DOI: 10.1039/B315551J
(Paper)
Photochem. Photobiol. Sci., 2004, 3, 1-5
Environmental effects of ozone depletion and its interactions with climate change: Progress Report 2003
First published on 10th December 2003
Abstract
United Nations Environment Programme, Environmental Effects Assessment Panel†
Introduction
The measures needed for the protection of the Earth's ozone layer are decided regularly by the Parties to the Montreal Protocol, now consisting of 184 countries. The Parties are advised on knowledge relevant to this task by three panels of experts: the Scientific, Environmental Effects, and Technology and Economic Assessment Panels. These panels produce an assessment every four years; the latest environmental effects assessment was published in Photochem. Photobiol. Sci., 2003, 2, 1–72. In the intermediate years the panels keep the Parties informed on new developments. The following Progress Report is the 2003 update by the Environmental Effects Assessment Panel.Since the first assessments in 1989, the complexity of the linkages between ozone depletion, UV-B radiation and climate change has become more apparent. This makes it even clearer than before that we are dealing with long-term developments, which can be complicated by large year to year variabilities (Fig. 1 and 2).
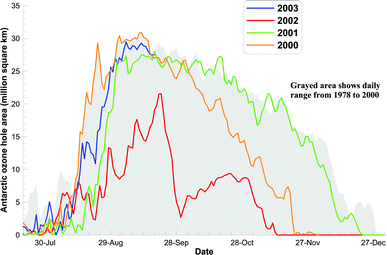 | ||
| Fig. 1 The Antarctic ozone hole area (area with O3 < 220 DU) for 2000, 2001, 2002 and 2003 to date, compared with a 1978–2000 climatology. These calculations are based on the NIWA assimilated total column ozone database.1 | ||
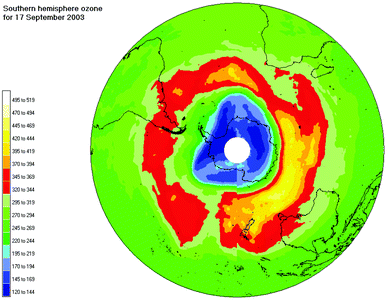 | ||
| Fig. 2 The southern hemisphere total column ozone distribution on 17 September 2003. Data are from the Earth Probe TOMS instrument and graphics are by NIWA. | ||
Ozone and UV changes
The smaller ozone hole in the spring of 2002 should not be interpreted as a sign of early recovery of the ozone layer
The extent of Antarctic ozone depletion varies greatly from year to year depending on meteorological conditions. Satellite data indicate that the ozone hole in 2003 developed rapidly and by early September its area was already close to the previous all-time record, which had been set in year 2000. This is in sharp contrast to the situation in 2002, when the ozone hole was only two thirds of that size. The environmental impacts depend critically on the geographical extent and seasonality of the ozone depletion. Ozone amounts were again low in the Arctic winter of 2002/2003.Evidence for a slowdown in ozone loss in the upper stratosphere since 1997 has been reported
A statistical analysis of satellite-derived ozone profiles indicates that the rate of ozone loss at 35–45 km altitude has diminished globally, and that these changes are consistent with changes in total stratospheric chlorine. This constitutes the first indication of a recovery of ozone at these altitudes.2 However, these reported changes are for ozone in the upper stratosphere only, and detection of any recovery in the total column ozone, which is most relevant for environmental impacts of UV radiation at the Earth's surface, would not be expected for several years.3 The future evolution of ozone remains uncertain, firstly because current chemical models are unable to reproduce accurately all of the observed ozone variability, secondly because the rates of future increases in greenhouse gases are not yet established, and thirdly because interactions between ozone depletion and climate change are not yet fully understood.New estimates of UV radiation extending further back in time are now available at some sites
For example, a method for estimating daily erythemal UV doses using total ozone, sunshine duration, and snow depth has been developed and applied at Sodankyla, Finland, for the period 1950–1999, constituting the longest UV series available to date.4There has been improved co-ordination between groups involved with ground-based measurements, satellite-derived measurements, and radiative transfer modelling, improving the reliability and usefulness of data
For example, the lower concentrations of summertime tropospheric ozone in the southern hemisphere compared with that in the northern hemisphere led to errors in derived ozone columns from some satellite sensors.5 The recent availability of measurements of ozone profiles in the southern hemisphere has enabled improvements in algorithms to retrieve ozone data from satellites. Measurements of UV radiation that are more directly relevant to environmental effects (e.g. photochemical and photobiological effects) are now more widely available.Understanding and quantifying the effects of clouds and aerosols on UV radiation have been improved
Further efforts have been made to assess the contributions from the various factors affecting UV measurements through modelling studies.6 Clouds, and to a lesser extent aerosols, are important modulators of surface UV radiation, which are likely to be affected by climate change. However, an understanding of their future changes, as well as their detailed effect on UV radiation still remains uncertain.There have been further improvements in standardization of UV information provided to the public
This has been achieved through the publication of international guidelines (UV Index) supported by all of the relevant international agencies.7 Several countries are now adopting these standards.Health
A recent analysis of epidemiological studies has confirmed that UV-B exposure contributes to the development of cortical cataract of the eye8
The same analysis concluded that UV-B exposure may contribute to posterior subcapsular cataract. Cataract remains the leading cause of blindness worldwide, and represents an increasing public health burden in both developed and developing countries.High-risk groups for some of the detrimental effects of UV-B radiation, for example immunosuppression, have been identified within the human population by genetic analyses
For skin cancer, important determinants of increased risk include skin type and associated variants of a particular gene (MC1R) related to pigmentation. Risk groups for UV-B induced suppression of immune responses to vaccination have been identified recently on the basis of variation in the gene coding for an immunologically important protein, interleukin-1β9 These people can only be identified by genetic analysis, and the increased risk may not be evident when assessing the population as a whole.It was hypothesized several years ago that exposure to UV-B radiation may prevent or reduce the severity of specific auto-immune diseases and several internal cancers, and new reports add to knowledge in this area
These studies are mainly ecological in nature, correlating geography or climate with disease prevalence. For example, increased latitude has been associated with increased prevalence of multiple sclerosis and insulin-dependent diabetes. Recently, a decreased risk of multiple sclerosis was associated with higher sun exposure at ages between 6 and 15 years.10 The risk of developing prostate cancer was reduced by personal behaviour leading to higher levels of solar exposure.11 Vitamin D synthesis, induced by UV radiation, is proposed as the key mechanism reducing progression of these diseases. Confirmation in an experimental system is required to validate these hypotheses.Additional information confirms that consumption of certain dietary components and supplements may be protective for many of the detrimental health effects associated with UV-B radiation12
Dietary components that have shown such effects include green tea and tomato paste; dietary supplements include lutein and antioxidants such as β-carotene and other carotenoids, vitamin E and vitamin C.Recent work indicates that mild, transient increases in temperature may stimulate immune responses13
The higher temperature associated with climate change may interact with the immunomodulating effects of UV-B radiation.Terrestrial ecosystems
A sizeable proportion of the many recent studies on plant and ecosystem response to UV radiation has been conducted in field settings, both with UV-B supplementation from lamps, and by using filters to manipulate present levels of solar UV
Such research allows more realistic assessments of the role of solar UV. New research on gene regulation in response to UV-B has furthered our understanding of the role played by this radiation, allowing us to make broader generalizations and predictions regarding UV-B effects on plants and ecosystems.Recent work supports earlier observations that under field conditions, higher plants, mosses and lichens are generally well protected from significant UV-B damage, since growth and production are seldom affected14
However, changes in morphology, UV-absorbing pigments, DNA, and antioxidant levels are frequently reported.15,16 Gene activation studies reveal that numerous groups of genes are regulated by UV-B radiation, which may lead to increased DNA repair, synthesis of antioxidants and UV-absorptive pigments.15 Chemical changes induced by UV-B include UV-screening pigments and other compounds that have diverse functions in plants, such as protection against pathogens and insects.16Studies show that the suites of genes activated by UV-B radiation in laboratory conditions are different from those activated in field experiments15
This underscores the importance of conducting studies in the field and may help explain the differences in sensitivity exhibited by plants under experimental conditions where UV-B is supplied by lamps together with low background levels of visible radiation as opposed to plants studied in the field with higher visible radiation.Changes in insect feeding preferences on plants in response to UV-B radiation support earlier observations of increased insect deterrence; however, exceptions have been reported
Gene activation studies indicate a distinct overlap in suites of genes activated by UV-B and those stimulated by insect wounding of plants. The altered expression of these genes may lead to increased insect deterrence.15An increasing number of studies on microbes have been conducted and many show that microbial communities are generally sensitive to UV-B radiation
When exposed to solar UV on the surface of foliage, soil, litter and peat, fungal and bacterial assemblages often change in species diversity and composition.17 Changes in proportions of fungal species on litter have been associated with changes in litter decomposition. Altered decomposition rates have important implications for nutrient cycling in ecosystems. Some microbes not exposed to sunlight, such as those associated with plant roots in the soil, appear to respond indirectly to solar UV.18 Similarly, populations of microarthropods (small invertebrates) living in Antarctic soils decreased with solar UV exposure.19Aquatic ecosystems
Recent research conducted in the field under natural conditions has confirmed results from previous laboratory and mesocosm studies showing that solar UV-B in conjunction with other stress factors can have detrimental effects on primary producers, consumers and decomposers in both freshwater and marine aquatic ecosystems
These studies continue to show that solar UV produces DNA lesions and photooxidative damage in the natural habitat and inhibits biomass productivity and developmental processes. However, they also indicate that many organisms and communities have developed strategies to mitigate detrimental effects of excessive UV. The new field studies indicate that major metabolic and developmental processes require the natural proportion of UV-B, UV-A and visible radiation for optimal performance.20Research has been directed toward understanding the genetic basis and molecular mechanisms of UV-induced responses and response to other stress factors (including elevated temperature, desiccation, increased salinity) in primary producers
Hundreds of genes were found to be involved in the early stress responses and the adaptation mechanisms of cyanobacteria, phytoplankton and macroalgae. These genes control previously unknown metabolic processes that regulate carbon uptake and basic cellular pathways. 21 The role of UV-B-induced reactive oxygen species (ROS) in molecular stress mechanisms has previously been underestimated.22Climate change can alter responses of primary and secondary consumers to UV radiation through temperature-mediated effects23
Future increases in coloured dissolved organic matter (CDOM) related to climate change or other environmental disturbances may subsequently decrease the exposure of zooplankton to UV-B radiation. Multiple factors seem to negatively affect amphibians. These factors include both site specific local effects (such as pesticide deposition, habitat destruction and disease) as well as global effects (such as increased UV-B exposure and global warming).Ecophysiological, paleoecological and genetic studies reaffirm that important components of aquatic ecosystems are sensitive to solar UV24
In addition to influencing biomass producers (phytoplankton and macroalgae), primary and secondary consumers and decomposers (bacteria, viruses and other heterotrophic microorganisms), UV affects alga–bacterium and virus–phytoplankton interactions.25 Although not fully understood, synergistic and antagonistic effects continue to be noted between climate change and the role of UV in aquatic ecosystems.Biogeochemical cycles
Enhanced UV-B radiation can reduce emissions of methane and carbon dioxide from northern peatland microcosms
UV-B results in a reduction in gross photosynthesis, net ecosystem CO2 exchange, and methane emissions from the microcosms. Morphological changes in plant structure can partly explain the reductions in methane emissions.26Satellite imagery has provided a first view of the global distribution of coloured dissolved organic materials (CDOM) that control the penetration of UV-B radiation into the sea and are an important source of sulfur and carbon emissions27
UV-induced photoreactions of CDOM and changes in ocean circulation patterns, such as those related to the 1997–1998 El Niño event, strongly influence CDOM distributions.Models and remotely-sensed data have been used to quantify the time scales of interaction between iron deposition and pigment biomass response
As these linkages are most pronounced at mid-to-high latitudes, these studies highlight the regions where UV flux may interact with the generation of global atmospheric iron and its deposition and the oceanic biological response system.28Sulfur emissions from the ocean are affected by interactions between UV-B radiation and climate change
Dimethylsulfide (DMS) emissions influence the balance between incoming and outgoing radiation in the marine atmosphere. New studies show that loss of DMS induced by UV-B is an important determinant of its concentrations in the surface ocean. These new observations have been made in the North Atlantic Ocean29 and the Southern Ocean.30 The results indicate that the influence of DMS-related atmospheric sulfate particles is modulated by the levels of surface UV radiationUV radiation induces nitrogen oxide (NOx) emissions from terrestrial plants and algae
NOx emissions participate in tropospheric reactions that affect air quality. The UV-induced emissions from plants occur at natural concentrations of NOx and they are possibly linked to plant metabolism or the conversion of nitrate, analogous to that observed on snow surfaces.31 Other research indicates that UV exposure stimulates NOx production from aquatic algae.A newly-identified terrestrial source of ozone-depleting chloromethane is strongly enhanced by global warming
New research has shown that chloride methylation by pectin, which is a widespread constituent in plant tissue, is an important source of atmospheric chloromethane (methyl chloride). The strength of this source increases dramatically when temperatures increase.32Air quality
Stratospheric ozone recovery is calculated to yield more tropospheric ozone and increased atmospheric warming in the future
Calculations predict that tropospheric ozone will increase globally by up to 50% by 2100, ignoring impacts on meteorology. This is estimated to increase by a further 10% due to stratospheric ozone recovery. The impact of this increase on climate is difficult to quantify as ozone concentrations are very variable, both in space and time.33Models suggest that there has been a significant reduction (up to 30%) in the amount of ozone transported from the stratosphere to the troposphere in the last 30 years34
Ozone concentrations in the troposphere have increased due to anthropogenic emissions. Stratospheric ozone depletion has both increased the rate of ozone production in the troposphere due to enhanced UV-B, and reduced the amount of ozone transported from the stratosphere. Current estimates suggest that in total, stratospheric ozone depletion has reduced tropospheric ozone concentrations.The impact of stratospheric ozone depletion on tropospheric OH is probably smaller than previously calculated
Estimates of global OH (the main chemical oxidant in the troposphere) have been made using measurements of atmospheric concentrations of methyl chloroform and an assumed release rate. Recent measurements suggest that methyl chloroform is being released to the atmosphere in quantities much greater than was assumed, and hence that the inferred downward trend for OH in the last decade could be incorrect.35Risks to humans and the environment from substances (such as trifluoroacetic acid and chlorodifluoroacetic acid) produced by atmospheric degradation of HCFCs and HFCs continue to be judged minimal
Although the amounts of these compounds are expected to continue to increase in the future because of climate change and increased use, recent studies have not identified additional environmental hazards.36Materials
The accelerating effect of higher ambient temperature resulting in higher rates of solar UV-induced damage to polymers was confirmed and quantitatively analyzed
A field exposure study on polyethylene and polycarbonate polymers estimated the increased detrimental effects of solar UV wavelengths at higher ambient temperatures. While the results are applicable to specific locations only, the finding could help in assessing the impact of global warming on solar UV degradation of polymers.37Studies on the stability of novel formulations of common polymers exposed to solar UV-B show the role of fillers and additives in enhancing the photodamage process
Some conventional fillers such as calcium carbonate and novel nano-structured inorganic fillers recently available, both contribute to enhanced UV susceptibility of filled polypropylenes.38,39Mixtures of available light stabilizers act synergistically in photostabilization of polyolefins
Recent data also confirm the synergistic effects of conventional light stabilizers used in combination with the hindered amine light stabilizers [HALS] in photostabilizing common polylefin plastics.39,40Recent studies on the solar UV damage to polymer-based coatings provide details of the degradation process and help identify more effective stabilizers
These studies on industrially important classes of coatings, including polyurethane and acylic coatings, provide an improved understanding of their photodegradation process. Several of the studies also confirm the previously reported damaging effects of temperature and humidity on the photodegradation of coatings.41,42References
- G. E. Bodeker, J. C. Scott, K. Kreher and R. L. McKenzie, Global ozone trends in potential vorticity coordinates using TOMS and GOME intercompared against the Dobson network: 1978–1998, J. Geophys. Res., 2001, 106(D19), 23029–23042 CAS.
- M. J. Newchurch, E.-S. Yang, D. M. Cunnold, G. C. Reinsel, J. M. Zawodny and J. M. Russell, III, Evidence for slowdown in stratospheric ozone loss: First stage of ozone recovery, J. Geophys. Res., 2003, 108, 4507, DOI:10.1029/2003JD003471.
- G. C. Reinsel, E. Weatherhead, G. C. Tiao, A. J. Miller, R. M. Nagatani, D. J. Wuebbles and L. E. Flynn, On detection of turnaround and recovery in trend for ozone, J. Geophys. Res., 2002, 107 DOI:10.1029/2001JD000500.
- A. V. Lindfors, A. Arola, J. Kaurola, P. Taalas and T. Svenøe, Long-term erythemal UV doses at Sodankylä estimated using total ozone, sunshine duration, and snow depth, J. Geophys. Res., 2003, 108, 4518, DOI:10.1029/2002JD003325.
- R. McKenzie, D. Smale, G. Bodeker and H. Claude, Ozone profile differences between Europe and New Zealand: Effects on surface UV irradiance and its estimation from satellite sensors, J. Geophys. Res., 2003, 108, 4179, DOI:10.1029/2002JD002770.
- A. Arola, K. Lakkala, A. Bais, J. Kaurola, C. Meleti and P. Taalas, Factors affecting short and long term changes of spectral UV irradiance at two European stations, J. Geophys. Res., 2003, 108 DOI:10.1029/2003JD003447.
- WHO, Global solar UV Index: A practical guide, World Health Organisation (WHO), World Meteorological Organisation (WMO), United Nations Environment Program (UNEP), and International Commission on Non-Ionising Radiation Protection (ICNRP) Report No. Geneva, p. 28.
- C. A. McCarty and H. R. Taylor, A review of the epidemiologic evidence linking ultraviolet radiation and cataracts, Dev. Opthalmol., 2002, 35, 21–31 Search PubMed.
- A. Sleijffers, B. Yucesoy, M. Kashon, J. Garssen, F. R. De Gruijl, G. J. Boland, J. Van Hattum, M. I. Luster and H. Van Loveren, Cytokine polymorphisms play a role in susceptibility to ultraviolet B-induced modulation of immune responses after hepatitis B vaccination, J. Immunol., 2003, 170, 3423–3428 Search PubMed.
- I. A. van der Mei, A. L. Ponsonby, T. Dwyer, L. Blizzard, R. Simmons, B. V. Taylor, H. Butzkueven and T. Kilpatrick, Past exposure to sun, skin phenotype, and risk of multiple sclerosis: case-control study, Br. Med. J., 2003, 327, 316–322 Search PubMed.
- D. Bodiwala, C. J. Luscombe, M. E. French, S. Liu, M. F. Saxby, P. W. Jones, S. Ramachandran, A. A. Fryer and R. C. Strange, Susceptibility to prostate cancer: studies on interactions between UVR exposure and skin type, Carcinogenesis, 2003, 24, 711–717 CrossRef CAS.
- H. Mukhtar, Eat plenty of green leafy vegetables for photoprotection: Emerging evidence, J. Invest. Dermatol., 2003, 121, viii CrossRef CAS.
- J. R. Ostberg, E. Kabingu and E. A. Repasky, Thermal regulation of dendritic cell activation and migration from skin explants, Int. J. Hypertherm., 2003, 19, 520–533 Search PubMed.
- C. F. Musil, S. M. B. Chimphango and F. D. Dakora, Effects of elevated ultraviolet-B radiation on native and cultivated plants of southern Africa, Ann. Bot., 2002, 90, 127–137 CrossRef CAS.
- C. L. Ballaré, Stress under the sun: Spotlight on ultraviolet-B responses, Plant Physiol., 2003, 132, 1725–1727 CrossRef CAS.
- R. Tegelberg, T. Vesala, P. J. Aphalo and R. Julkunen-Tiitto, Clonal differences in growth and phenolics of willows exposed to elevated ultraviolet-B radiation, Basic Appl. Biol., 2003, 4, 219–228 Search PubMed.
- H. Kadivar and A. E. Stapleton, Ultraviolet radiation alters maize phyllosphere bacterial diversity, Microb. Ecol., 2003, 45, 353–361 Search PubMed.
- L. M. Avery, R. I. Lewis Smith and H. M. West, Response of rhizosphere microbial communities associated with Antarctic hairgrass (Deschampsia antarctica) to UV radiation, Polar Biol., 2003, 26, 525–529 Search PubMed.
- P. Convey, P. J. A. Pugh, C. Jackson, A. W. Murray, C. T. Ruhland, F. S. Xiong and T. A. Day, Response of Antarctic terrestrial microarthropods to long-term climate manipulations, Ecology, 2002, 83, 3130–3140 Search PubMed.
- T. B. Norris, T. R. McDermott and R. W. Castenholz, The long-term effects of UV exclusion on the microbial composition and photosynthetic competence of bacteria in hot spring microbial mats, FEMS Microb. Ecol., 2002, 39, 193–209 Search PubMed.
- L. Huang, M. P. McCluskey, H. Ni and R. A. LaRossa, Global gene expression profiles of the cyanobacterium Synechocystis sp. strain PCC 6803 in response to irradiation with UV-B and white light, J. Bacteriol., 2002, 184, 6845–6858 CrossRef CAS.
- Y.-Y. He, M. Klisch and P.-D. Häder, Adaptation of cyanobacteria to UV-B stress correlated with oxidative stress and oxidative damage, Photochem. Photobiol., 2002, 76, 188–196 CAS.
- C. E. Williamson, G. Grad, H. J. De Lange and S. Gilroy, Temperature-dependent ultraviolet responses in zooplankton: Implications of climate change, Limnol. Oceanogr., 2002, 47, 1844–1848 Search PubMed.
- K. Bischof, D. Hanelt, J. Aguilera, U. Karsten, B. Vögele, T. Sawall and C. Wiencke, Seasonal variation in ecophysiological pattern in macroalgae from an Arctic fjord. I. Sensitivity of photosynthesis to ultraviolet radiation, Mar. Biol., 2002, 140, 1097–1106 Search PubMed.
- P. Carrillo, J. M. Medina-Sanchez and M. Villar-Argaiz, The interaction of phytoplankton and bacteria in a high mountain lake: Importance of the spectral composition of solar radiation, Limnol. Oceanogr., 2002, 47, 1294–1306 Search PubMed.
- R. Niemi, P. J. Martikainen, J. Silvola, A. Wulff, S. Turtola and T. Holopainen, Elevated UV-B radiation alters fluxes of methane and carbon dioxide in peatland microcosms, Global Change Biol., 2002, 8, 361–371 Search PubMed.
- D. A. Siegel, S. Maritorena, N. B. Nelson, D. A. Hansell and M. Lorenzi-Kayser, Global distribution and dynamics of colored dissolved and detrital organic materials, J. Geophys. Res., 2002, 107, 3228, DOI:10.1029/2001JC000965.
- D. J. Erickson, III, J. L. Hernandez, P. Ginoux, W. W. Gregg, C. McClain and J. Christian, Atmospheric iron delivery and surface ocean biological activity in the Southern Ocean and Patagonian region, Geophys. Res. Lett., 2003, 30, 1609, DOI:10.1029/2003GL017241.
- D. J. Kieber, R. P. Kiene, D. A. Siegel and N. B. Nelson, Photolysis and the dimethylsulfide (DMS) summer paradox in the Sargasso Sea, Limnol. Oceanogr., 2003, 48, 1088–1100 Search PubMed.
- D. R. Kniveton, M. C. Todd and J. Sciare, Variability of atmospheric dimethylysulfide over the Southern Ocean due to changes in ultraviolet radiatio, in 7th International Conference on Southern Hemisphere Meteorology and Oceanography, Wellington, New Zealand, 2003, pp. 218–219 Search PubMed.
- P. Hari, M. Raivonen, T. Vesala, J. W. Munger, K. Pilegaard and M. Kulmala, Ultraviolet light and leaf emission of NOx, Nature, 2003, 422 Search PubMed.
- J. T. Hamilton, G. W. C. McRoberts, F. Keppler, R. M. Kalin and D. B. Harper, Chloride methylation by plant pectin: An efficient environmentally significant process, Science, 2003, 301, 206–208 CrossRef CAS.
- M. Gauss, G. Myhre, G. Pitari, M. Prather, I. S. A. Isaksen, T. K. Berntsen, G. P. Brasseur, F. J. Dentener, R. G. Derwent, D. A. Hauglustaine, L. W. Horowitz, D. J. Jacob, M. Johnson, K. S. Law, L. J. Mickley, J.-F. Müller, P.-H. Plantevin, J. A. Pyle, H. L. Rogers, D. S. Stevenson, J. K. Sundet, M. van Weele and O. Wild, Radiative forcing in the 21st century due to ozone changes in the troposphere and the lower stratosphere, J. Geophys. Res., 2003, 108, 4292.
- A. C. Fusco and J. A. Logan, Analysis of 1970–1995 trends in tropospheric ozone at Northern Hemisphere midlatitudes with the GEOS-CHEM model–art. no. 4449, J. Geophys. Res. Atmos., 2003, 108, 4449–4449 Search PubMed.
- M. C. Krol, J. Lelieveld, D. E. Oram, G. A. Sturrock, S. A. Penkett, C. A. M. Brenninkmeijer, V. Gros, J. Williams and H. A. Scheeren, Continuing emissions of methyl chloroform from Europe, Nature, 2003, 421, 131–135 CrossRef CAS.
- M. L. Hanson and K. R. Solomon, New technique for estimating thresholds of toxicity in ecological risk assessment, Environ. Sci. Technol., 2002, 36, 3257–3264 CrossRef CAS.
- T. Kuriyama and I. Narisawa, New techniques for evaluating degradation by weathering, in 5th International Symposium on Weatherability, Kogakuin University, Japan, 2003, pp. 97–100 Search PubMed.
- M. Liu and A. R. Horrocks, Effect of carbon black on UV stability of LDPE films under artificial weathering conditions, Polym. Degrad. Stab., 2002, 75, 485–499 CrossRef CAS.
- H. L. Qin, C. G. Zhao, S. M. Zhang, G. M. Chen and M. S. Yang, Photo-oxidative degradation of polyethylene/montmorillionite-nanocomposite, Polym. Degrad. Stab., 2003, 81, 497–500 CrossRef CAS.
- I. Hussain and H. Redhwi, Development of polypropylene-based ultraviolet-stabilized formulations for harsh environments, J. Mater. Eng. Perform., 2002, 11, 317–321 Search PubMed.
- A. D. Skaja and S. G. Croll, Quantitative UV spectroscopy in the weathering of a model polyester–urethane coating, Polym. Degrad. Stab., 2003, 79, 123–131 CrossRef CAS.
- C. Decker and K. Zahouily, Photostabilization of polymeric materials by photoset acrylate coatings, Radiat. Phys. Chem., 2002, 63, 3–8 CrossRef CAS.
Footnote |
| † List of contributing authors in alphabetical order: Anthony Andrady, Pieter J. Aucamp, Alkiviadis F. Bais, Carlos L. Ballaré, Lars Olof Björn, Janet F. Bornman, Martyn Caldwell, Anthony P. Cullen, David J. Erickson, Frank R. de Gruijl, Donat-P. Häder, Mohammad Ilyas, G. Kulandaivelu, H.D. Kumar, Janice Longstreth, Richard L. McKenzie, Mary Norval, Halim Hamid Redhwi, Raymond C. Smith, Keith R. Solomon, Yukio Takizawa, Xiaoyan Tang, Alan H. Teramura, Ayako Torikai, Jan C. van der Leun, Stephen Wilson, Robert C. Worrest and Richard G. Zepp. |
| This journal is © The Royal Society of Chemistry and Owner Societies 2004 |