs
HIPV-blends are complex and consist of a variety of divergent VOCs, including alkanes, alkenes, aldehydes, alcohols, ketones, ethers, esters and carboxylic acids. It is unlikely that every VOC emitted by plants serves an ecological or physiological role27 and there is still disagreement about the evolutionary history and function of plant VOCs.20,27–33Undoubtedly, HIPVs play a central role in mediating interactions between plants and herbivores, between herbivores and their natural enemies, between plants and microorganisms, but also between plants themselves.21,34,35 For example, laboratory studies have shown that HIPVs deter oviposition by arthropod herbivores,36,37 attract natural enemies of these herbivores4,5,9 and even induce defence genes and VOC emission in neighbouring plants38–40 or prime these plants to respond faster to future herbivore attack.41 In field studies, HIPVs have been shown to have the potential to reduce the damage by pests of various plants including crop plants.9,42–45 This opens the possibility of exploiting HIPVs for the development of novel strategies in crop protection. Indeed, an experiment whereby a hop yard was baited with methyl salicylate (MeSA) resulted in a significant increase in the numbers of beneficial predatory insects and in a dramatic reduction in spider mite numbers, the major arthropod pest of hops.46 Recent advances in the biochemistry and molecular genetics of terpene biosynthesis in various plant families47–59 should enable breeders to engineer plants to emit odours that more effectively attract the enemies of herbivores and thus reduce herbivory.60,61 That this is a realistic possibility was demonstrated with Arabidopsis thaliana plants that were transformed with sesquiterpenoid synthase genes, making them release two additional terpenoid compounds that were attractive to the predatory mites, Phytoseiulus persimili.62 Further field studies with synthetic HIPVs and genetically modified plants releasing such HIPVs will provide additional indications as to whether such manipulations could be included in agricultural practices to protect crops and the environment.
Various studies have provided detailed knowledge on the physiological and molecular basis of plant volatile synthesis and indirect defence responses in plants (reviewed by ref. 1,18,19). Still, our understanding of how the biosynthetic pathways are induced and interact with each other is rudimentary, probably because the induction and emission of HIPVs depends on the interaction between biotic factors, such as plant hormones,19,63–68 herbivore-derived elicitors,69–74 associated microorganisms75,76 and abiotic factors, such as wounding,77–80 O3 and CO2 concentration,81–84 temperature and light,85–87 UV-radiation88 and many other factors.89,90 Applying several stresses simultaneously to get a better understanding of how different pathways interact, will be one of the challenges in future studies on the physiology of plant volatiles.
There is a current boom in interest among ecologists to include the belowground interactions in studies on plants and associated organisms,91–94 and VOCs have been found also to be involved in tritrophic signalling belowground.16,95–97 Chen et al.98 characterized a root-specific Arabidopsis terpene synthase responsible for the formation of the volatile monoterpene 1,8-cineole, which is possibly involved in belowground interactions. Recently, Rasmann et al.9 identified a sesquiterpene, (E)-β-caryophyllene, as a belowground herbivore-induced volatile signal that attracts entomopathogenic nematodes, which infect and kill larvae of the corn rootworm, Diabrotica virgifera virgifera, a voracious pest of maize. Another challenge in future studies will be to determine how such belowground interactions might influence aboveground volatile emissions and vice versa.
Collection and analysis of HIPVs
Sampling aboveground
The literature on HIPVs released by vegetative plant parts is vast and is continuously growing. In most cases it is unknown which compounds have biological activity and therefore biologists mostly try to sample and analyse the full range of HIPVs. This is usually achieved by collecting VOCs in the headspace of herbivore-induced plants that are enclosed in collection chambers using an adsorbing material with a relatively broad spectrum of adsorption (e.g.ref. 4,99). Subsequently the collected volatiles can be analysed by gas-chromatography (GC) and mass-spectrometry (MS) or a combination of both (GC/MS).Fig. 3 shows the frequency with which the most commonly used sampling methods have been applied in the research of HIPVs during the last 10 years. Although this literature survey probably includes only part of all studies on herbivore-induced volatiles, it is clear that collection on adsorbents followed by desorption with solvents has been the most commonly used method. One advantage associated with solvent desorption is that it results in liquid samples that can be stored, which then can be used several times. This can be very useful for repeated analyses in GC, GC/MS and GC-EAD (gas-chromatography and electroantennogram-detection),100 for further fractionation,101 for peak enhancement coinjections102 and for bioassays.103–105 Adsorbents and solvents used in these studies vary, but the porous polymer Super Q106 (80–100 mesh, Alltech, Deerfield, Illinois, USA) and the solvent dichloromethane are widely used by many groups. Although this adsorbent–solvent combination has proven highly effective in HIPV adsorption and desorption, Harper107 argues that more than one adsorbent might be required to cover different classes of compounds, such as amines, aldehydes and aromatic hydrocarbons. There exist a wide range of adsorbent materials that can be used for this purpose, including activated charcoal, Anasorb 747, carboxens, silica gel, carbon molecular sieves and porous polymers such as Tenax, Chromosorb.107,108 Choosing a combination of appropriate adsorbents may increase the number of compounds found in HIPV-blends. Still, only a small volume of the desorbed compounds can be injected into the GC, leaving minor compounds undetected, and the solvent peak can mask some highly volatile compounds. These problems can be avoided by using a desorption method without solvent, like thermal desorption, or solid-phase microextraction (SPME).22,24,108–113
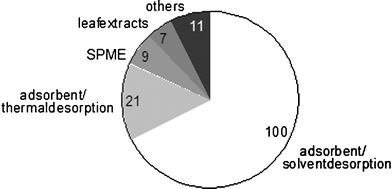 | ||
| Fig. 3 The most commonly used HIPV collection methods during the last ten years. The numbers represent the number of studies found with a search on the “Web of Science” by entering the terms: “induc* and volatil* and plant*” from 1995–2004. | ||
The SPME technique employs a small fibre coated with an adsorbing material and has found widespread application in many fields, mostly for qualitative and semi-quantitative analyses in environmental and food analyses, but also quantitatively in analytical chemistry.22 SPME has also gained increasing popularity in analyses of living biological samples, including plant VOCs111 (Fig. 4). It has a number of advantages, like simplifying sample preparation, increasing reliability, selectivity and sensitivity and reducing analysis time. Various coating materials for the SPME fibres are available,22,24,111 and choosing the appropriate fibre and sampling conditions are crucial for this technique to be effective. Still, the biggest limitation of SPME in studies on HIPVs is that the sample is lost after analysis and cannot be used for further analyses, GC-EAD, bioassays or fractionation.
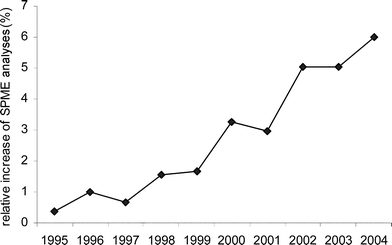 | ||
| Fig. 4 Relative increase of the use of SPME to collect plant volatiles. The graph depicts the percentage of all surveyed plant volatile collection studies that employed the SPME technique. This information was obtained with a search on the “Web of Science” using the terms: “SPME and plant* and volatil*” and “volatil* and plant*”. | ||
Higher selectivity and sensitivity could also be obtained by using techniques specifically adapted for compounds with known functional groups. For example, damaged Allium plants produce and release sulfur containing VOCs, presumably to prevent insect herbivory.114 To analyse true onion volatiles, Arnault et al.115 used a combination of SPME-GC/MS and solvent extraction followed by GC/MS analyses specifically developed for these compounds. Other compounds might be trapped by using microchemical techniques such as derivatization and degradation methods.116,117 Such techniques have played crucial roles in the determination of the structure of insect pheromones118,119 and are widely used in ambient air analyses.110,120–122 In pheromone studies, micro-reactions have three main uses; derivatization to aid chromatographic properties, functional group modification to help with MS structure determination, and assignment of absolute configuration to chiral centres.118 In analyses of HIPVs these techniques are rarely used, despite the fact that such techniques seem ideal for sampling trace amounts or highly reactive VOCs, which are difficult to sample with any other technique. For example, aldehydes can be sampled using a 2,4-dinitrophenylhydrazine (DNPH)-coated filter, and amines with a filter coated with 1-naphthyl isothiocyanate.122 Pre-treated filters are available from various companies (e.g. Supelco, SKC) and can easily be adapted to selectively adsorb HIPVs with a specific functional group. In our research, we used DNPH-coated silica (Supelco) to selectively adsorb 3-(Z)-hexenal and 2-(E)-hexenal, the two major aldehydes emitted by maize plants under caterpillar attack (see below and Fig. 5).
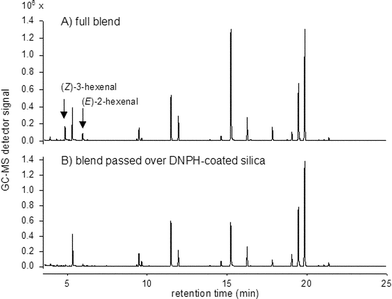 | ||
| Fig. 5 GC-MS chromatograms obtained with the collected volatiles of caterpillar-infested maize seedlings. (A) Complete caterpillar-induced maize blend adsorbed on Super-Q filters and desorbed with dichloromethane. (B) Volatiles from the same blend after it was passed over a DNPH-coated silica filter, which selectively adsorbs carbonyl compounds. This breakthrough blend of HIPVs was collected and analysed as in (A). Further details on material and methods are given by D'Alessandro and Turlings.103 | ||
An additional problem faced in analyses of HIPVs is that the release of some volatiles is highly dynamic, depending on the time after the plant is damaged,3,123 but also on the time of day36,124 and even season.89,125,126 Insects can perceive minor changes in odour quantity and quality. Therefore, a good understanding of the kinetics of formation and release of HIPV is highly desired. Specifically designed and automated headspace collection systems and GC-analyses allow time dependent collection of volatile plant compounds,3,99,127 and novel techniques even allow real time analyses of the emission of VOCs. For example, proton-transfer-reaction mass spectrometry (PTR-MS)128–130 and portable artificial noses131 permit fast sampling and real time (one measurement per second) analyses of plant volatiles and thus provide new insights into the kinetics of plant volatile releases. These techniques have the limitation that they do not distinguish between different VOCs with the same mass. Yet, Penuelas, Filella and co-workers132 nicely show that the PTR-MS technique can be used to monitor small volatiles, like methanol released by Succisa pratensis leaves infested with caterpillars of Euphydryas aurinia. Such highly volatile compounds are likely to break through from sorbent tubes or are masked by the solvent peak in liquid sample GC-analyses and are seldom reported in HIPV studies using sorbent trapping.
To fully understand how HIPVs are produced and released, more comprehensive metabolomic approaches are needed. Schmelz et al.66,133 have introduced an elegant technique based on vapour phase extraction and analysis by chemical ionization-GC/MS that simultaneously analyses phytohormones, phytotoxins, and VOCs in plants. Derivatization techniques are widely employed in metabolic profiling (reviewed in ref. 134) and combining metabolic and transcript analyses might provide new insights into which genes are involved in the production of HIPVs.135
Deciding on which technique to use remains difficult, as several comparisons of different sampling techniques can result in quantitative and qualitative differences.136–138 We suggest that the most reliable results in HIPV-analyses can be expected from the use of a combination of different approaches, including techniques specifically developed and calibrated for quantification of compounds with known functional groups. Knowing the exact relative quantities can be essential because often ratios among different compounds can be important in determining the attractiveness of odour blends to insects.139–141
Sampling belowground
Studies on belowground interactions bring new challenges to develop collection and analytical methods. So far, research on belowground allelochemicals mainly used extracts of root exudates,142–144 but other methods to analyse VOCs from soil samples are also available.130,145–150 As in aerial VOC analyses, comparing various sampling methods can lead to different quantities of VOCs.148 It is therefore pertinent to include more than one method in the belowground collections. The soil is much more heterogeneous than the air and large differences in soil properties and VOC concentrations can occur over extremely short distances, which must be taken into account during sampling.151 Biotic and abiotic factors that are known to affect the quantity and quality of HIPV-blends aboveground are likely to significantly affect belowground volatiles as well. Therefore, a major challenge for the analyses of belowground HIPVs will not only be to choose appropriate sampling and analysis tools, but also to obtain detailed knowledge on their kinetics, concentrations and distribution in the soil. Probably a good way to start is to analyse HIPVs in relatively well-defined, homogeneous soil types and then to repeat the measurements in more complex soils as well as in field experiments. For example, to determine the role of insect-induced (E)-β-caryophyllene from maize roots as an attractant for nematodes, an initial comparison was made between insect-damaged and undamaged maize roots, using pulverized roots. In subsequent tests, it was confirmed that (E)-β-caryophyllene rapidly diffuses through sand and attracts the nematodes, and finally the attractiveness of this sesquiterpene was tested in field experiments under natural conditions.9Bioassays and bioassay-linked HIPV analyses
Using HIPVs as novel tools in crop protection implies not only a need for detailed knowledge about the biosynthesis and release of plant VOCs, but also about the perception and exploitation of these chemical signals by the animals. The identification of the specific, behaviourally active compounds within a complex blend has proven to be difficult. Commonly, such compounds are identified with a combination of behavioural studies and chemical analyses of the tested odour blends.36,103,105,152–156 Another frequently practised method is gas chromatography combined with electroantennogram detection (GC-EAD) or with single cell recordings (GC-SCR),157 whereby the end of the GC column is split and one part of the effluent passes into the normal flame ionization detector (FID) and the other part is passed over an antenna or a single olfactory sensillum of the study insect. With the use of electrodes and amplifiers the responses of the antennal receptors can be measured and registered. Correlating these responses with the chromatogram obtained with FID detection reveals which compounds are perceived by the antennae.100,158–162 Advances in this methodology that allow recordings to be made inside the central nervous system in combination with the novel technique of in vivo calcium-imaging have helped neurobiologists to start to understand how plant VOCs are detected and processed by insects.163–165While methods such as GC-EAD or GC-SCR might give information on whether a compound is perceived by the olfactory system of an animal,157 only behavioural tests will show if the animals are indeed attracted or repelled by a particular compound. So far, methods for testing the attractiveness of odour blends to arthropods consisted mainly of dynamic air bioassays using olfactometers,166,167 wind tunnels,4,168,169 or static air bioassays.170,171 In these experiments arthropods are released at a defined distance from an odour source and their behaviour and attraction is recorded. In most cases chemical information on the odour sources had to be obtained in separate analytical studies. However, natural HIPV-blends exhibit not only high interspecific and intraspecific variability,172–174 but quantity and quality of the compounds depends also on the degree and time after infestation.3,175 To account for such variation, newer approaches attempt to standardize odour blends and to closely link chemical profiles with the insect behaviour.175,176 For example, we have developed an olfactometer in which six odour sources can be tested simultaneously for their relative attractiveness, and during the assays part of each odour blend can be trapped for further analyses.175 This device not only provides a direct link between the chemical profiles and the insect behaviour, but it also allows direct comparison of the attractiveness of a multitude of odour sources with multivariate statistical approaches, including principal component analysis (PCA) and redundancy analysis (RDA). Multivariate statistical analyses are widely applied in analyses of food volatiles177–179 and might also provide additional information in the analyses of HIPVs172,180 and reveal key compounds that attract arthropods.
Another way of studying the importance of individual VOCs within a complex blend is to compare the attractiveness of volatile blends differing in only a few known compounds. These blends can be obtained by using different chemical elicitors and inhibitors181–183 or by silencing genes involved in indirect defences.184,185 Adding back missing VOCs to incomplete blends is a sound way to study the importance of individual compounds. For example the predatory mite Phytoseiulus persimilis prefers a methyl salicylate (MeSA)-containing volatile blend, induced by the spider mite Tetranychus urticae to a similar but MeSA-free blend, induced by jasmonic acid.152 Adding synthetic MeSA to the MeSA-free blend significantly increased attraction to this odour, suggesting an important role for MeSA as a foraging cue for this predatory arthropod.152,186
We used an additional approach to obtain blends differing in only a few known compounds,103 for which we modified the six-arm olfactometer described by Turlings et al.175 in order to install adsorbing filter tubes between the odour source vessels and the arms of the olfactometers. By passing the induced volatile blend over these filter tubes, some compounds can be retained by the adsorbent material, while others break through and can be tested for attraction (Fig. 5). Furthermore, synthetic compounds can readily be added to the HIPV-blend and thus we are able to evaluate the importance of individual or groups of specific compounds. This novel technique has revealed an unexpected importance of minor compounds for the attraction of parasitic wasps to the complex HIPV-blends, whereas several of the dominating compounds appear to be only important after the wasps learn to associate them with the presence of hosts.103 These experiments once more showed that responses towards HIPVs are not fixed. Indeed, it is well known that responses of insects to VOCs are highly plastic, depending on the physiological state of the animals and on previous experiences with odour sources.187–189 Different forms of learning may modify the perception of chemical compounds and the response of insects towards odour sources.190–193 This flexibility in the insects' responses, as well as the great variability in the odours produced172,174,194–196 duly complicates the elucidation of key attractants, and learning experiments should be included in behavioural assays.
Conclusions and prospects
HIPVs play crucial roles in the interactions among various organisms. A good understanding of the key compounds involved in these interactions will not only provide important fundamental ecological knowledge, but should also allow us to manipulate certain of these interactions to our advantage, especially in agriculture. The complexity and variability of volatile blends emitted by herbivore-infested plants have proven to greatly complicate the identification of the principal compounds mediating interactions between the emitting plants and associated organisms. Renewed efforts should be made to integrate and adapt the latest techniques in analytical chemistry for this purpose. Choosing appropriate methods for sampling and analysing HIPVs is crucial. It might be possible to directly exploit insects' olfaction and behaviour to develop new selective and sensitive biosensors.197–201 Insect chemoreception is known to be more sensitive and specific than any chemical detection system available today.189 For example, in the moth Spodoptera littoralis, a change in heartbeat frequency can be triggered when fewer than six molecules of a key pheromone component hit the antennae of the insect.202 Thus, the insects themselves might be most capable of informing us on which are the key substances that they use in their behaviour. Including well-designed electro-physiological and behavioural bioassays can provide essential additional information about bioactive compounds that cannot or can barely be detected in chemical analyses. In future studies biosensing techniques including electronic noses203–206 might add to classical chemical analyses of HIPVs. If biologists, chemists and engineers join forces in this effort they will undoubtedly be able to fully unravel the fascinating world of chemically-mediated interactions between plants and their biotic environment. Such insight will then open the way to manipulate the interactions for our own benefit.Acknowledgements
We greatly benefited from inspiring discussions with all colleagues in the group of Martine Rahier at the University of Neuchâtel and we are particularly grateful to Matthias Held and Russell Naisbit for their comments on an earlier version of this manuscript. We wish to acknowledge the financial support for our work by the Swiss National Science Foundation (grant 31-058865.99) and the Swiss National Centre of Competence in Research “Plant Survival”.References
- N. Dudareva, E. Pichersky and J. Gershenzon, Plant Physiol., 2004, 135, 1893–1902 CrossRef CAS.
- P. W. Paré and J. H. Tumlinson, Plant Physiol., 1997, 114, 1161–1167 CAS.
- T. C. J. Turlings, U. B. Lengwiler, M. L. Bernasconi and D. Wechsler, Planta, 1998, 207, 146–152 CrossRef CAS.
- T. C. J. Turlings, J. H. Tumlinson and W. J. Lewis, Science, 1990, 250, 1251–1253 CrossRef CAS.
- M. Dicke and M. W. Sabelis, Neth. J. Zool., 1988, 38, 148–165 Search PubMed.
- R. M. P. Van Poecke, M. A. Posthumus and M. Dicke, J. Chem. Ecol., 2001, 27, 1911–1928 CrossRef CAS.
- T. Maeda and J. Takabayashi, Appl. Entomol. Zool., 2001, 36, 47–52 Search PubMed.
- U. S. R. Röse and J. H. Tumlinson, Planta, 2004, 218, 824–832 CrossRef.
- S. Rasmann, T. G. Köllner, J. Degenhardt, I. Hiltpold, S. Töpfer, U. Kuhlmann, J. Gershenzon and T. C. J. Turlings, Nature, 2005, 434, 732–737 CrossRef CAS.
- T. Meiners and M. Hilker, Oecologia, 1997, 112, 87–93 CrossRef.
- R. Schröder, M. Forstreuter and M. Hilker, Plant Physiol., 2005, 138, 470–477 CrossRef.
- R. Mumm, K. Schrank, R. Wegener, S. Schulz and M. Hilker, J. Chem. Ecol., 2003, 29, 1235–1252 CrossRef CAS.
- T. C. J. Turlings and J. H. Tumlinson, Proc. Natl. Acad. Sci. USA, 1992, 89, 8399–8402 CAS.
- U. S. R. Röse, W. J. Lewis and J. H. Tumlinson, J. Chem. Ecol., 1998, 24, 303–319 CrossRef CAS.
- L. Mattiacci, B. A. Rocca, N. Scascighini, M. D'Alessandro, A. Hern and S. Dorn, J. Chem. Ecol., 2001, 27, 2233–2252 CrossRef CAS.
- N. Neveu, J. Grandgirard, J. P. Nenon and A. M. Cortesero, J. Chem. Ecol., 2002, 28, 1717–1732 CrossRef CAS.
- E. Guerrieri, G. M. Poppy, W. Powell, E. Tremblay and F. Pennacchio, J. Chem. Ecol., 1999, 25, 1247–1261 CrossRef CAS.
- G. Arimura, C. Kost and W. Boland, Biochim. Biophys. Acta, Mol. Cell Biol. Lipids, 2005, 1734, 91–111 CAS.
- R. M. P. van Poecke and M. Dicke, Plant Biol., 2004, 6, 387–401 CrossRef CAS.
- J. K. Holopainen, Trends Plant Sci., 2004, 9, 529–533 CrossRef CAS.
- T. C. J. Turlings and F. Wäckers, in Advances in Insect Chemical Ecology, ed. R. T. Cardé and J. G. Millar, Cambridge University Press, Cambridge, UK, 2004 Search PubMed.
- G. Vas and K. Vekey, J. Mass Spectrom., 2004, 39, 233–254 CrossRef CAS.
- R. M. Smith, J. Chromatogr., A, 2003, 1000, 3–27 Search PubMed.
- E. Baltussen, C. A. Cramers and P. J. F. Sandra, Anal. Bioanal. Chem., 2002, 373, 3–22 CrossRef CAS.
- P. Ciccioli, E. Brancaleoni, M. Frattoni and C. Maris, in Environmental Monitoring Handbook, ed. F. R. Burden, I. McKelvie, U. Forstner and A. Guenther, McGrawHill, New York, 2002, ch. 21 Search PubMed.
- J. G. Millar and K. F. Haynes, Methods in Chemical Ecology, Chapman & Hall, New York, 1998 Search PubMed.
- J. Penuelas and J. Llusia, Trends Ecol. Evol., 2004, 19, 402–404 Search PubMed.
- D. R. Gang, Annu. Rev. Plant Biol., 2005, 56, 301–325 Search PubMed.
- S. M. Owen and J. Penuelas, Trends Plant Sci., 2005, 10, 420–426 CrossRef CAS.
- U. Niinemets, F. Loreto and M. Reichstein, Trends Plant Sci., 2004, 9, 180–186 CrossRef CAS.
- T. N. Rosenstiel, A. L. Ebbets, W. C. Khatri, R. Fall and R. K. Monson, Plant Biol., 2004, 6, 12–21 CrossRef CAS.
- A. Janssen, M. W. Sabelis and J. Bruin, Oikos, 2002, 97, 134–138 CrossRef.
- M. Dicke, in The Ecology and Evolution of Inducible Defenses, ed. R. Tollrian and C. D. Harvell, Princeton University Press, Princeton, 1999, pp. 62–88 Search PubMed.
- M. Dicke, R. M. P. van Poecke and J. G. de Boer, Basic Appl. Ecol., 2003, 4, 27–42 Search PubMed.
- I. T. Baldwin, A. Kessler and R. Halitschke, Curr. Opin. Plant Biol., 2002, 5, 351–354 CrossRef CAS.
- C. M. De Moraes, M. C. Mescher and J. H. Tumlinson, Nature, 2001, 410, 577–580 CrossRef CAS.
- P. J. Landolt, Entomol. Exp. Appl., 1993, 67, 79–85 Search PubMed.
- J. Ruther and S. Kleier, J. Chem. Ecol., 2005, 31, 2217–2222 CrossRef CAS.
- M. A. Farag, M. Fokar, H. A. Zhang, R. D. Allen and P. W. Paré, Planta, 2005, 220, 900–909 CrossRef CAS.
- G. Arimura, R. Ozawa, T. Shimoda, T. Nishioka, W. Boland and J. Takabyashi, Nature, 2000, 406, 512–515 CrossRef CAS.
- J. Engelberth, H. T. Alborn, E. A. Schmelz and J. H. Tumlinson, Proc. Natl. Acad. Sci. USA, 2004, 101, 1781–1785 CrossRef CAS.
- A. Kessler and I. T. Baldwin, Science, 2001, 291, 2141–2144 CrossRef CAS.
- J. S. Thaler, Nature, 1999, 399, 686–688 CrossRef CAS.
- M. L. Bernasconi, T. C. J. Turlings, L. Ambrosetti, P. Bassetti and S. Dorn, Entomol. Exp. Appl., 1998, 87, 133–142 Search PubMed.
- D. G. James, J. Chem. Ecol., 2003, 29, 1601–1609 CrossRef CAS.
- D. G. James and T. S. Price, J. Chem. Ecol., 2004, 30, 1613–1628 CrossRef CAS.
- S. Aubourg, A. Lecharny and J. Bohlmann, Mol. Genet. Genomics, 2002, 267, 730–745 CrossRef CAS.
- S. C. Trapp and R. B. Croteau, Genetics, 2001, 158, 811–832 Search PubMed.
- E. Fridman and E. Pichersky, Curr. Opin. Plant Biol., 2005, 8, 242–248 CrossRef CAS.
- D. Tholl, F. Chen, J. Petri, J. Gershenzon and E. Pichersky, Plant J., 2005, 42, 757–771 CrossRef CAS.
- Y. Jia, D. R. Xia and E. S. Louzada, J. Am. Soc. Hortic. Sci., 2005, 130, 454–458 Search PubMed.
- D. M. Martin, J. Faldt and J. Bohlmann, Plant Physiol., 2004, 135, 1908–1927 CrossRef CAS.
- A. Aharoni, A. P. Giri, S. Deuerlein, F. Griepink, W. J. de Kogel, F. W. A. Verstappen, H. A. Verhoeven, M. A. Jongsma, W. Schwab and H. J. Bouwmeester, Plant Cell, 2003, 15, 2866–2884 CrossRef CAS.
- J. Faldt, G. Arimura, J. Gershenzon, J. Takabayashi and J. Bohlmann, Planta, 2003, 216, 745–751 CAS.
- F. Chen, D. Tholl, J. C. D'Auria, A. Farooq, E. Pichersky and J. Gershenzon, Plant Cell, 2003, 15, 481–494 CrossRef CAS.
- J. Degenhardt and J. Gershenzon, Planta, 2000, 210, 815–822 CrossRef CAS.
- C. Schnee, T. G. Köllner, J. Gershenzon and J. Degenhardt, Plant Physiol., 2002, 130, 2049–2060 CrossRef CAS.
- T. G. Köllner, C. Schnee, J. Gershenzon and J. Degenhardt, Plant Cell, 2004, 16, 1115–1131 CrossRef.
- T. G. Köllner, C. Schnee, J. Gershenzon and J. Degenhardt, Phytochemistry, 2004, 65, 1895–1902 CrossRef CAS.
- J. Degenhardt, J. Gershenzon, I. T. Baldwin and A. Kessler, Curr. Opin. Biotechnol., 2003, 14, 169–176 CrossRef CAS.
- N. Dudareva and F. Negre, Curr. Opin. Plant Biol., 2005, 8, 113–118 CrossRef CAS.
- I. F. Kappers, A. Aharoni, T. van Herpen, L. L. P. Luckerhoff, M. Dicke and H. J. Bouwmeester, Science, 2005, 309, 2070–2072 CrossRef.
- G. L. de Bruxelles and M. R. Roberts, Crit. Rev. Plant Sci., 2001, 20, 487–521 CrossRef CAS.
- E. Rojo, R. Solano and J. J. Sanchez-Serrano, J. Plant Growth Regul., 2003, 22, 82–98 CrossRef CAS.
- J. S. Thaler, R. Karban, D. E. Ullman, K. Boege and R. M. Bostock, Oecologia, 2002, 131, 227–235 Search PubMed.
- E. A. Schmelz, J. Engelberth, H. T. Alborn, P. O'Donnell, M. Sammons, H. Toshima and J. H. Tumlinson, Proc. Natl. Acad. Sci. USA, 2003, 100, 10552–10557 CrossRef CAS.
- K. Ament, M. R. Kant, M. W. Sabelis, M. A. Haring and R. C. Schuurink, Plant Physiol., 2004, 135, 2025–2037 CrossRef CAS.
- E. E. Farmer, E. Almeras and V. Krishnamurthy, Curr. Opin. Plant Biol., 2003, 6, 372–378 CrossRef CAS.
- J. H. Tumlinson and C. G. Lait, Arch. Insect Biochem. Physiol., 2005, 58, 54–68 CrossRef CAS.
- D. Spiteller and W. Boland, J. Org. Chem., 2003, 68, 8743–8749 CrossRef CAS.
- R. Halitschke, U. Schittko, G. Pohnert, W. Boland and I. T. Baldwin, Plant Physiol., 2001, 125, 711–717 CrossRef CAS.
- L. Mattiacci, M. Dicke and M. A. Posthumus, Proc. Natl. Acad. Sci. USA, 1995, 92, 2036–2040.
- T. Alborn, T. C. J. Turlings, T. H. Jones, G. Stenhagen, J. H. Loughrin and J. H. Tumlinson, Science, 1997, 276, 945–949 CrossRef CAS.
- M. Merkx-Jacques and J. C. Bede, Phytoprotection, 2004, 85, 33–37 Search PubMed.
- J. Huang, Y. J. Cardoza, E. A. Schmelz, R. Raina, J. Engelberth and J. H. Tumlinson, Planta, 2003, 217, 767–775 CrossRef CAS.
- Y. J. Cardoza, H. T. Alborn and J. H. Tumlinson, J. Chem. Ecol., 2002, 28, 161–174 CrossRef CAS.
- R. Fall, T. Karl, A. Hansel, A. Jordan and W. Lindinger, J. Geophys. Res., [Atmos.], 1999, 104, 15963–15974 CrossRef CAS.
- G. A. Howe, J. Plant Growth Regul., 2004, 23, 223–237 CrossRef CAS.
- E. A. Schmelz, H. T. Alborn and J. H. Tumlinson, Planta, 2001, 214, 171–179 CrossRef CAS.
- A. Mithöfer, G. Wanner and W. Boland, Plant Physiol., 2005, 137, 1160–1168 CrossRef.
- J. Beauchamp, A. Wisthaler, A. Hansel, E. Kleist, M. Miebach, U. Niinemets, U. Schurr and J. Wildt, Plant, Cell Environ., 2005, 28, 1334–1343 CrossRef CAS.
- T. Vuorinen, A. M. Nerg and J. K. Holopainen, Environ. Pollut., 2004, 131, 305–311 CrossRef CAS.
- T. Vuorinen, A. M. Nerg, M. A. Ibrahim, G. V. P. Reddy and J. K. Holopainen, Plant Physiol., 2004, 135, 1984–1992 CrossRef CAS.
- R. Jasoni, C. Kane, C. Green, E. Peffley, D. Tissue, L. Thompson, P. Payton and P. W. Paré, Environ. Exp. Bot., 2004, 51, 273–280 CrossRef CAS.
- A. B. Guenther, P. R. Zimmerman, P. C. Harley, R. K. Monson and R. Fall, J. Geophys. Res., [Atmos.], 1993, 98, 12609–12617 Search PubMed.
- J. Takabayashi, M. Dicke and M. A. Posthumus, J. Chem. Ecol., 1994, 20(6), 1329–1354 CrossRef CAS.
- S. P. Gouinguené and T. C. J. Turlings, Plant Physiol., 2002, 129, 1296–1307 CrossRef CAS.
- C. B. Johnson, J. Kirby, G. Naxakis and S. Pearson, Phytochemistry, 1999, 51, 507–510 CrossRef CAS.
- J. Penuelas and J. Llusia, Biol. Plant., 2001, 44, 481–487 CrossRef CAS.
- J. Kesselmeier and M. Staudt, J. Atmos. Chem., 1999, 33, 23–88 CrossRef CAS.
- D. A. Wardle, R. D. Bardgett, J. N. Klironomos, H. Setala, W. H. van der Putten and D. H. Wall, Science, 2004, 304, 1629–1633 CrossRef CAS.
- T. M. Bezemer, R. Wagenaar, N. M. Van Dam and F. L. Wäckers, Oikos, 2003, 101, 555–562 CrossRef.
- N. M. van Dam, J. A. Harvey, F. L. Wäckers, T. M. Bezemer, W. H. van der Putten and L. E. M. Vet, Basic Appl. Ecol., 2003, 4, 63–77 Search PubMed.
- W. H. Van der Putten, L. E. M. Vet, J. A. Harvey and F. L. Wäckers, Trends Ecol. Evol., 2001, 16, 547–554 Search PubMed.
- N. S. Aratchige, I. Lesna and M. W. Sabelis, Exp. Appl. Acarol., 2004, 33, 21–30 CrossRef CAS.
- R. van Tol, A. T. C. van der Sommen, M. I. C. Boff, J. van Bezooijen, M. W. Sabelis and P. H. Smits, Ecol. Lett., 2001, 4, 292–294 CrossRef.
- M. I. C. Boff, F. C. Zoon and P. H. Smits, Entomol. Exp. Appl., 2001, 98, 329–337 Search PubMed.
- F. Chen, D. K. Ro, J. Petri, J. Gershenzon, J. Bohlmann, E. Pichersky and D. Tholl, Plant Physiol., 2004, 135, 1956–1966 CrossRef CAS.
- N. G. Agelopoulos, A. M. Hooper, S. P. Maniar, J. A. Pickett and L. J. Wadhams, J. Chem. Ecol., 1999, 25, 1411–1425 CrossRef CAS.
- S. Gouinguené, J. A. Pickett, L. J. Wadhams, M. A. Birkett and T. C. J. Turlings, J. Chem. Ecol., 2005, 31, 1023–1038 CrossRef CAS.
- T. C. J. Turlings and M. E. Fritzsche, in Insect-Plant Interactions and Induced Plant Defence, N. f. S. 223, ed. D.J. Chadwick and J. A. Goode, Wiley, Chichester, UK, 1999, pp. 21–38 Search PubMed.
- J. A. Pickett, in Chromatography and Isolation of Insect Hormones and Pheromones, ed. A. R. McCaffery and I. D. Wilson, Plenum Press, New York, 1990, pp. 299–309 Search PubMed.
- M. D'Alessandro and T. C. J. Turlings, Chem. Senses, 2005, 30, 739–753 Search PubMed.
- S. Udayagiri and R. L. Jones, J. Chem. Ecol., 1992, 18, 1841–1855 CrossRef CAS.
- T. C. J. Turlings, J. H. Tumlinson, R. R. Heath, A. T. Proveaux and R. E. Doolittle, J. Chem. Ecol., 1991, 17, 2235–2251 CAS.
- R. R. Heath and A. Manukian, J. Chem. Ecol., 1992, 18, 1209–1226 CrossRef CAS.
- M. Harper, J. Chromatogr., A, 2000, 885, 129–151 Search PubMed.
- K. Dettmer and W. Engewald, Anal. Bioanal. Chem., 2002, 373, 490–500 CrossRef CAS.
- T. Gorecki and J. Namiesnik, TrAC, Trends Anal. Chem., 2002, 21, 276–291 CrossRef CAS.
- J. A. Koziel and I. Novak, TrAC, Trends Anal. Chem., 2002, 21, 840–850 CrossRef CAS.
- F. Augusto and A. L. P. Valente, TrAC, Trends Anal. Chem., 2002, 21, 428–438 CrossRef CAS.
- G. A. Mills and V. Walker, J. Chromatogr., A, 2000, 902, 267–287 Search PubMed.
- L. Pillonel, J. O. Bossett and R. Tabacchi, Lebensm.-Wiss. Technol., 2002, 35, 1–14 CrossRef CAS.
- S. Dugravot, E. Thibout, A. Abo-Ghalia and J. Huignard, Entomol. Exp. Appl., 2004, 113, 173–179 Search PubMed.
- I. Arnault, N. Mondy, F. Cadoux and J. Auger, J. Chromatogr., A, 2000, 896, 117–124 Search PubMed.
- A. B. Attygalle, in Methods in Chemical Ecology, ed. J. G. Millar and K. F. Haynes, Chapman & Hall, New York, 1998, vol. 1 Search PubMed.
- E. E. Stashenko and J. R. Martinez, TrAC, Trends Anal. Chem., 2004, 23, 553–561 CrossRef CAS.
- G. R. Jones and N. J. Oldham, J. Chromatogr., A, 1999, 843, 199–236 Search PubMed.
- A. B. Attygalle and E. D. Morgan, Angew. Chem., Int. Ed. Engl., 1988, 27, 460–478 CrossRef.
- S. Uchiyama and S. Hasegawa, Atmos. Environ., 1999, 33, 1999–2005 CrossRef CAS.
- P. von Zweigbergk, R. Lindahl, A. Ostin, J. Ekman and J. O. Levin, J. Environ. Monit., 2002, 4, 663–666 RSC.
- J. O. Levin and R. Lindahl, Analyst, 1994, 119, 79–83 RSC.
- M. E. Hoballah and T. C. J. Turlings, J. Chem. Ecol., 2005, 31, 2003–2018 CrossRef CAS.
- J. H. Loughrin, A. Manukian, R. R. Heath, T. C. J. Turlings and J. H. Tumlinson, Proc. Natl. Acad. Sci. USA, 1994, 91, 11836–11840 CAS.
- A. Blaakmeer, J. B. F. Geervliet, J. J. A. Vanloon, M. A. Posthumus, T. A. Vanbeek and A. Degroot, Entomol. Exp. Appl., 1994, 73, 175–182 Search PubMed.
- A. Hern and S. Dorn, Phytochem. Anal., 2003, 14, 232–240 CrossRef CAS.
- R. R. Heath and A. Manukian, J. Chem. Ecol., 1994, 20, 593–608 CAS.
- J. A. De Gouw, C. J. Howard, T. G. Custer, B. M. Baker and R. Fall, Environ. Sci. Technol., 2000, 34, 2640–2648 CrossRef CAS.
- S. Hayward, A. Tani, S. M. Owen and C. N. Hewitt, Tree Physiol., 2004, 24, 721–728 Search PubMed.
- M. Steeghs, H. P. Bais, J. de Gouw, P. Goldan, W. Kuster, M. Northway, R. Fall and J. M. Vivanco, Plant Physiol., 2004, 135, 47–58 CrossRef CAS.
- M. Kunert, A. Biedermann, T. Koch and W. Boland, J. Sep. Sci., 2002, 25, 677–684 Search PubMed.
- J. Penuelas, I. Filella, C. Stefanescu and J. Llusia, New Phytol., 2005, 167, 851–857 Search PubMed.
- E. A. Schmelz, J. Engelberth, J. H. Tumlinson, A. Block and H. T. Alborn, Plant J., 2004, 39, 790–808 CrossRef CAS.
- J. M. Halket, D. Waterman, A. M. Przyborowska, R. K. P. Patel, P. D. Fraser and P. M. Bramley, J. Exp. Bot., 2005, 56, 219–243 Search PubMed.
- P. Mercke, I. F. Kappers, F. W. A. Verstappen, O. Vorst, M. Dicke and H. J. Bouwmeester, Plant Physiol., 2004, 135, 2012–2024 CrossRef CAS.
- J. Faldt, M. Eriksson, I. Valterova and A. K. Borg-Karlson, Z. Naturforsch., C: Biosci., 2000, 55, 180–188 CAS.
- N. G. Agelopoulos and J. A. Pickett, J. Chem. Ecol., 1998, 24, 1161–1172 CrossRef CAS.
- H. Rothweiler, P. A. Wager and C. Schlatter, Atmos. Environ., Part B, 1991, 25, 231–235 CrossRef.
- C. M. De Moraes, W. J. Lewis, P. W. Paré, H. T. Alborn and J. H. Tumlinson, Nature, 1998, 393, 570–573 CrossRef CAS.
- J. F. Tooker, W. A. Koenig and L. M. Hanks, Proc. Natl. Acad. Sci. USA, 2002, 99, 15486–15491 CrossRef CAS.
- T. J. A. Bruce, L. J. Wadhams and C. M. Woodcock, Trends Plant Sci., 2005, 10, 269–274 CrossRef CAS.
- C. Nguyen, Agronomie, 2003, 23, 375–396 Search PubMed.
- C. Bertin, X. H. Yang and L. A. Weston, Plant Soil, 2003, 256, 67–83 Search PubMed.
- Inderjit and S. O. Duke, Planta, 2003, 217, 529–539 CrossRef.
- J. S. Alvarado and C. Rose, Talanta, 2004, 62, 17–23 CrossRef CAS.
- T. E. Lewis, A. B. Crockett and R. L. Siegrist, Environ. Monit. Assess., 1994, 30, 213–246 CrossRef CAS.
- M. B. Wise, C. V. Thompson, R. Merriweather and M. R. Guerin, Field Anal. Chem. Technol., 1997, 1, 251–276 Search PubMed.
- M. W. Couch, C. J. Schmidt and S. C. Wasdo, Adv. Environ. Res., 2000, 4, 97–102 Search PubMed.
- M. Ojala, I. Mattila, V. Tarkiainen, T. Sarme, R. A. Ketola, A. Maattanen, R. Kostiainen and T. Kotiaho, Anal. Chem., 2001, 73, 3624–3631 CrossRef CAS.
- M. D. G. Meniconi, R. Parris and C. L. P. Thomas, Analyst, 2003, 128, 1232–1237 RSC.
- B. A. Schumacher and M. M. Minnich, Environ. Sci. Technol., 2000, 34, 3611–3616 CrossRef CAS.
- J. G. De Boer and M. Dicke, J. Chem. Ecol., 2004, 30, 255–271 CrossRef CAS.
- M. Dicke, T. A. van Beek, M. A. Posthumus, N. B. Dom, H. van Bokhoven and A. de Groot, J. Chem. Ecol., 1990, 16(2), 381–396 CrossRef CAS.
- M. E. F. Hoballah, C. Tamó and T. C. J. Turlings, J. Chem. Ecol., 2002, 28, 951–968 CrossRef CAS.
- M. A. Birkett, C. A. M. Campbell, K. Chamberlain, E. Guerrieri, A. J. Hick, J. L. Martin, M. Matthes, J. A. Napier, J. Pettersson, J. A. Pickett, G. M. Poppy, E. M. Pow, B. J. Pye, L. E. Smart, G. H. Wadhams, L. J. Wadhams and C. M. Woodcock, Proc. Natl. Acad. Sci. USA, 2000, 97, 9329–9334 CrossRef CAS.
- P. Scutareanu, B. Drukker, J. Bruin, M. A. Posthumus and M. W. Sabelis, J. Chem. Ecol., 1997, 23, 2241–2260 CrossRef CAS.
- A. Wibe, J. Insect Physiol., 2004, 50, 497–503 CrossRef CAS.
- H. A. Smid, J. J. A. van Loon, M. A. Posthumus and L. E. M. Vet, Chemoecology, 2002, 12, 169–176 Search PubMed.
- T. Rostelien, A. K. Borg-Karlson and H. Mustaparta, J. Comp. Physiol. A: Sens. Neural Behav. Physiol., 2000, 186, 833–847 Search PubMed.
- B. Weissbecker, J. J. A. Van Loon, M. A. Posthumus, H. J. Bouwmeester and M. Dicke, J. Chem. Ecol., 2000, 26, 1433–1445 CrossRef CAS.
- Y. J. Du, G. M. Poppy, W. Powell, J. A. Pickett, L. J. Wadhams and C. M. Woodcock, J. Chem. Ecol., 1998, 24, 1355–1368 CrossRef CAS.
- E. N. Barata, H. Mustaparta, J. A. Pickett, L. J. Wadhams and J. Araujo, J. Comp. Physiol. A: Neuroethol. Sens. Neural Behav. Physiol., 2002, 188, 121–133 Search PubMed.
- H. T. Skiri, C. G. Galizia and H. Mustaparta, Chem. Senses, 2004, 29, 253–267 Search PubMed.
- C. G. Galizia, S. Sachse and H. Mustaparta, J. Comp. Physiol. A: Sens. Neural Behav. Physiol., 2000, 186, 1049–1063 Search PubMed.
- H. Mustaparta, Entomol. Exp. Appl., 2002, 104, 1–13 Search PubMed.
- S. Steinberg, M. Dicke, L. E. M. Vet and R. Wanningen, Entomol. Exp. Appl., 1992, 63, 163–175 Search PubMed.
- L. E. M. Vet, J. C. van Lenteren, M. Heymans and E. Meelis, Physiol. Entomol., 1983, 8, 97–106 CrossRef.
- T. C. Baker and R. T. Cardé, in Techniques in Pheromone Research, ed. H. E. Hummel and T. A. Miller, Springer-Verlag, New York, 1984, pp. 45–73 Search PubMed.
- J. H. Visser, Entomol. Exp. Appl., 1976, 20, 275–288 Search PubMed.
- J. Ruther and J. L. M. Steidle, J. Chem. Ecol., 2000, 26, 1205–1217 CrossRef CAS.
- O. T. Jones and T. H. Coaker, Physiol. Entomol., 1977, 2, 189–197 CrossRef CAS.
- T. Degen, C. Dillmann, F. Marion-Poll and T. C. J. Turlings, Plant Physiol., 2004, 135, 1928–1938 CrossRef CAS.
- S. Gouinguené, T. Degen and T. C. J. Turlings, Chemoecology, 2001, 11, 9–16 Search PubMed.
- C. E. M. Van Den Boom, T. A. Van Beek, M. A. Posthumus, A. De Groot and M. Dicke, J. Chem. Ecol., 2004, 30, 69–89 CrossRef CAS.
- T. C. J. Turlings, A. C. Davison and C. Tamó, Physiol. Entomol., 2004, 29, 45–55 CrossRef.
- N. Scascighini, L. Mattiacci, M. D'Alessandro, A. Hern, A. S. Rott and S. Dorn, Chemoecology, 2005, 15, 97–104 Search PubMed.
- A. C. Noble and S. E. Ebeler, Food Rev. Int., 2002, 18, 1–21 Search PubMed.
- C. G. Zambonin, L. Balest, G. E. De Benedetto and F. Palmisano, Talanta, 2005, 66, 261–265 CrossRef CAS.
- S. Mildner-Szkudlarz, H. H. Jelen, R. Zawirska-Wojtasiak and E. Wasowicz, Food Chem., 2003, 83, 515–522 CrossRef CAS.
- R. Mumm, T. Tiemann, S. Schulz and M. Hilker, Phytochemistry, 2004, 65, 3221–3230 CrossRef CAS.
- M. Dicke, R. Gols, D. Ludeking and M. A. Posthumus, J. Chem. Ecol., 1999, 25, 1907–1922 CrossRef CAS.
- T. Koch, T. Krumm, V. Jung, J. Engelberth and W. Boland, Plant Physiol., 1999, 121, 153–162 CrossRef CAS.
- T. C. J. Turlings, H. T. Alborn, J. H. Loughrin and J. H. Tumlinson, J. Chem. Ecol., 2000, 26, 189–202 CrossRef CAS.
- A. Kessler, R. Halitschke and I. T. Baldwin, Science, 2004, 305, 665–668 CrossRef CAS.
- R. M. P. van Poecke and M. Dicke, Plant, Cell Environ., 2003, 26, 1541–1548 CrossRef CAS.
- J. G. de Boer and M. Dicke, Entomol. Exp. Appl., 2004, 110, 181–189 Search PubMed.
- K. Ueno and T. Ueno, J. Ethol., 2005, 23, 43–49 Search PubMed.
- L. E. M. Vet and M. Dicke, Annu. Rev. Entomol., 1992, 37, 141–172 CrossRef.
- M. Dicke, Entomol. Exp. Appl., 1999, 91, 131–142 Search PubMed.
- R. L. Davis, Neuron, 2004, 44, 31–48 CrossRef CAS.
- T. C. J. Turlings, F. L. Wäckers, L. E. M. Vet, W. J. Lewis and J. H. Tumlinson, in Insect Learning. Ecological and Evolutionary Perspectives., ed. D. R. Papaj and A. C. Lewis, Chapman & Hall, New York, 1993, pp. 51–78 Search PubMed.
- L. E. M. Vet, W. J. Lewis and R. T. Cardé, in Chemical Ecology of Insects, ed. R. T. Cardé and W. J. Bell, Chapman & Hall., New York, 1995, pp. 65–101 Search PubMed.
- D. R. Papaj and A. C. Lewis, Insect Learning, Chapman and Hall, New York, London, 1993 Search PubMed.
- O. E. Krips, P. E. L. Willems, R. Gols, M. A. Posthumus, G. Gort and M. Dicke, J. Chem. Ecol., 2001, 27, 1355–1372 CrossRef CAS.
- J. Takabayashi, M. Dicke and M. A. Posthumus, Chemoecology, 1991, 2, 1–6 CrossRef CAS.
- J. H. Loughrin, A. Manukian, R. R. Heath and J. H. Tumlinson, J. Chem. Ecol., 1995, 21, 1217–1227 CAS.
- K. C. Park and T. C. Baker, J. Insect Physiol., 2002, 48, 1139–1145 CrossRef CAS.
- S. Schutz, B. Weissbecker, U. T. Koch and H. E. Hummel, Biosens. Bioelectron., 1999, 14, 221–228 CrossRef.
- Y. Kuwana, S. Nagasawa, I. Shimoyama and R. Kanzaki, Biosens. Bioelectron., 1999, 14, 195–202 CrossRef CAS.
- G. C. Rains, J. K. Tomberlin, M. D'Alessandro and W. J. Lewis, Trans. ASAE, 2004, 47, 2145–2152 Search PubMed.
- M. Tertuliano, D. M. Olson, G. C. Rains and W. J. Lewis, Entomol. Exp. Appl., 2004, 110, 165–172 Search PubMed.
- A. M. Angioy, A. Desogus, I. T. Barbarossa, P. Anderson and B. S. Hansson, Chem. Senses, 2003, 28, 279–284 Search PubMed.
- D. James, S. M. Scott, Z. Ali and W. T. O'Hare, Microchim. Acta, 2005, 149, 1–17 Search PubMed.
- M. Trojanowicz and M. Szewczynska, TrAC, Trends Anal. Chem., 2005, 24, 92–106 CrossRef CAS.
- D. L. Garcia-Gonzalez and R. Aparicio, Grasas Aceites, 2002, 53, 96–114 Search PubMed.
- C. G. Pinto, M. E. F. Laespada, J. L. P. Pavon and B. M. Cordero, Quim. Anal., 2001, 20, 3–11 Search PubMed.
| This journal is © The Royal Society of Chemistry 2006 |