
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceThe role of Caenorhabditis elegans in the discovery of natural products for healthy aging†
Benjamin
Kirchweger
 *,
Julia
Zwirchmayr
,
Ulrike
Grienke
and
Judith M.
Rollinger
*,
Julia
Zwirchmayr
,
Ulrike
Grienke
and
Judith M.
Rollinger
Division of Pharmacognosy, Department of Pharmaceutical Sciences, University of Vienna, Josef-Holaubek-Platz 2, 1090 Vienna, Austria. E-mail: benjamin.kirchweger@univie.ac.at
First published on 16th August 2023
Abstract
Covering: 2012 to 2023
The human population is aging. Thus, the greatest risk factor for numerous diseases, such as diabetes, cancer and neurodegenerative disorders, is increasing worldwide. Age-related diseases do not typically occur in isolation, but as a result of multi-factorial causes, which in turn require holistic approaches to identify and decipher the mode of action of potential remedies. With the advent of C. elegans as the primary model organism for aging, researchers now have a powerful in vivo tool for identifying and studying agents that effect lifespan and health span. Natural products have been focal research subjects in this respect. This review article covers key developments of the last decade (2012–2023) that have led to the discovery of natural products with healthy aging properties in C. elegans. We (i) discuss the state of knowledge on the effects of natural products on worm aging including methods, assays and involved pathways; (ii) analyze the literature on natural compounds in terms of their molecular properties and the translatability of effects on mammals; (iii) examine the literature on multi-component mixtures with special attention to the studied organisms, extraction methods and efforts regarding the characterization of their chemical composition and their bioactive components. (iv) We further propose to combine small in vivo model organisms such as C. elegans and sophisticated analytical approaches (“wormomics”) to guide the way to dissect complex natural products with anti-aging properties.
 Benjamin Kirchweger | Benjamin Kirchweger is a postdoctoral researcher at the Division of Pharmacognosy, Department of Pharmaceutical Sciences at the University of Vienna, Austria. From 2010 to 2017 he studied pharmacy at the University of Vienna, where he earned his PhD in 2022. His research focuses on the application of Caenorhabditis elegans as well as the use of cheminformatic methods for the discovery of bioactive natural products in the field of aging, metabolism and viral diseases. |
 Julia Zwirchmayr | Julia Zwirchmayr studied Pharmacy at the University of Vienna, Austria, where she received her diploma in 2017 and her PhD degree in 2022. She conducted her doctoral thesis at the Division of Pharmacognosy, Department of Pharmaceutical Sciences under the mentorship of Ulrike Grienke and Judith M. Rollinger. During her stay as a post-doctoral researcher in the lab of professor Rollinger she focussed mainly on the discovery of bioactive secondary metabolites from Nature by implementing NMR-based biochemometric approaches and in vivo studies in Caenorhabditis elegans. |
 Ulrike Grienke | Ulrike Grienke studied Pharmacy and received her PhD in 2011 from the University of Innsbruck, Austria. She has completed several postdocs/research stays in Austria, Ireland, and Brazil. In 2021, she received her habilitation in Pharmacognosy from the University of Vienna, Austria, where she currently works as senior scientist. Her research is focused on the discovery of bioactive constituents from natural sources connecting different methodologies including NMR-based biochemometric approaches and supercritical fluid-based systems. |
 Judith M. Rollinger | Judith M. Rollinger is pharmacist by training. In 2014, she was appointed full professor of Pharmacognosy/Pharmaceutical Biology at the University of Vienna, Austria, where she heads the research group Phytochemistry & Biodiscovery. Since 2020 she is president of the Society for Medicinal Plant and Natural Product Research (GA) dedicated to research on natural products and nature-based drug discovery. In her research she focuses on the interdisciplinary field of integrating big data analytics (cheminformatics, chemometrics) and phenotypic screening (Caenorhabditis elegans) as strategy for the discovery of novel lead structures against infections, metabolic syndrome, and inflammation. |
1 Introduction
1.1. Historical development
Aging is a fundamental and fascinating process that affects all natural organisms.1–3 For humans, aging has always been a subject of curiosity and cultural reflection and it is addressed in religious, philosophical, and pop-culture contexts. It is a complex multi-factorial phenomenon manifesting as a decline of tissue and cell functions and an increase in the risks of many so-called age-related diseases, which include e.g., Alzheimer's disease, Parkinson's disease, osteoporosis, osteoarthritis, type 2 diabetes, cardiovascular disease, chronic obstructive pulmonary disease, and glaucoma. Over the last decades, our understanding of aging has changed dramatically, with a paradigm shift from viewing it as a passive, entropic process to an actively regulated process influenced by a combination of hereditary, environmental, and lifestyle factors,4 which occur despite complex pathways of maintenance and repair.5Hereby, invertebrate model organisms such as the rhabditid nematode Caenorhabditis elegans (C. elegans) have paved the way to effectively study the phenomenon of aging and its pathways.6,7 In the 1980s and early 1990s, research on C. elegans provided some remarkable results. It was discovered that mutations in highly conserved ancient genes such as the daf-2 or age-1 cause a doubling of life span depending on daf-16.8,9 There is strong evidence that the mammal orthologues of these genes, the insulin/insulin-like growth factor receptor, phosphatidylinositol-3-OH kinase and FOXO3 genes also contribute (negatively and positively) to mammal longevity.10–13 These early studies furthered the understanding of genes involved in the regulation of aging and revealed that the function of these gene products, hence aging, is open to interventions.6,14 Today we have identified several drugs that can increase mammal life span significantly.15–18 Although it is still unclear how these approaches translate to humans,19 there is growing consensus that aging and late life multi-morbidity might be open to pharmaceutical intervention. That would provide the opportunity for holistic rather than individual intervention to combat the inevitable effects of aging like cancer, neurodegeneration, cerebral and cardiovascular diseases, blindness, sarcopenia and wrinkles – a regulatory pathway that gets these jobs done, all at once.
1.2. From lifespan to health span
For most of human history, human lifespan stagnated at around 20 to 40 years. Only recently with improvements in medicine, agriculture, sanitation and general living conditions life expectancy has been boosted to over 70 years.20,21 However, the proportion of life spent in good health has not increased to the same extent, meaning that more and more years are spent in poor health.22,23 Hence, multimorbidity and polypharmacy is the norm in old age.24 Thus, the question arises as to whether potential life-prolonging therapy will only extend life, or will also compress morbidity? Interestingly, slowing aging does not necessarily mean reducing morbidity caused by age-related diseases such as cancer, cardiovascular disease, or Alzheimer's disease. Instead, research shows that mutations which extend lifespan can also come with an increased proportion of total life time spent healthy.25–28 However, there are also mutations where increased lifespan simply translates to an extended period of frailty.29 In 2013, Rattan proposed using the term “healthy aging” instead of “anti-aging” to shift the understanding of the aging process as an evolving phenotype due to the failure of homeodynamics.5 According to the World Health Organization (WHO), the term “healthy aging” is defined “as the process of developing and maintaining the functional ability that enables well-being in older age” rather than just the absence of a disease.30 The emerging field of geroscience is an interdisciplinary research area striving to understand the connection between aging and age-related diseases.23,31–33 The goal of geroscience is not to increase life span, but to understand the biology of aging in order to delay diseases and improve health span. In 2013, a conceptual framework of aging was established whereby nine common hallmarks of aging were defined.34 Hereby primary hallmarks develop progressively with time. They comprise genomic instability, epigenetic alterations, telomere attrition and loss of proteostasis. Secondary hallmarks, which include mitochondrial dysfunction, deregulated nutrient-sensing and cellular senescence, evolve as a result. Together with tertiary hallmarks of altered cell communication and reduced stem cell turnover they produce the aged phenotype. A geroscientific hypothesis is that targeting the hallmarks of aging holistically would be an effective approach to delay the pathogenesis of age-related diseases.35,36 Thus, the early prevention of the onset of age-related diseases is considered as ideal approach for extending the health span and achieving healthy aging.37 However, the framework of nine hallmarks of aging has been challenged in the last ten years.38,39 Despite much progress, many challenges remain in formulating an enhanced paradigm of aging.40Yet, it has been shown that the rate of aging, at least in part, can be delayed in mammalian model organisms by genetic, behavioural and also pharmacological means,15–18,41–45 and when aging is delayed, the rate of age-related diseases and conditions will also slow down.46 Since aging is associated with a progressive deterioration of multiple organs, tissues and physiological functions, the usefulness of multicellular organisms for testing is undeniable in the search for aging-modulators. However, there are considerable and pervasive experimental challenges to studying aging in vertebrates and humans. This is especially true for natural products (NPs) including botanicals, because the challenges of sufficient and well-defined material are added. Short-lived animal models that are both, amenable to experimentation and miniaturization, are essential in aging research and provide meaningful guides to the biology of aging in humans.47 Thus, the most-used experimental model organisms for screening the impact of NPs on their age and health span extending properties are small invertebrates such as Drosophila melanogaster and C. elegans.
1.3. Current status
NPs are a recognized source of bioactive molecules. They contributed significantly to the arsenal of approved drugs.48 Isolated NPs and their derivatives, such as rapamycin, resveratrol, and metformin, are probably among the most studied and preclinically advanced interventions against aging and age-related diseases. Botanicals and multi-component mixtures have also been studied early on49 in the context of dietetics.50,51 The interface of healthy aging, C. elegans and NPs has grown immensely since then. A recent analysis of Saul and coworkers52 analysed C. elegans health span literature: according to their analysis, out of the 42 most studied samples in different aging phenotypes, 27 are natural compounds, 12 are extracts; only three samples are not from natural origin. Looking at the 34 most studied agents in test conditions (number of concentrations, temperatures, exposure timings, food and strains), 17 are natural compounds, 12 are extracts, three are NP derivatives (e.g., acetylsalicylic acid) and only two have no natural origin. This dominance of NPs in health span research is supposed to have manifold reasons, such as (i) their increased likelihood for bioactivity;53,54 (ii) the many advantages of C. elegans, especially for NP research;55 (iii) the contradicted but popular free radical theory of aging has led many scientists to focus on antioxidants, such as polyphenols, which are widely found in nature;56–58 (iv) dietary interventions as a primary target for healthy aging and in case of botanicals only a blurry distinction between dietary and pharmacological intervention; (v) traditional knowledge on herbal remedies for symptoms related to healthy aging. (vi) Additionally, some NPs have been subject of anti-aging research through mammal models before the use of C. elegans became the premier model organism in this field;59 (vii) according to the xenohormesis theory, naturally occurring metabolites, such as plant polyphenols might have an evolutionary role as signals to stimulate protective pathways in organisms consuming them.60 Hence their dominance in the field stems from the high likelihood to have health span promoting effects due to a speculated evolutionary imprint.The last decade has witnessed tremendous increase in research activities dealing with C. elegans used to search for anti-aging NP panaceas. The main aspects have been surveyed in the last ten years providing different perspectives on this research field, e.g.,: in 2013, Argyropoulou and colleagues61 gave a profound overview on natural compounds with anti-aging activities derived from phenotypic studies; Pallauf et al. 2017 (ref. 62) reviewed the lifespan-extending effects of flavonoids in different model organisms; Ding et al. 2017 (ref. 63) summarized treatments with reported activity in aging models; Matsunami, 2018 (ref. 64) reviewed literature on C. elegans as model for frailty. Chattopadhyay and Thirumurugan 2018,65 who reviewed 18 dietary and medicinal plants, and Wang et al. 2021,66 who focused on 23 plants from traditional Chinese medicine, both analyzed the longevity promoting effects of plant genera. Several reviews on the longevity promoting effect of natural compounds were also published.67,68 NPs active against transgenic C. elegans models of Alzheimer's disease were surveyed by Navarro-Hortal et al.69 Shen and coworkers70,71 reviewed the use of C. elegans as a model for researching bioactive compounds in food against aging, obesity and Alzheimer's disease. A review on the oxidative and antioxidative potential of NP in C. elegans was also published recently.72 In 2021, Saul et al.52 presented the “healthy worm database” and analyzed literature on health span promoting treatments, including extracts, in C. elegans. These reviews have summarized the outcome of hundreds of research initiatives dedicated to the identification of anti-aging NPs samples by means of C. elegans.
1.4. Scope of this review
A very important research question in pharmacognosy still remains on how to cope with the phenotypic effects from multicomponent mixtures and to identify which molecules trigger those observed bioactivities. It is also imperative to understand how well a read-out from a whole organism, such as C. elegans wild type or mutant strains, can accelerate the search for efficient (herbal) drugs for the benefit for human health in particular with respect to an ever-increasing number of elderly people. The essentials of health span research in C. elegans, which includes worm biology, methods, and assays, were reviewed. In addition, a representative number of studies (169) on NPs research in C. elegans from 2012 onwards, were analyzed with special emphasis on the phytochemical aspects of extracts. This review presents past and present approaches for the discovery of anti-aging NPs implemented in pharmacognostic workflows as well as novel strategies to dissect complex outcomes.2. Caenorhabditis elegans and healthy aging
C. elegans, a basal metazoan organism, offers several advantages as model organism for life sciences. Its use has contributed significantly to many important discoveries. Three of them were honored with noble prizes:73 (i) the discovery of the genetic regulation of organ development and programmed cell death; (ii) the discovery of RNA interference and gene silencing with double-stranded RNA; and (iii) the development of green fluorescent protein as marker for gene expression. C. elegans is also an invaluable model organism especially in the field of health span research and for NP screens. Their small size allows for miniaturized high-throughput procedures at the scale of conventional cell culture methods,74 which offers considerable advantages in terms of resources and the necessary quantities of test substances.75 Large populations of genetically identical animals of synchronized age can be cultured and maintained easily within short time frames and with relatively simple procedures.76 The worm has a rapid life cycle of two to three weeks under standard laboratory conditions and a completely sequenced and well-annotated genome with many well-established resources and protocols available.77,78 The nematode undergoes progressive, degenerative changes while aging, thus providing physiologically relevant data and insights into the underlying mechanism of human diseases.79 Half of the human protein-coding genes have recognizable orthologues in C. elegans.80 Thus, the nematodes have the potential to bridge the gap between in vivo and in vitro approaches in the context of a whole-animal setting,81,82 and have become a popular animal model in aging research.83 Moreover, an invertebrate model comes with certain advantages with respect to legal, regulatory and ethical issues.84,85This chapter briefly outlines main characteristics of C. elegans biology and anatomy, its use in aging research (Table 1), and pathways and genes known to affect its lifespan and health span (Table 2).
| Assay | Principle | Strains | Readout | Advantages | Limitations | Lit |
|---|---|---|---|---|---|---|
| Lifespan | Age-synchronized worms are incubated together with bacteria and test substances for several weeks. Living and dead worms are counted at certain times | N2, BA17, SS104 | Kaplan–Meier plots | De facto standard in most laboratories most direct method to measure aging | Time consuming and tedious process | 75 and 171–173 |
| Mean lifespan | Low technical requirements | No continuous observation | ||||
| Maximal lifespan. Death time 50% (DT50) % survival at specified time point | Researcher bias | |||||
| Automated lifespan | Scanner-based assays | Continuous observation, unbiased readout | High technical requirements, low throughput | 122 | ||
| Flow-cell based assays | 121 | |||||
| Body movement | Manual counting of body bends, bending angle or crawling speed | N2 | Maximum velocity | Most informative metric of health span | Measures are noisy as moving and quiescent states are episodic, researcher bias | 123 and 174 |
| Division of worms into movement classes | N2 | Number of body bends | 148 and 175 | |||
| % of population that can be assigned to defined classes e.g., spontaneously moving, movement only after stimulation or no movements | ||||||
| Pharyngeal pumping | Counting of strokes per time which declines gradually with age | N2 | Pumping rate | Low technical requirements | Measures are noisy and observed effects can be independent of age | 96 and 137–139 |
| Intestinal autofluorescence | Autofluorescent lipofuscin which accumulates with age is quantified | N2 | Either by microscopy and image processing or with a plate reader | Good marker of health | Not aging-specific | 105 and 176–179 |
| Reproductive period | Decline in reproductive ability is an early aging phenotype | N2 | Time span during reproduction is observed | Reproductive span is shorter than lifespan | Regulated independently to other health parameters, not correlated with lifespan | 180 |
| Counting of progenies manually/automated for several days (usually 3–5 days) | ||||||
| Antioxidant capacity | Indirect measurement of ROS levels with dichlorodihydrofluorescein | N2 | Relative fluorescence | Antioxidative effect in vivo | Many interferences, not specific for aging | 150, 151 and 181 |
| Abiotic stress resistance | Worms are challenged with reactive oxygen species producing agents (H2O2, naphthoquinones, paraquat) or heat (35–37 °C) | N2 | Kaplan–Meier plots, % survival at specified time point | Low technical requirements impaired stress resistance is a hallmark of aging | Mechanistic link of ROS and aging is debated not specific for aging | 182–185 |
| Proteotoxicity | Alpha synuclein expression | NL5901, OW13 | Quantification of YFP reporter marked alpha synuclein | Appropriate companion assays, link between longevity pathways and cellular processes (e.g., autophagy, vesicle trafficking and protein quality control systems) required for rescue of these phenotypes | Alpha synuclein is expressed in muscles and not in nerval system | 186–188 |
| Amyloid beta expression | CL4176, CL2006, CL2659 | Protection from paralysis induced by amyloid beta in body wall muscles | Amyloid beta is expressed in muscles and not in nerval system | |||
| CL2331 | Quantification of GFP reporter marked amyloid beta | |||||
| Transcription factor translocation | Measurement of nuclear translocation GFP-marked daf-16, skn-1 or hsf-1 | TJ356, LD1, MAH97, CF1824 | Classification of phenotypes into nuclear translocated, intermediate and cytosolic forms | Mechanistic insight, low technical requirements | Subjective classification of intermediate forms | 120 and 189 |
| Gene | Influence | Human orthologue | Role | Significance | Exemplary NPs |
|---|---|---|---|---|---|
| skn-1 | ↑ ↑ | NRF2 | Transcription factor | skn-1 mutants show decreased resistance to oxidative stress and shortened lifespan220 | Urolithin A,140 curcumin,214 baicalein,222 withanolide A,223 ginsenosides;132 extracts of Lycium barbarum,224,225Hibiscus sabdariffa,226Crataegus pinnatifida,227Apios americana,228Vaccinium corymbosum,229Anacardium occidentale230 and the essential oil of Juniperus communis231 |
| Cellular regulator of oxidative stress response | skn-1 transgenic strains with constitutive nuclear localization exhibit increased oxidative stress resistance, improved health span parameters and mean lifespan is extended by 5–21% (ref. 221) | ||||
| hsf-1 | ↑ ↑ | HSF1 | Transcription factor | hsf-1 overexpression promotes longevity and delays age-related protein misfolding and proteotoxicity232,233 | Trigonelline,212 caffeine;234 extracts of Cratoxylum formosum,187Coffea arabica,234Lonicera japonica158 and Vicatia thibetica235 |
| Protects against age-related proteotoxicity | |||||
| daf-2 | ↓ ↓ ↓ | Insulin/insulin-like growth factor receptor (IR/IGF-1R) | Transmembrane tyrosine kinase receptor | Loss of function mutations in the insulin/insulin-like growth factor signaling (IIS) pathway extend lifespan two-fold8,236 | Orientin202 and naringin;200 extracts of Rubus idaeus,237Ribes fasciculatum,238Hedyotis diffusa192 and Morus alba239 |
| Various functions in metabolism, growth, and reproduction | |||||
| age-1 | ↓ ↓ ↓ | Phosphoinositide-3-kinase (PI3K) | Kinase downstream to daf-2 | ||
| daf-16 | ↑ ↑ ↑ | FOXO | Transcription factor | daf-16 is required for increased life span and enhanced resistance to stress when the IIS pathway is downregulated240,241 | Indicaxanthin,120 geniposide,127 4-hydroxy-E-globularinin,242 hydroxytyrosol,218,243 epigallocatechin gallate,230,244–249 flavonoids149 and silymarin;211 extracts of Rhodiola sp.,131Eugenia uniflora,250Warburgia salutaris248 and Punica granatum251 |
| Various functions in metabolism, cell proliferation and stress resistance | |||||
| sir-2.1 | ↑ ↑ | SIRT1 | NAD+-dependent protein; senses cellular NAD+/NADH levels to regulate transcription, genome stability and many metabolic processes252 | Required for life span extension by caloric restriction; sirtuin overexpression increases longevity by up to 50%253 | Stilbenes,148,195 pentagalloylglucose;190 extracts of Vaccinum uliginosum254 and Camellia sinensis255,256 |
| let-363 | ↑ | TOR | Part of TORC1 and TORC2 kinase complexes; nutrient and energy sensor, tunes protein synthesis and autophagy | Adulthood knockdown of TORC1 pathway genes extends lifespan ∼25% and enhances health span parameters such as motility, stress resistance and pharyngeal pumping257 | Rapamycin257 and extracts of Vigna radiata133 and Ganoderma lucidum258 |
| aak-1/aak-2 | ↑ | AMPK | Kinase; cellular sensor of energy levels, tunes cellular catabolic and anabolic processes | Couples lifespan to information about environmental stressors, energy levels and daf-2 signaling259 mediates lifespan extension through dietary restriction259–261 | Trigonelline212 and orientin;202 extracts of Vigna radiata133 |
2.1. Biology
C. elegans is a soil dwelling non-parasitic nematode which feeds on bacteria and yeasts found in its natural habitat of compost heaps, rotting fruits and plants.86,87 In these microorganism-rich environments it interacts with a diversity of microorganisms and they not only serve as food but also as commensals. In the lab it is usually maintained on agar plates with axenic uracil auxotroph Escherichia coli as bacterial diet. Under these conditions, 99.9% of C. elegans are self-fertilizing hermaphrodites which can each produce more than 250 eggs.88 The eggs are protected by a resilient shell, which allows them to resist even very adverse environments. After hatching, the life cycle of C. elegans consists of four larval stages L1 to L4. The end of each larval stage is characterized by a molt, until the reproductive adult stage is reached. In the absence of food, worms can arrest in L1 stage and survive for several days, or, if they have already reached the L2 stage, enter the dauer stage at which point they worms can survive for several months.89 For longer periods of time, larvae can even be stored in a cryogenically frozen state at −80 °C.76,90 Thus, large numbers of strains can be easily maintained. To generate an age-synchronized culture for lifespan experiments, worm cultures are bleached with an alkaline hypochlorite solution, whereby worms are sensitive to the bleach and disintegrate while the egg shell protects the embryos from death. The eggs are then agitated until they hatch, while in the absence of food they arrest at a synchronized L1 stage.91 Protocols that make use of filters to isolate eggs have also been described.922.2. Anatomy
C. elegans (Fig. 1) is an unsegmented pseudocoelomate and lacks respiratory and circulatory systems. It is enveloped in a cuticle, a collagenous, extracellular, exoskeleton that shields the animal from its environment and maintains the morphology and integrity of the worm.93 The cuticle is synthesized by a hypodermal cell layer and plays a critical role in body movement because it is attached to muscles.94 The worm's mouth is at the anterior of its body. It is a small opening with a cavity that is separated from the intestinal lumen by the pharynx, a neuromuscular pump and one of the most complex organs of the worm. The pharynx is comprised of eight muscle cells, 20 neurons, and epithelial, support, and gland cells.95,96 The rhythmic contraction of pharyngeal muscles, referred to as pumping, sucks bacteria into the mouth of the worm, mechanically grinds them and transports the food into the intestinal lumen. The pumping rate is dependent on food availability, worm age and can be altered by compounds that modulate e.g., serotonin signaling.97 The intestine consists of 20 cells and has various functions apart from digestion and nutrient absorption including nutrient storage and synthesis of macromolecules and yolk. Yolk particles are lipoproteins, which are packaged into oocytes in fertile worms. Four enteric muscles located at the posterior of the intestine and body work for defecation.98 The gonad of the hermaphrodites is an ovotestis that produces sperm in the L4 stage and oocytes in the adult stage. As a result, the worm produces up to 300 progenies by self-fertilization within a few days, a process that involves self-destructive biomass repurposing.99 Noteworthy features of the worm's anatomy are also six macrophage-like scavenger cells, the coelomocytes,100 and 95 body wall muscle cells.101 For the worms' 302 neurons a complete map of synaptic connections, the connectome, is available. | ||
| Fig. 1 Brightfield image of adult hermaphrodite C. elegans with scale and visible organs. | ||
2.3. C. elegans assays related to healthy aging
The aged phenotype of C. elegans develops dynamically over time until death. Many age-related changes on the molecular and tissue level become apparent at the level of the whole organism and can be easily assayed e.g., via behavioral phenotypes and morphological markers. These changes start from the first day of adulthood. Already at day 2 of adulthood learning and long-term memory worsens.102 Around day 4 of adulthood the hermaphrodite enters the post reproductive period and yolk lipoproteins start to accumulate ectopically in the body cavity.103,104 This is accompanied by gradual intestinal atrophy and an increase of autofluorescent pigments which represent probably advanced glycation end products.105 The two posterior V5-derived (PVD) neurons for sensation of mechanical stimuli and cold temperatures degenerate.106,107 At day 7 of adulthood, sarcopenia is apparent histologically by a progressive disorganization of sarcomers,108 and behaviorally by a reduced motor activity. The latter one is also caused by stiffening and thickening of the cuticle caused by an unregulated collagen biosynthesis.108 Pharyngeal pumping also slows gradually due to an aging-related decline of pharyngeal muscles and other pathologic changes. While young adult worms have a pumping rate of more than 200 pumps per minute, old adults achieve less than 50 pumps per minute.109,110 Since the duration of contractions is extended, the grinder fails to break down the bacterial food. Hence, living bacteria increasingly colonize the intestinal lumen causing it to bloat.111,112 Whether and how this dysbiosis subsides with a more natural bacterial culture and a functional microbiome, however, still needs to be investigated. Around day 10, the body motions become sporadic. Their response to stimuli like plate shaking, light or touching decreases. Overall, old adults shorten and get thicker around the midbody area and their bodies begin to wrinkle, an indicator of physical deterioration.108,113 Death in C. elegans is usually assessed as non-response to harsh stimuli. A wave of necrotic cell death in the intestine and a burst of intense blue “death” fluorescence are reliable indicators of organismal death.112,114 The wormatlas has reviewed the aging of the worm and provides very good figures and video sequences that are available to the scientific community.115 To assess NPs-based biological effects in C. elegans, the following pharmacological endpoints can be determined by means of (i) survival (lifespan and lethality), (ii) behavioral changes (pharyngeal pumping and locomotor activity), (iii) histological parameters (autofluorescence, muscle fiber organization), (iv) reproduction (morphologic deformity, brood size and larval development), (v) resistance to stressors (heat or reactive oxygen species (ROS)), (vi) biochemical markers, e.g., measuring ROS levels, or (vii) gene/protein expression and cellular localization. Noteworthy assessments are also (viii) proteotoxicity assays that model amyloid beta, α-synuclein or polyglutamine toxicity (Table 1).72,116(i) Survival assays. Lifespan assays are probably the most direct method to determine the effect of test samples on aging. Hereby, cohorts of worms from the same synchronized populations are created by separating them into different wells or different petri dishes. Then they are challenged by interventions such as different diets, compound treatments, or double-strand RNA (dsRNA). Living and dead worms are counted at specified intervals (e.g., every day, every second day). Usually, Kaplan–Meier survival plots are generated to illustrate the percentages of live worms in the cohorts over time, and the data are statistically analyzed.117 These assays can be performed either on solid agar or in liquid media. Both come with advantages and limitations reviewed elsewhere.117,118 Manual counting in these conventional lifespan assays is time-consuming and tedious and it comes with several limitations such as researcher-oriented bias and the exposure of worms repeatedly to light, heat or mechanical stresses. That is why more and more approaches to automate lifespan assays have been developed. The lifespan machine automatically captures sequential images of worm populations on agar plates. It determines death events when worms stop crawling for long periods of time with image processing software.119,120 Other methods employ flow cells where fresh media, food and interventions enter, while eggs, metabolites and debris exit the flow cell. Digital video recording allows for continuous observation.121 Beside death events, health span parameters like locomotion, morphology and behaviour can be assayed.122 Of course, this comes at the expense of high technical requirements and a decreased throughput compared to microwell-based approaches.
(ii) Behavioural assays. Maintaining mobility and physical fitness is an important indicator of healthy aging. In C. elegans it is also an important predictor of lifespan.110,123,124 The worm's locomotion gradually declines with age and is therefore a straight-forward health span marker. The rate of body movement can be accessed with markers such as maximum bending amplitude, crawling speed on agar,125–127 number of body bends during fixed intervals of time or bending angle.128 The locomotion of worms can also be measured at the cohort level.129–131 In this case, cohorts of worms are generated and treated with samples, similar to life span studies. Then the nature of the worms' movements on successive days is examined, dividing the worms into motility classes as in the most unbiased way as possible e.g., healthy class worms that exhibit symmetrical and spontaneous movements, aged class worms that exhibit uncoordinated, stiff movements, and very aged class worms that move only the tail or head in response to stimuli. In this way it can be shown how a treatment slows down the age-dependent motility decline.132,133 However, the classification of worms is prone to researchers' bias and body movements are noisy, because behavioural states are episodic. Food and neuromodulators like exogenous 5-HT can substantially inhibit movements without necessarily affecting aging.124,134–136 A further measure of aging in C. elegans is the amplitude of pharyngeal pumping.137 Hereby, pharyngeal contractions are counted during fixed intervals of time. As the pumping rate can have irregular rhythms, e.g., showing pumping bursts,97 it is necessary to count pumping of worms several times and calculate the mean pumping rate.136 Technical requirements are low, a routine stereomicroscope and a hand counter is all that is needed.96 However, the repeated measurements of pumping are cumbersome and prone to researcher bias. There are also methods to automate this measurement with image analysis of consecutive image frames138 or by recording electropharyngeograms with electrodes.139
(iii) Histological parameters. A histological marker of health span is the intensity of intestinal autofluorescence. Red channel fluorescence correlates well with the worm's remaining days of life, and is therefore a good marker of health.105 Although not as frequently employed, muscle fibre organization is also a good surrogate parameter of health in aged worms. It can be assayed with transgene strains showing GFP-marked myosin (e.g., strain RW1596).108,140,141
(iv) Reproduction. Another frequently employed health span assay is the measurement of the reproductive period. In the L4 stage, the worms produce around 300 sperm cells. With the advent of adulthood, they switch to the production of oocytes which are continuously fertilized with sperm. When everything runs smoothly the worm can produce up to 300 progenies. The ability to produce eggs for a longer time period is sometimes regarded as health parameter but it is largely regulated independently of other health parameters and has a poor correspondence with lifespan.110 Nevertheless, it is useful to probe if health span is achieved via suppression of reproduction.142,143
(v) Resistance to stressors. As most health span prolonging pathways also promote resistance to various abiotic factors like ROS generating chemicals, ultraviolet radiation or heat stress, assays that test for such properties are frequently used as surrogates to identify new health-promoting genes and agents.144–147 This stress resistance is often measured by exposing worms to elevated temperatures (e.g., 35 °C) or a ROS generating chemical (e.g., juglone, paraquat or peroxide) and comparing the survival of treated and untreated worms. This is also performed semi-automatically in plate-reader format using the Sytox®Green reagent which can only penetrate necrotic cells where it leads to a strong increase of fluorescence. Thus, SYTOX fluorescence is positively correlated to worm mortality rate and negatively related to their stress resistance.148,149
(vi) Biochemical markers. 2′,7′-dichlorodihydrofluorescein (DCF) is a small-molecule fluorescent probe which is frequently used to assess intracellular ROS in C. elegans. Upon cellular uptake DCF is oxidized by ROS to the fluorescent product 2′,7′-dichlorofluorescein. Thus, DCF fluorescence serves as an indicator of the accumulation of oxidative damage in worms, which is associated with senescence.150 However, this oxidative stress theory of aging is increasingly challenged.151,152 In addition, most hits of these DCF assays are not followed up in detail and the documentation of the experimental protocol is sometimes inadequate. This is critical because the measurement of ROS and oxidative damage in general, and in particular with DCF, can be prone to many interferences and problems as summarized recently.153
(vii) Protein expression and cellular localization. For a more mechanistic insight into health span promotion by treatments, western blot analysis, quantitative PCR or MS-based proteomics are used.83,154,155 The expression of several gene products which are associated with an improved health span e.g., detoxification factors such as heat shock proteins, metallothioneins and superoxiddismutase, can be monitored by transgene worms carrying GFP-reporters.156–159 Noteworthy are also strains carrying GFP fusion proteins with pro-longevity transcription factors such as daf-16 and skn-1 where a translocation to the nucleus can be monitored in response to treatments with fluorescence microcopy.160
2.4. NPs in C. elegans-based drug screening
Treatment with test agents in C. elegans can be achieved by several methods which vary between studies. When worms are tested in a liquid buffer, test samples can be added to the medium in a given concentration similar to cell culture. However, C. elegans cultures on agar plates require mixing into or the spreading of the samples onto the agar. An alternative is to supplement the agents to the worm's food, the bacteria. The concentrations of test substance used and the concentrations achieved in the worm can often differ by several orders of magnitude. The agents can be absorbed by diffusion through the cuticle, the uptake via sensory cilia of neurons or through ingestion of bacteria which have taken up the samples. The latter approach is the most relevant for most drug-like molecules.161 This comes with some caveats as the living uracil-auxotroph E. coli has a considerable potential to metabolize test agents.162,163 The alternative of feeding inactivated bacteria, proposed by some researchers,163 significantly impairs worm development.164,165 However, C. elegans also has a remarkable xenobiotic defense system with many xenobiotic detoxification enzymes and efflux pumps.166,167 It should therefore come as no surprise that drug concentrations several magnitudes higher than can be achieved in human plasma are used. Burns and colleagues reported that only 2% of bioactive compounds can produce a robust phenotype at a concentration of 25 μM.168 However, certain NP classes such as flavonoids show good bioavailability.149 Therefore, it is necessary to consider appropriate carrier concentrations, solubility and properties of test extracts and compounds, to prevent precipitation. We and other groups have observed that DMSO concentrations up to 1% do not lead to significant changes of lifespan,75,169 although it is clear that high DMSO concentrations have impact regulatory pathways, physiologic rates and drug bioavailability.170Drug delivery, dosage, and time of administration also have major effects and may also be a major cause for contradictory results in the literature. Just to name two examples: for instance, when resveratrol is added to worms at L1 stage, it extends lifespan.190–192 However, if resveratrol is added later in life at L4 or adult stage, it is reported to have highly variable outcomes with either a significant lifespan extension or not.148,193–196 Another example is naphtazarin treatment, which in concentrations of up to 500 μM causes a lifespan extending effect on solid media,197 while in liquid culture it exerts a potent nematotoxic effect already at 79 μM.198 This controversial effect may be due to a greater uptake of compounds in liquid culture as it is reported for resveratrol and fluoxuridine.161 “Sola dosis facit venenum” – the dose makes the poison. This maxim attributed to Paracelsus applies very well to C. elegans lifespan modulation. Low doses of toxins can sometimes have beneficial effects on lifespan through not fully understood mechanisms which may include the upregulation of stress response pathways. This was shown for naphtoquinones197 and arsenite,199 nor can the increase in lifespan be expected to be linear or exponential with the dose of supposedly non-toxic compounds. For most compounds inverted U-shaped dose response curves with highest effects at medium doses were shown for e.g., naringin,200 withanolide A,201 orientin202 and quercetin.203 This non-linear dose–response poses a problem both for classical bioassay-guided fractionation but also for statistical correlations where active ingredients are identified based on the correlation of bioactivity and compound concentration.
Attentiveness is also warranted in the interpretation of results: C. elegans health span is highly sensitive to environmental variables like temperature, light, solid or liquid culture and composition of media. While these variables can be minimized with specialized equipment and handling, other parameters like standardization of the bacterial food source are more complex. Therefore, two-tiered cell banking systems, OD measurements for optimal harvesting points as well as precise harvesting and storage protocols need to be implemented to reduce variations. However, variation in adult survival of synchronized worms even in the same well can still be substantially different. Age-related pathologies appear to occur stochastically, and so do the resulting deaths.108 According to the disposable soma theory the post-reproductive survival is not under evolutionary pressure. It is therefore more likely to vary. A high number of technical replicates and several parallel and independent experiments are necessary to confirm and quantitate lifespan increase.
2.5. Pharmacological dissection
A major advantage of C. elegans is that after identification of health span promoting compounds, the deconvolution of required pathways and targets is feasible. Several pathways have been discovered that mediate the life-prolonging effects of NPs. The best studied is probably the insulin and insulin-like growth factor-1 (IIS) pathway with the tyrosine kinase receptor daf-2 and the downstream phosphoinositide- 3-kinase age-1. When activated, this pathway inhibits the transcription factor daf-16 and thus the expression of downstream genes involved in longevity. Knockdown of daf-2 and age-1 genes can prolong lifespan up to two times. There are other pathways frequently named in mediating lifespan prolonging activities of NPs. The putatively best studied ones are summarized in Table 2. NPs with a well demonstrated influence on such pathways have also shown to impact healthy aging in other model organisms and mammals e.g., the urolithins.140The worms' simplicity and amenability to sophisticated, yet convenient, genetic techniques is a big advantage.204 There are long established genetic tools and resources available.78,79,205–208 The screening of nonsense and missense mutant strains, which are easily available (e.g., from the Caenorhabditis Genetics Center)207 can foster the deconvolution of gene products that are required for life span extension.209 In this way, a plethora of NPs, including emodin, trigonelline, naringin, silymarin flavanolignans and hibiscus extract, have been shown to extend health span in a daf-16 -dependent manner.200,202,210–212 Other NPs like acetyl-phenethylamine and ginsenosides require (also) the function of sir-2.1 or skn-1,132,213,214 whereas orientin and urolithin A require aak-2.140,202 Another possibility is to employ RNA interference which can be achieved by feeding bacteria expressing gene-specific dsRNA. It allows for knockdown in adults to avoid development defects (e.g., in daf-2 mutant). With specific promoters, tissue-specific RNA silencing is possible.215,216 The worms' transparent body allows for researchers to visualize and quantify molecular and cellular processes like the expression, localization and activity of proteins.217 A frequently employed approach is to use transgene worms carrying fluorescent protein tagged transcription factors to test for nuclear translocation in response to treatment.218 In this case daf-16::gfp (e.g., strain TJ356) and skn-1::gfp (e.g. strain LD001) are useful to prove an involvement of these pathways. Also transgene worms expressing hsp-16.2::gfp, sod-3::gfp and gst-7::gfp are employed to test the induction of these antioxidant enzymes and stress reporters.219 Gene expression levels can also be determined via western blot but larger populations (up to 1000 worms) or on RNA level via quantitative real-time polymerase chain reaction.175 A shortcoming of these techniques is that there is no smoking gun in the sense of a discovered specific drug – target binding event. Since that event is causative for the modulation of protein function and the disorganization of a pathway, it results in lifespan extension. This knowledge could foster linking chemo-structural details to health span promoting phenomena at the organism level, which is valuable to structurally optimize compounds towards improved efficacy.
3. Natural compounds with anti-aging effects in C. elegans
A variety of natural compounds from almost all structural classes have been investigated in the past ten years for their effects on aging in C. elegans. They are presented in ESI Tables ESI1–14† and are grouped into structural classes without making a claim to completeness. Additionally, Tables ESI1–14† provide information on the experiments, their parameters (e.g., dose, strain) in C. elegans, the findings of the respective studies and the report of signalling pathways or genes involved in the observed mechanism. We have gathered data from 160 compounds reported in 85 representative studies that show healthy aging effects in C. elegans. The most researched compounds were epigallocatechin gallate (1, thirty experiments), resveratrol (3, seventeen experiments), withanolide A (2, fourteen experiments), hydroxytyrosol (4, fourteen experiments), cannabidiol (5, eleven experiments), quercetin (6, ten experiments), tyrosol (7, ten experiments) and 4-hydroxybenzoic acid (8, ten experiments).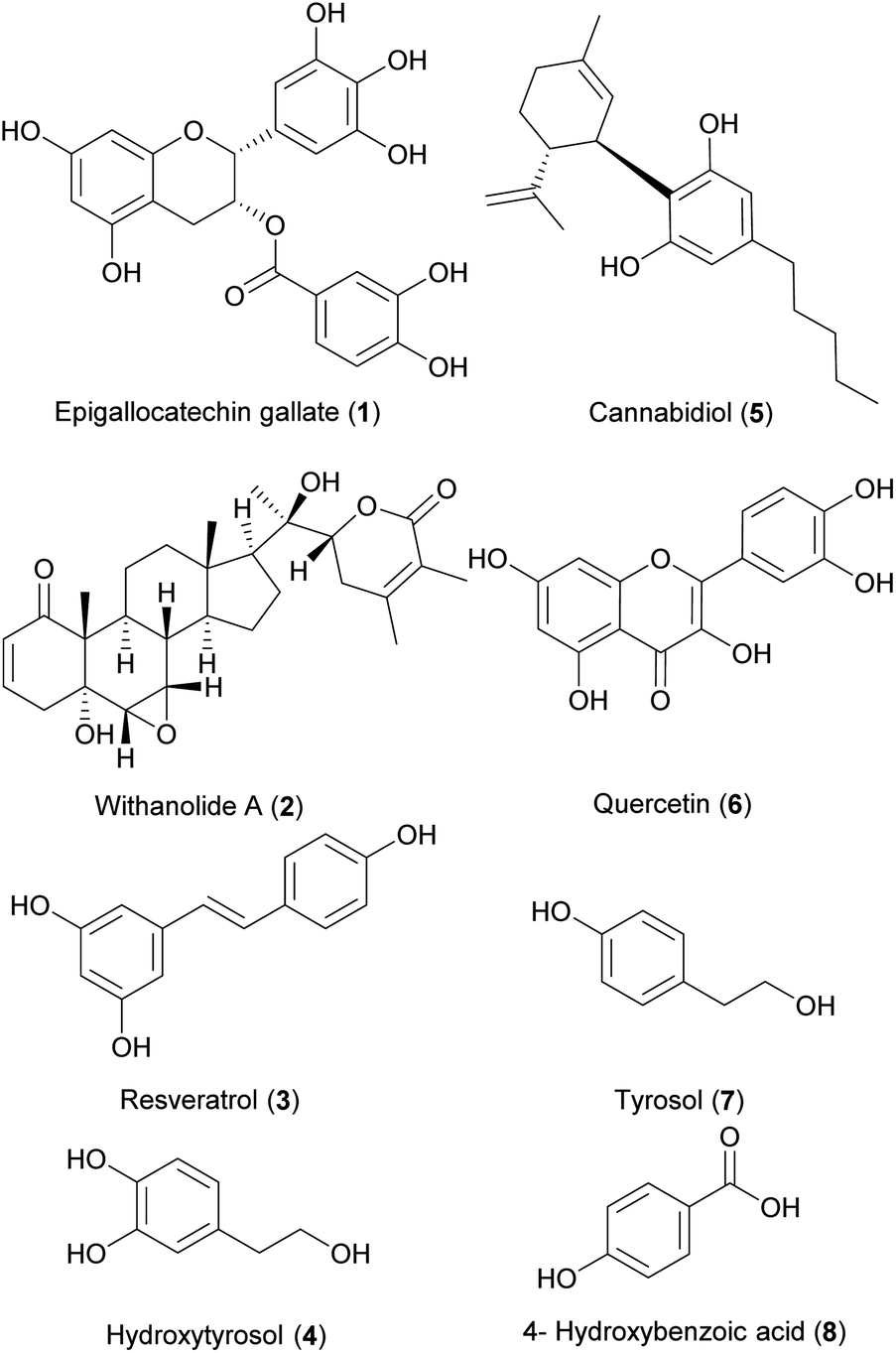
A continued focus on structural classes with phenolic structures were observed (Fig. 2) with 92 out of 166 compounds being phenols. Most of these compounds can be ascribed to the compound classes of flavonoids (fourty compounds), hydroxycinnamates (eleven compounds), tannins (thirteen compounds), and stilbenes (seven compounds). This focus can be attributed to the fact that dietary polyphenols, e.g., from olives, tea, fruits, herbal teas and vegetables have long been suspected of preventing aging associated diseases.
 | ||
| Fig. 2 Structural classes of NPs investigated in C. elegans for their health span promoting effects. | ||
It is also striking that almost one in four compounds were glycosides (thirty-nine compounds) with saponins, iridoids and flavonoids as frequent scaffolds. This is interesting because glycosylated NPs are usually not in the focus of interest for drug discovery projects,219 due to their poor bioavailability in humans caused mainly by hydrolysis by the intestinal flora.262 Whether there is a similar metabolism in the C. elegans – OP50 model remains elusive. Only two studies had looked into the fate of glycosylated NPs during incubation with bacteria.263,264 It would be interesting to determine which metabolites actually arrive in the worm and cause the observed effects.
However, there are also reports on healthy aging promoting terpenes, alkaloids, betalains, diarylheptanoids, anthraquinones and non-proteinogenic amino acids (Fig. 2). A considerable part of the collection was subsumed under the class “fragment-like NPs”.
3.1. Examples for investigated natural compounds
Urolithin A (9) is one of the major gut bacterial transformation products of ellagitannins e.g., of pomegranate fruits and berries. In humans the endogenous formation of 9 varies dramatically depending on the individual microbiome composition.265 Therefore, it was suggested that 9 could also be supplemented directly to the diet. An application for 9 as a novel food ingredient in the European Union was filed in 2018.266 Supplemented 9 has shown to promote healthy aging across several species including C. elegans, D. melanogaster, and Mus musculus. The proposed mechanism of supplemented 9 is the induction of mitophagy, which comes with a contradictory improved mitochondrial function and proteostasis in later life, probably due to the elimination of dysfunctional mitochondria. Long-term treatment of C. elegans with urolithin A influences the regulation of mitophagy and mitochondrial biogenesis through the transcription factor skn-1. It increases mean and median lifespan but also improves other health span parameters like pharyngeal pumping, respiratory capacity, mitochondrial content, muscle fibre organization and locomotion in aged worms. The urolithin A mediated induction of mitophagy is conserved over species and also in aged rodents enhances muscle strength and running endurance.140 A phase 1 clinical study confirms that urolithin A is bioavailable in humans. A long-term oral consumption (4 weeks period) has been demonstrated to be safe and shows signs of mitochondrial and cellular health improvements also in aged humans.267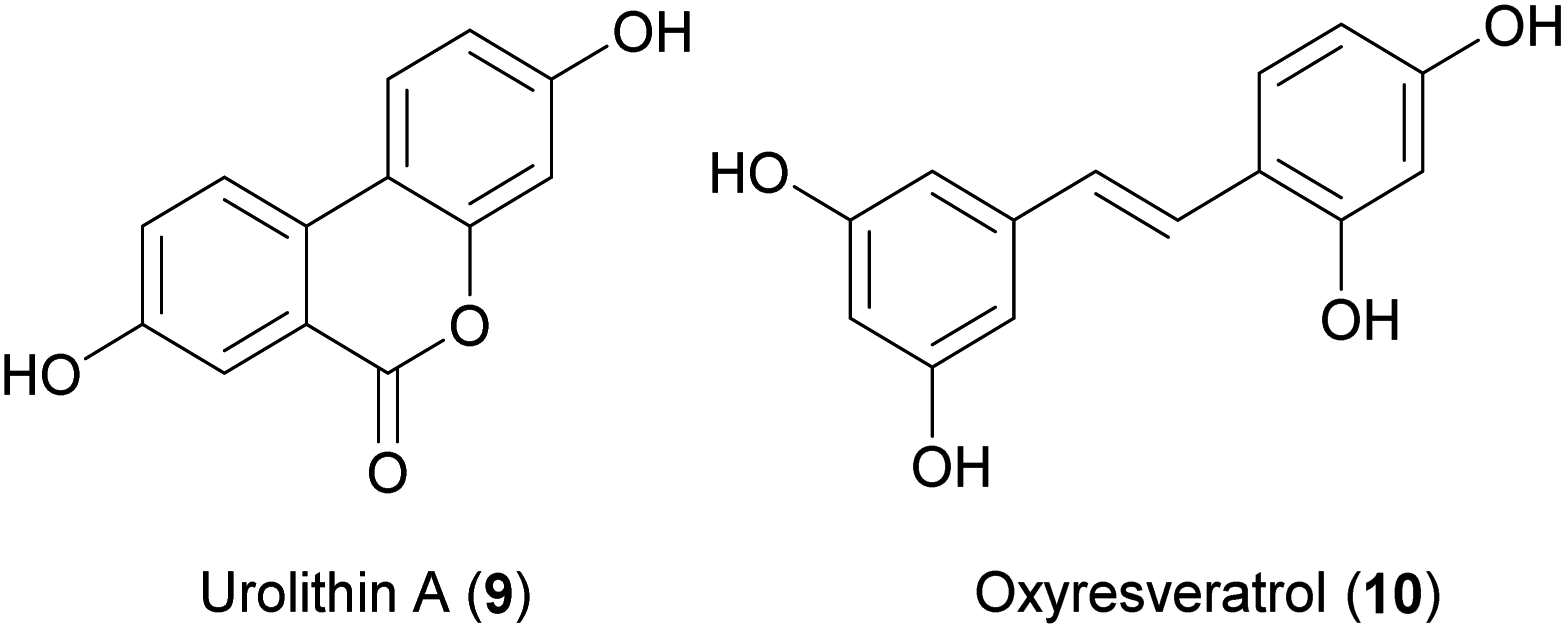
The structure class of stilbenes, with its well-known representatives' resveratrol (trans-3,4′,5-trihydroxystilbene, 3), oxyresveratrol (10) and pterostilbene, has been repeatedly investigated in the context of aging across different species. A study on the life-extending effect of resveratrol in small metazoan organisms including C. elegans was published as early as 2004.268 Although resveratrol has been used as a positive control for lifespan experiments in many studies over the last 10 years,190,191,269 showing mainly positive effects, there are two studies that could not reproduce this lifespan-extending effect.148,193 Also, natural and synthetic derivatives of 3 were investigated: it has been shown that 10 has a stronger life-prolonging effect than 3,195,196,270 whereas pterostilbene has no effect.148 Several synthesized stilbene derivatives have been reported to increase the lifespan of C. elegans more robustly than 3.148
After nearly 20 years of anti-aging research with 3, it is still highly controversial whether it can extend the lifespan of C. elegans or other model organisms.148,195,271 Two studies performed with rodents have found no statistically significant life extension upon treatment with 3.45,272 One study reported that 3 improved life- and health span of mice on a high-calorie diet;273 another noted a positive influence on healthy aging on mice fed a standard diet.274 Sirtuins (sir-2.1 in C. elegans) have been proposed to account for the health span promoting effects of 3 similar to a caloric restriction mimetic.275 Mammal SIRT1 activation of resveratrol was shown in vitro and in vivo.276,277 As caloric restriction can vary widely by genotype or diet, it is a conceivable cause of conflicting in vivo results.278 However, the interaction of 3 with sirtuins is also not without controversy as biochemical assays were called into question and results in model organisms are often not reproducible271,279–2813 is contained in282 larger amounts in berries of the genus Vaccinium (e.g., cranberry, billberry, cowberry)283 and redcurrant, as well as in smaller amounts in many other plants such as peanuts, pistacchios, apples, tomatoes, grapes and cocoa.284 However, the concentrations of stilbenes in food and that of in this context often cited red wine284,285 are low compared to doses of preclinical and clinical studies. Considering also the poor bioavailability of 3,286 claimed health benefits for different foods are questionable.
Flavonoids are consumed in substantial quantities by humans. Dietary intake is associated with positive effects in epidemiological studies against age-related diseases including diabetes mellitus type II,287 cardiovascular disease288,289 and cognition in elderly.290 Therefore, they were among the first research subjects tested on C. elegans,49,146,291 and they remain a prominent NP-class in the literature over recent years (Fig. 2). Baicalein (11), a flavonoid isolated from Scutellaria baicalensis, is known to be an activator of the transcription factor Nrf2 in mammalian cell lines.222 In C. elegans it modulated stress-resistance against lethal thermal and sodium arsenite stress and dose-dependently extended the nematodes' lifespan via the Nrf2 orthologue skn-1. The FOXO orthologue daf-16 was not involved in these processes.292 The polymethoxylated flavonoid tangeretin (12), a constituent of citrus fruits, extended the mean lifespan C. elegans at 30 and 100 μM. It slowed aging related functional declines and increased the resistance against heat-shock stress. On the mRNA expression level, daf-16, hsp-16.2, and hsp-16.49 were upregulated. Tangeretin promoted the nuclear localization of daf-16 and enhanced the expression of hsp-16.2. No effects were observed on the lifespan of daf-2, age-1, and daf-16 mutants. Hence, it was suggested that besides an extension of lifespan, tangeretin enhances heat stress tolerance in an insulin/insulin-like growth factor signaling dependent manner.175 Naringin (13), a flavanone-7-O-glycoside consisting of the flavanone moiety naringenin and the disaccharide neohesperidose, is another well-known constituent of citrus fruits, in particular of grapefruit. This compound was found to extend the lifespan of C. elegans. Moreover, it increased the thermal and oxidative stress tolerance and reduced the accumulation of lipofuscin. In Alzheimer's and Parkinson's disease models, it delayed their progression via daf-16.200 Orientin (14), the 8-C glucoside of the flavonoid luteolin, occurs in a wide range of medicinal plants, for instance in plants of the genus Nasturtium. In C. elegans, it increased the lifespan, improved heat, oxidative, and pathogenic stress resistance through the activation of stress responses, including hsf-1-mediated heat shock response, skn-1-mediated xenobiotic and oxidation response, mitochondria unfolded responses, endoplasmic unfolded protein response, and increased autophagy activity. Moreover, it activated AMPK and daf-16. In neurodegenerative disease models of C. elegans, it reduced the accumulation of toxic proteins (α-synuclein, β-amyloid, and poly-Q) and delayed the onset of the respective disease.202
Several studies have shown good bioavailability of flavonoid aglycones along with their extensive metabolization by conjugation in C. elegans293–295 Treatment with quercetin-3-O-glucoside led to higher accumulation of 6 compared to treatment with the aglycone, which shows that the flavonoid glycoside was taken up and deglycosylated by the worm.264
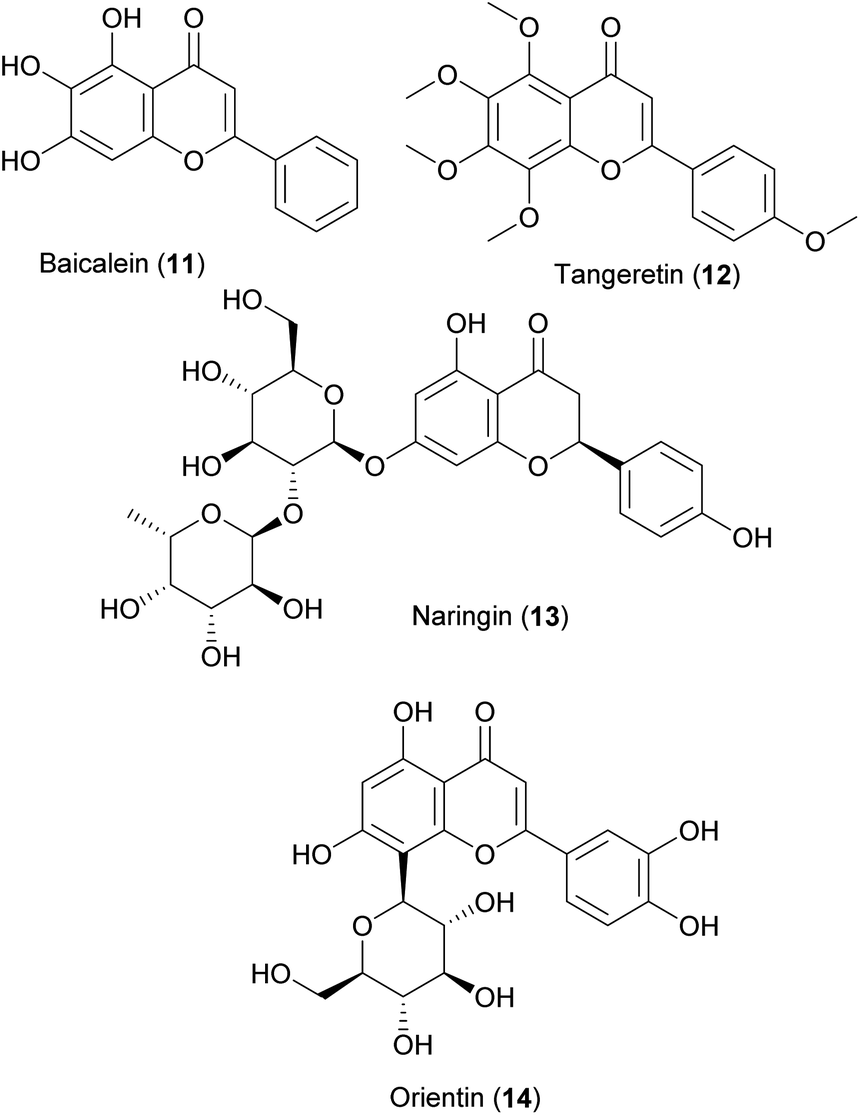
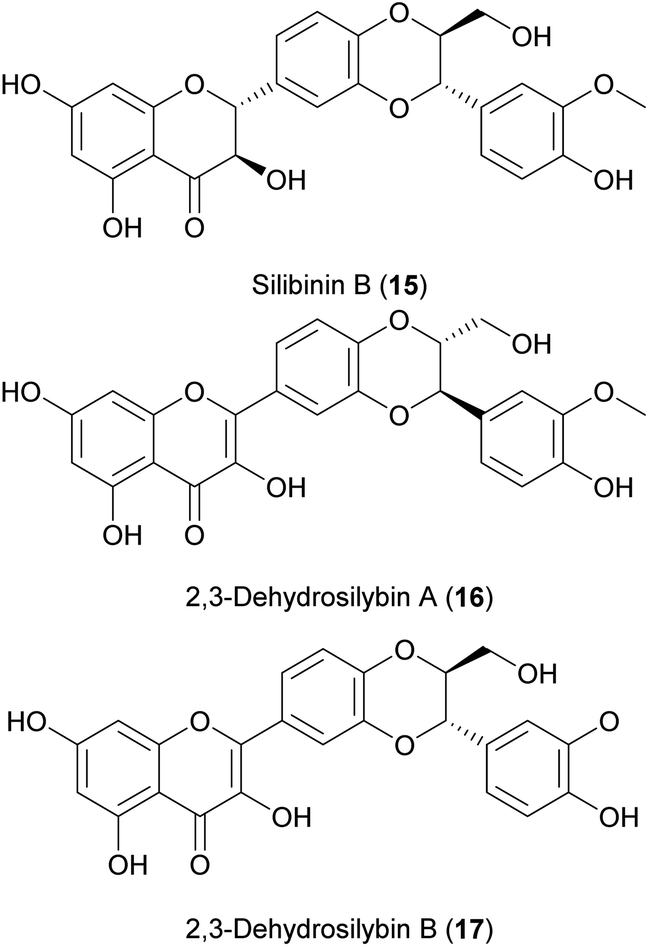
In 2017, Sciacca and co-workers, investigated four pure compounds of the NP flavonoid complex silymarin from milk thistle (Silybum marianum (L.) Gaertn.), i.e. silybin A, silybin B (15), 2,3-dehydrosilybin A (16) and 2,3-dehydrosilybin B (17), regarding their effects on the inhibition of Aβ amyloid growth and toxicity in C. elegans. Underlining the crucial role of stereochemistry, from all four constituents tested, silybin B was found to be the most effective in counteracting Aβ proteotoxicity.296 In the same year, Filippopoulou and co-workers published a study investigating the mixture of two flavonolignans: 16 and 17. The mixture of the compounds was able to extend the lifespan of C. elegans depending on fgt-1 and daf-16. In a nematode model of Alzheimer's disease, the compound mixture led to a decrease in the disease progression.211 However, in this study silybin A and B were not tested.
High consumption of extra virgin olive oil (Olea europea L.) as the most representative food of the Mediterranean diet has been associated with longevity and reduced incidences of age-related diseases in many epidemiological and intervention studies.297–299 The triglyceride profile of olive oil with a particularly high proportion of esterified monounsaturated ω-9 fatty acids seems to be involved in these health-promoting effect.300 Supplementation of ω-9 fatty acids such as oleic acid can indeed promote longevity in C. elegans.301 However, this effect seems to be dependent on the test protocol, as other groups did not show this effect.302–304 Olive fruits and leaves also contain an interesting profile of phenolic secondary metabolites which are responsible for bitter and pungent notes in olive oil and include hydroxytyrosol (4), tyrosol (7), as well as the secoiridoids oleuropein (18), oleacein (19) and oleocanthal (20). 4, 7 and 18 have shown to improve C. elegans life- and health span in numerous studies dependent on daf-16, hsf-1 and skn-2.218,243,269,305–308 The secoiridoids 19 and 20 have shown beneficial effects against aging and senescence in human fibroblasts and D. melanogaster but have never been evaluated in C. elegans.309,310
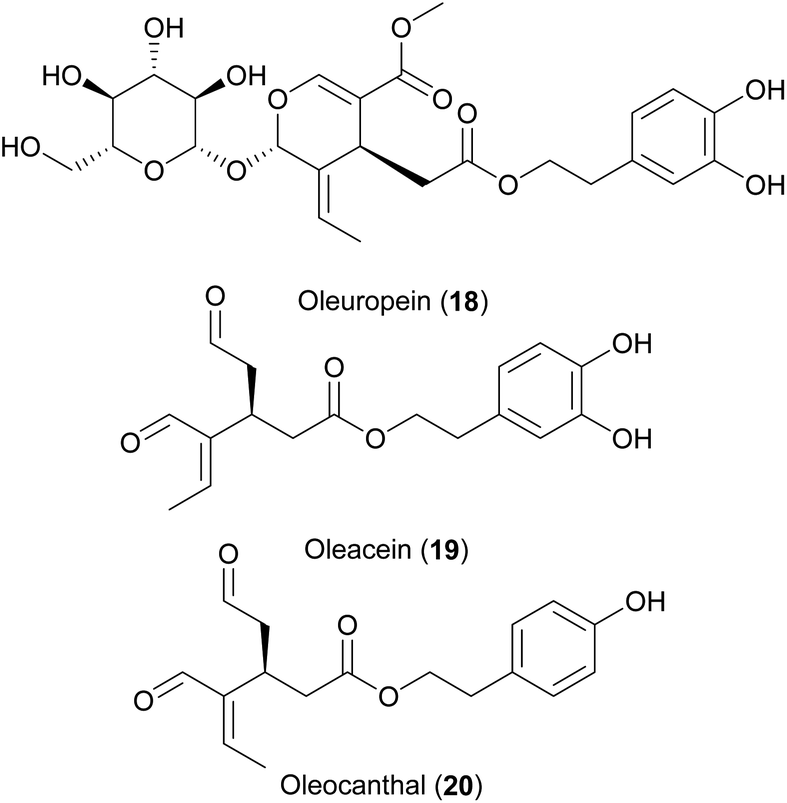
Betalains are water-soluble nitrogen pigments responsible for the colour of different plant parts of Caryophyllales. Due to their stability and safety they are popular food additives. Several medicinal plants of the order Caryophyllales are used in traditional medicine and their biological actions are sometimes attributed to the betalain constituents. Betanin (betacyanin, 21), the pigment of beetroots, indicaxanthin (betaxanthin, 22) present in Opuntia fruits, and 15 other betalains were investigated in C. elegans survival assays using the automated lifespan machine and in vivo antioxidative assays.120,311 Treatment with 25 μM of betalain for 48 h prior to the reproductive period could prolong lifespan up to 30%.311 These effects were accompanied by increased sir-2.1 and heat shock proteins expression. In response to betalain treatment, daf-16 was nuclearized in the transgene worms TJ365, and lifespan extension was abolished in mutant daf-16 worms. The two studies expand the alimentarium of lifespan prolonging NP with an established structure–activity relationship and mode of action.120 However, their experimental setup with treatment restricted to the larval stages complicates comparison with other studies, where it is rather standard to start the treatments in the L4 or adult stadium.
Recently cannabidiol (5) was discovered to influence the aging process of C. elegans. In the course of a toxicity screen, C. elegans was exposed to different concentrations of cannabidiol and it was revealed that instead of shortening, it increased lifespan up to 18% (at 40 μM). 5 doubled worm motility at old age, and increased their resilience to heat stress.312 Similar effects on C. elegans and D. rerio health span parameters were reported later.313–316 Especially, the age-associated decline of neuronal health was slowed by promotion of autophagy via sir-2.1, bec-1 and sqst-1. Whether the endocannabinoid system of the worm is involved in these effects remains to be clarified. The C. elegans cannabinoid receptor NPR-19 is sensitive to the conserved endocannabinoids 2-arachidonoylglycerol and anandamide. C. elegans endocannabinoids are involved in monoaminergic signalling and feeding behaviour,317,318 axon regeneration,319 development,320 and the coordination of nutrient status, metabolism and aging.321
4. Multicomponent mixtures from natural sources with anti-aging effects in C. elegans
We have gathered data on over 255 different NP extracts/fractions from 163 different plant, fungal and marine species probed in C. elegans for healthy aging between 2012 and 2023. The extracts are presented in the ESI† grouped into marine invertebrate- (Table ESI15†), fungal- (Table ESI16†), and plant- (Table ESI17†) sourced samples. The tables provide information on chemical aspects (species, organ, extraction solvent, type of characterization, and description of multicomponent mixture), and information on the experiments in C. elegans, important parameters (e.g., dose, strain), the findings of the respective studies and the report of signalling pathways or genes involved in the observed effect. Based on these data we classified the extracts regarding extraction solvent (Fig. 3A) and the level of chemical dissection of the multicomponent mixture (Fig. 3B).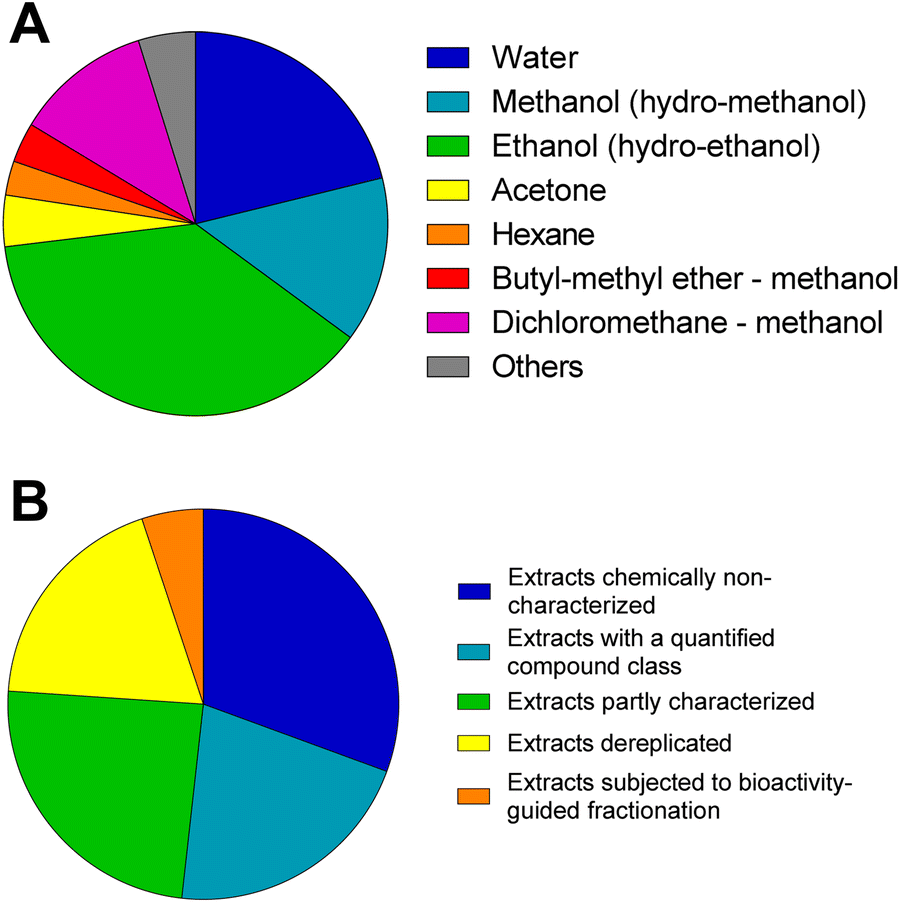 | ||
| Fig. 3 Analysis of literature dealing with multicomponent mixtures regarding (A) the used solvents for extract preparation (n = 208) and (B) their degree of analytical characterization (n = 255). | ||
Regarding extraction solvent, most studies used (hydro-) ethanol (40.0%) or (hydro-) methanol (13.9%) for extraction. 21.3% of all multi-component mixtures were aqueous extracts. Combined extracts with a broad polarity range of metabolites were also reported mainly using combined dichloromethane and methanol extracts (11.5%), as well as combined butyl-methyl ether and methanol extracts (3.4%). Only few studies test multi-component mixtures generated by midpolar and nonpolar solvents such as acetone (4.3%) and hexane (2.9%); ethyl acetate, dichloromethane or isopropanol are hardly represented in the literature.
Regarding the chemical profile, the multicomponent mixtures can be classified into five levels: (i) extracts chemically non-characterized, (ii) extracts with certain NP classes (e.g., total phenol, flavonoid, or anthocyanins) quantified with a spectrophotometer, (iii) extracts partly characterized with several annotated constituents and/or main constituents quantified, (iv) extract qualitatively characterized (e.g., with LC-MS(/MS), GC-MS, HPLC-DAD) to obtain a phytochemical profile, and (v) extracts subjected to bioactivity-guided fractionation. Nearly one third of the studies published between 2012 and 2023, do not provide a characterisation of the tested multicomponent mixtures (30.6%). This is critical, as it severely compromises the reproducibility and interpretation of results – an essential prerequisite for the progress of the research field. A considerable part of the studies performed provide some information about the total phenolic and total flavonoid contents using e.g., Folin–Ciocalteu assay (21.2%). 43.1% of the published literature contains at least one type of chromatographic analysis (e.g., LC-DAD, LC-MS/MS, or GC-MS). In most of these analyses, the phytochemical profile of the respective extract is further characterised by compound annotation using databases or single reference compounds for comparison. The level of characterisation differs significantly among the studies. In most cases the tested extracts have not been further chemically investigated to isolate the constituents contributing to the observed effect. Admittedly, it requires knowledge and instrumentation in analytics, chromatography and characterization of isolated compounds or collaboration partners covering these requirements. In general, the chemical analysis of bioactive extracts is often biased towards well-known antioxidants (e.g., investigation of the phenolic profile) or limited to well-known (and often well-investigated) chemical entities provided by suppliers. This generally inhibits research on novel, unexpected or minor compound classes. From the surveyed studies in this review it is obvious that extracts with already known potential radical scavenging in vitro activities are prioritized for C. elegans assays.57,322 However, approaches relying on in vitro antioxidant activities to screen for lifespan extending extracts has limitations. An extract screening study which included 30 different flower extracts concluded that the potency of in vitro radical scavenging activity shows no correlation to a prolongation of mean and median lifespan of nematodes.127
Regarding the experimental parameters for the C. elegans tests, we observed an extremely wide range of tested extract concentrations (0.1 μg ml−1 to 240 mg ml−1).177,187 The median tested concentration in C. elegans assays of 125 μg ml−1 is considerably higher than that reported for in vitro assays. About 30% of all tested concentrations were even above 10 mg ml−1 which raises questions on how these concentrations were achieved in the test media without precipitation, influencing pH and without causing osmotic stress on bacteria and worms. The validity of these experimental protocols and the reported effects have to be considered with caution.
4.1. Examples for investigated NPs
Among the investigated plant materials, a clear tendency towards edible plants can be observed. Especially from nutritional sciences, there is a huge interest in substantiating anti-aging claims of so called “super food”, nutraceuticals, and functional foods. In this respect many studies regarding anti-aging properties in C. elegans focus on the investigation of berries, e.g., açaí,169,257 goji,258 cranberries,259,260 blueberries,261 raspberries,262 black mulberries,263 and juniper berries.264 Interestingly, there is also a strong focus on botanicals consumed as beverages, e.g., tea (black,265 green,266 oolong,267 rooibos,268 and mate269–271 tea), guaraná,272 hops,273 and coffee.140,274 Another large portion of investigated extracts focuses on industrial waste products e.g., coffee silver skin,275 corncob - a by-product of the corn industry,276 different fruit peels (e.g., from apple277–279 or pomegranate280,281). On species level, the most studied plants (number of experiments per species) in recent years was Camellia sinensis,127,255,256,323–326 followed by Camellia tenuifolia,327Ginkgo biloba,187,328–330Lycium barbarum (goji)224,225,331 and Anacardium occidentale (cashew).230 Also species of the genus Vaccinium229,254,332–334 and Syzygium55,335,336 were repeatedly investigated.4.2. Following up studies of bioactive extracts
In a few studies, a bioactivity-guided isolation was performed using a C. elegans model for the bioactivity evaluation.257–259 These articles can be further divided into (i) studies that use C. elegans as an in vivo model for the confirmation of their in vitro results337,338 and (ii) studies where C. elegans is used as the primary model of bio-guided fractionation.339,340An example of the latter one is a study of Sayed and co-workers who compared the lifespan extending effects of Cuscuta chinensis and Eucommnia ulmoides extracts generated with a mixture of butyl-methyl ether and methanol (1![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :1) followed by an extraction with 100% methanol. Both extracts improved the resistance towards oxidative stress, and decreased the intracellular level of ROS.126 UPLC-Q-Exactive Orbitrap MS analysis allowed for metabolite identification and annotation. The authors also put efforts into unravelling the responsible single components. Hence, the extracts were fractionated via RP-HPLC and MPLC. 20 fractions of the E. ulmoides extract were again tested in the C. elegans heat stress assay. As a result, 7 fractions showed similar activity as the crude extract. 17 fractions of C. chinensis were further tested. From the extract of C. chinensis, enriched fractions containing astragalin (23, purity by 1H NMR ∼75%), pinoresinol (24, purity by 1H NMR ∼75%), and zingibroside R1 (25, purity by 1H NMR ∼50%) were obtained. The fraction from the C. chinensis extract enriched in zingibroside R1 improved the life span, the survival after heat stress, and the locomotion in a manner similar to the full C. chinensis extract and therefore zingibroside R1 could be (partly) responsible for the observed health benefits of C. chinensis. Furthermore, 2,3,4,5-tetrahydroxybenzoic acid (26) and the sterol lipid 4-α-formyl-stigmasta-7,24(241)dien-3-β-ol (27) are abundantly present in the C. chinensis extract and its most bioactive fraction.
:1) followed by an extraction with 100% methanol. Both extracts improved the resistance towards oxidative stress, and decreased the intracellular level of ROS.126 UPLC-Q-Exactive Orbitrap MS analysis allowed for metabolite identification and annotation. The authors also put efforts into unravelling the responsible single components. Hence, the extracts were fractionated via RP-HPLC and MPLC. 20 fractions of the E. ulmoides extract were again tested in the C. elegans heat stress assay. As a result, 7 fractions showed similar activity as the crude extract. 17 fractions of C. chinensis were further tested. From the extract of C. chinensis, enriched fractions containing astragalin (23, purity by 1H NMR ∼75%), pinoresinol (24, purity by 1H NMR ∼75%), and zingibroside R1 (25, purity by 1H NMR ∼50%) were obtained. The fraction from the C. chinensis extract enriched in zingibroside R1 improved the life span, the survival after heat stress, and the locomotion in a manner similar to the full C. chinensis extract and therefore zingibroside R1 could be (partly) responsible for the observed health benefits of C. chinensis. Furthermore, 2,3,4,5-tetrahydroxybenzoic acid (26) and the sterol lipid 4-α-formyl-stigmasta-7,24(241)dien-3-β-ol (27) are abundantly present in the C. chinensis extract and its most bioactive fraction.

Another example of a bioactivity-guided fractionation is the study of Jia and coworkers who followed up on a bioactive extract of clams (Meretrix meretrix). They discovered that a peptide-rich extract of M. meretrix could increase the survival of worms und oxidative stress conditions induced by paraquat.339 They separated the extract based on molecular weight into two fractions by ultrafiltration whereby the low-molecular weight peptides showed better activity in the paraquat stress survival assay. Thus, the fraction was further separated into five subfractions by gel filtration chromatography. The first fraction (F1) showed the best activity which decreased over the consecutive fractions. By RP-nano-LC-MS/MS they identified 25 peptides in F1 and subsequently synthesized and tested them. Three peptides were shown to be responsible for the observed effect which was further explained by an increased daf-16 dependent expression of sod-3, ctl-1 and ctl-2 after treatment.
4.3. C. elegans microbiome and natural products
Maintaining C. elegans in an unnatural but simplified and well-characterized culture with inactivated or dead OP50 as food sources is advantageous to directly probe the effects of NPs on worms. However, as in humans, in C. elegans its commensal microbial communities play a crucial physiological role.341 Some naturally associated bacteria influence behaviour, stress- and infection resistance, fecundity, metabolism, and life span of C. elegans;342–347e.g., different bacterial food sources can cause different uptake/accumulation of flavonoids.293 Maintenance of C. elegans on Bacillis subtilis strains present in its natural habitat increased lifespan by more than 50% and increased heat shock survival by more than 200% compared to standard OP50 diet.348It is well known that many NPs including dietary polyphenols interact with the human gut microbiome. Genuine NPs can be metabolized to different metabolites265 and vice versa NPs affect the composition of gut microbiota.349,350 Although C. elegans commensals are not identical to those of humans,351 the worm is proposed as a model for microbiome research.341 Recently an experimental microbiome modelling the native worm consisting of 14 bacterial strains was presented.352 Future studies will show how this microbiome interacts with natural products and how this affects worm health.
4.4. Intrinsic complexity of multicomponent NP mixtures and the challenge of combinatorial effects
For the elucidation of biological networks, the discipline of systems biology has integrated -omics approaches (e.g., genomics, transcriptomics, proteomics, and metabolomics) focussing on large-scale data deriving from C. elegans assays.353 However, to answer questions regarding the anti-aging effects of multi-component NPs on C. elegans, further disciplines need to be involved. From the NP side, the bottleneck is still the identification and elucidation of the bioactive principle on the molecular level – which compounds are responsible for certain phenotypic effects (e.g., the prolongation of life span). Multicomponent mixtures such as extracts generated from natural sources are known to contain hundreds of metabolites. Working with multicomponent mixtures thus requires holistic approaches on both the (phyto-)chemical as well as on the biological level. Regarding the screening of C. elegans, NP extracts are not often dissected in detail. Instead, specific (major) constituents known to be present in the respective mixture are exemplarily tested as representatives of the extract. Only in rare cases, the increase in lifespan observed for the single constituent is similar to that observed for the complex mixture, and thus only gives an incomplete picture.Moreover, NPs are particularly prone to exert combinatorial effects including an amplification of activity (when 1 + 1 > 2), potentiation (when 0 + 1 > 1), or antagonism (when 0 + 0 < 0). Another effect which can be observed is synergy (when 0 + 0 > 0). In this instance, the combination of multiple constituents is active, while the constituents separately are inactive. Apart from chemical challenges (e.g., poor chromatographic separation, analytical problems), these phenomena have to be considered.354,355 A possible case where additive effects may occur, might happen if some extract constituents inhibit efflux pumps, while others are genuinely active but not stable in the organism. In a recent case study in our group, it was found that the action of ostruthin, one of the major active constituents of the extract of masterwort (Peucedanum ostruthium), is enhanced by other constituents such as imperatorin and isoimperatorin, which are themselves inactive and presumably act by inhibiting xenobiotic defenses.302 Such additive effects are the proposed mechanism of many botanical drugs. Traditional phytotherapy not only makes use of molecular mixtures from one plant organ, but also employs complex mixtures of several plants potentiating the chemical complexity. This approach is very similar to Nature's which also employs complex mixtures of metabolites with distinct strategies to modulate biological processes. C. elegans might be a key model to unravel the therapeutic effect of such complexity in a living organism with possible implications for mammals too.
When following up on a bioactive extract, the goal should be to minimize the isolation of unwanted, inactive, or already well-investigated compounds. Traditionally, bioactivity-guided fractionation is one of the most common techniques applied to isolate and identify the bioactive principle of a multicomponent mixture. Successful examples, where this technique was used in combination with a C. elegans set-up were described before.322,327,329,338–340,356 Interestingly, further techniques to dissect anti-aging multicomponent NPs using C. elegans as model organism are scarce in the scientific literature.
Useful approaches might be high-performance liquid chromatography (HPLC)-based (micro)-fractionation,357 MS-based techniques such as molecular networking,358 or biochemometric approaches, where bioactivity data is correlated with chemical data from spectroscopy (e.g., NMR) or spectrometry (e.g., MS). One such example is the biochemometric approach ELINA (Eliciting Nature's Activities) which correlates activity with 1H NMR data to detect spectral features responsible for an observed effect.359–361 With this technique, it is possible to distinguish bioactives from inactives prior to isolation, as shown by the example of masterwort.362
5. Conclusions
An analysis of the literature shows that reports differ greatly in terms of the experimental focus. Many studies have elaborately established multiple assays to determine the function of the extracts not only on lifespan, but also on abiotic stress resistance, motility, proteotoxicity and more. Many mutant and transgenic C. elegans strains are widely established to unravel involved genes and pathways. However, a large part of studies does not deal with the chemical characterization or the isolation of constituents of the tested multicomponent mixtures. On the other hand, awareness has increased during the past years that extracts are chemically complex. Their composition can vary tremendously depending on the underlying material and its preparation. Accordingly, a chemical characterization of extracts used for any pharmacological investigation is mandatory.363There is also a strong bias towards well-studied structural NP classes such as flavonoids, stilbenes and tannins. This bias is not unusual in NP drug discovery. The same structures are also overrepresented in hit lists of in vitro bioactivity screenings. These compounds are often termed pan-assay interference compounds (PAINS) or invalid metabolic panaceas (IMPs).364,365 The PAINS term was originally coined for high-throughput screenings employing recombinant enzymes in which flavonoids, catechol and other phenols frequently inhibited enzyme activity through aggregation or oxidation rather than by specific interaction. Therefore, the PAINS concept cannot be translated to an in vivo model. However, there are indications that the effect of many phenols is also not a specific pharmacological effect. It is known that some polyphenols have moderate bactericidal effects,203,366,367 which might cause the lower availability of the bacterial nutritional source in C. elegans assays, at least in studies applying mg ml−1 doses of NPs. Thus, observed lifespan effects are likely to be caused by dietary restriction or the mitigation of bacterial colonization of the intestine in aged worms. Interestingly, many polyphenols are investigated for anthelminthic effects caused by tanning of cuticle, buccal cavity and intestine297 which at certain concentrations might also lead to a dietary restriction mediated lifespan increase. However, the strong focus of scientific research on polyphenols is impacted by their essential role in food, as well as their relatively easy acquisition; quite the opposite of more “exotic” or novel natural compounds. Nevertheless, efforts should be at least partially directed towards new structural classes for health span increase in C. elegans.
To achieve this goal, the authors highly recommend investigating promising extracts in more detail, e.g., through -omics studies. By introducing the term “wormomics”, a new workflow for the discovery of anti-aging NPs could be established. The nematodes have multiple advantages that allow for setting up experiments to discover (new) bioactive constituents of extracts prior to their isolation, e.g., by MS/MS molecular networking, NMR- or HPLC/MS-based bioactivity profiling and biochemometric approaches. These advantages can best be exploited by high content miniaturized screenings in multiwell-plates to test samples in parallel in a higher throughput, but also by the wide range of well-established assays that are increasingly automated. It is expected that extracts with health-promoting effects in C. elegans warrant further employment in particular with respect to the use of high resolution analytical techniques and big data analysis to provide insight into new chemistries and thus to unveil hidden treasures in complex mixtures.
6. Conflicts of interest
There are no conflicts to declare.7. Acknowledgements
The authors thank Anusha Shayegan for proofreading and Martina Redl for valuable discussions.8. References
- E. J. Stewart, R. Madden, G. Paul and F. Taddei, PLoS Biol., 2005, 3, e45 CrossRef PubMed.
- A. Currais, Ageing Res. Rev., 2017, 35, 297–300 CrossRef PubMed.
- R. Z. Moger-Reischer and J. T. Lennon, Nat. Rev. Microbiol., 2019, 17, 679–690 CrossRef CAS PubMed.
- G. Bjørklund, M. Shanaida, R. Lysiuk, M. Butnariu, M. Peana, I. Sarac, O. Strus, K. Smetanina and S. Chirumbolo, Molecules, 2022, 27, 7084 CrossRef PubMed.
- S. I. S. Rattan, Biogerontology, 2013, 14, 673–677 CrossRef PubMed.
- C. J. Kenyon, Nature, 2010, 464, 504–512 CrossRef CAS PubMed.
- M. R. Klass, Mech. Ageing Dev., 1983, 22, 279–286 CrossRef CAS PubMed.
- C. Kenyon, J. Chang, E. Gensch, A. Rudner and R. Tabtiang, Nature, 1993, 366, 461–464 CrossRef CAS PubMed.
- D. B. Friedman and T. E. Johnson, Genetics, 1988, 118, 75–86 CrossRef CAS PubMed.
- Y. Suh, G. Atzmon, M. O. Cho, D. Hwang, B. Liu, D. J. Leahy, N. Barzilai and P. Cohen, Proc. Natl. Acad. Sci. U. S. A., 2008, 105, 3438–3442 CrossRef CAS PubMed.
- B. J. Willcox, T. A. Donlon, Q. He, R. Chen, J. S. Grove, K. Yano, K. H. Masaki, D. C. Willcox, B. Rodriguez and J. D. Curb, Proc. Natl. Acad. Sci. U. S. A., 2008, 105, 13987–13992 CrossRef CAS PubMed.
- L. Pawlikowska, D. Hu, S. Huntsman, A. Sung, C. Chu, J. Chen, A. H. Joyner, N. J. Schork, W. C. Hsueh, A. P. Reiner, B. M. Psaty, G. Atzmon, N. Barzilai, S. R. Cummings, W. S. Browner, P. Y. Kwok and E. Ziv, Aging Cell, 2009, 8, 460–472 CrossRef CAS PubMed.
- M. Holzenberger, J. Dupont, B. Ducos, P. Leneuve, A. Géloën, P. C. Even, P. Cervera and Y. Le Bouc, Nature, 2003, 421, 182–187 CrossRef CAS PubMed.
- L. Guarente and C. Kenyon, Nature, 2000, 408, 255–262 CrossRef CAS PubMed.
- A. Martin-Montalvo, E. M. Mercken, S. J. Mitchell, H. H. Palacios, P. L. Mote, M. Scheibye-Knudsen, A. P. Gomes, T. M. Ward, R. K. Minor, M. J. Blouin, M. Schwab, M. Pollak, Y. Zhang, Y. Yu, K. G. Becker, V. A. Bohr, D. K. Ingram, D. A. Sinclair, N. S. Wolf, S. R. Spindler, M. Bernier and R. de Cabo, Nat. Commun., 2013, 4, 2192 CrossRef PubMed.
- R. Strong, R. A. Miller, A. Antebi, C. M. Astle, M. Bogue, M. S. Denzel, E. Fernandez, K. Flurkey, K. L. Hamilton, D. W. Lamming, M. A. Javors, J. P. de Magalhães, P. A. Martinez, J. M. McCord, B. F. Miller, M. Müller, J. F. Nelson, J. Ndukum, G. E. Rainger, A. Richardson, D. M. Sabatini, A. B. Salmon, J. W. Simpkins, W. T. Steegenga, N. L. Nadon and D. E. Harrison, Aging Cell, 2016, 15, 872–884 CrossRef CAS PubMed.
- R. Strong, R. A. Miller, C. J. Cheng, J. F. Nelson, J. Gelfond, S. K. Allani, V. Diaz, A. O. Dorigatti, J. Dorigatti, E. Fernandez, A. Galecki, B. Ginsburg, K. L. Hamilton, M. A. Javors, K. Kornfeld, M. Kaeberlein, S. Kumar, D. B. Lombard, M. Lopez-Cruzan, B. F. Miller, P. Rabinovitch, P. Reifsnyder, N. A. Rosenthal, M. A. Bogue, A. B. Salmon, Y. Suh, E. Verdin, H. Weissbach, J. Newman, F. Maccchiarini and D. E. Harrison, Aging Cell, 2022, 21, e13724 CrossRef CAS PubMed.
- D. E. Harrison, R. Strong, Z. D. Sharp, J. F. Nelson, C. M. Astle, K. Flurkey, N. L. Nadon, J. E. Wilkinson, K. Frenkel, C. S. Carter, M. Pahor, M. A. Javors, E. Fernandez and R. A. Miller, Nature, 2009, 460, 392–395 CrossRef CAS PubMed.
- J. P. de Magalhães, M. Stevens and D. Thornton, Trends Biotechnol., 2017, 35, 1062–1073 CrossRef PubMed.
- WHO, World Health Statistics Overview 2019: Monitoring Health for the SDGs, Sustainable Development Goals, 2019, https://apps.who.int/iris/bitstream/handle/10665/311696/WHO-DAD-2019.1-eng.pdf, accessed 13.01.2023 Search PubMed.
- J. Oeppen and J. W. Vaupel, Science, 2002, 296, 1029–1031 CrossRef CAS PubMed.
- A. J. Scott, M. Ellison and D. A. Sinclair, Nat. Aging, 2021, 1, 616–623 CrossRef PubMed.
- Y.-C. Tsai, L.-H. Cheng, Y.-W. Liu, O.-J. Jeng and Y.-K. Lee, Biosci. Microbiota, Food Health, 2021, 40, 1–11 CrossRef CAS PubMed.
- C. Ermogenous, C. Green, T. Jackson, M. Ferguson and J. M. Lord, Drug Discovery Today, 2020, 25, 1403–1415 CrossRef CAS PubMed.
- K. A. Steinkraus, E. D. Smith, C. Davis, D. Carr, W. R. Pendergrass, G. L. Sutphin, B. K. Kennedy and M. Kaeberlein, Aging Cell, 2008, 7, 394–404 CrossRef CAS PubMed.
- E. Cohen, J. Bieschke, R. M. Perciavalle, J. W. Kelly and A. Dillin, Science, 2006, 313, 1604–1610 CrossRef CAS PubMed.
- N. Arantes-Oliveira, J. R. Berman and C. Kenyon, Science, 2003, 302, 611 CrossRef CAS PubMed.
- C. Selman, S. Lingard, A. I. Choudhury, R. L. Batterham, M. Claret, M. Clements, F. Ramadani, K. Okkenhaug, E. Schuster and E. Blanc, FASEB J., 2008, 22, 807–818 CrossRef CAS PubMed.
- J. A. Rollins, A. C. Howard, S. K. Dobbins, E. H. Washburn and A. N. Rogers, J. Gerontol., 2017, 72, 473–480 CrossRef CAS PubMed.
- J. R. Beard, A. Officer, I. A. de Carvalho, R. Sadana, A. M. Pot, J.-P. Michel, P. Lloyd-Sherlock, J. E. Epping-Jordan, G. M. E. E. Peeters, W. R. Mahanani, J. A. Thiyagarajan and S. Chatterji, Lancet, 2016, 387, 2145–2154 CrossRef PubMed.
- B. K. Kennedy, S. L. Berger, A. Brunet, J. Campisi, A. M. Cuervo, E. S. Epel, C. Franceschi, G. J. Lithgow, R. I. Morimoto, J. E. Pessin, T. A. Rando, A. Richardson, E. E. Schadt, T. Wyss-Coray and F. Sierra, Cell, 2014, 159, 709–713 CrossRef CAS PubMed.
- A. Yabluchanskiy, Z. Ungvari, A. Csiszar and S. Tarantini, Physiol. Int., 2018, 105, 298–308 CAS.
- D. R. Seals, J. N. Justice and T. J. LaRocca, J. Physiol., 2016, 594, 2001–2024 CrossRef CAS PubMed.
- C. López-Otín, M. A. Blasco, L. Partridge, M. Serrano and G. Kroemer, Cell, 2013, 153, 1194–1217 CrossRef PubMed.
- L. M. DeVito, N. Barzilai, A. M. Cuervo, L. J. Niedernhofer, S. Milman, M. Levine, D. Promislow, L. Ferrucci, G. A. Kuchel, J. Mannick, J. Justice, M. M. Gonzales, J. L. Kirkland, P. Cohen and J. Campisi, Ann. N. Y. Acad. Sci., 2022, 1507, 70–83 CrossRef PubMed.
- J. N. Justice, L. Niedernhofer, P. D. Robbins, V. R. Aroda, M. A. Espeland, S. B. Kritchevsky, G. A. Kuchel and N. Barzilai, Cardiovasc. Endocrinol. Metab., 2018, 7, 80–83 CrossRef PubMed.
- S. I. Rattan, Biogerontology, 2013, 14, 673–677 CrossRef PubMed.
- V. Codd, C. P. Nelson, E. Albrecht, M. Mangino, J. Deelen, J. L. Buxton, J. J. Hottenga, K. Fischer, T. Esko, I. Surakka, L. Broer, D. R. Nyholt, I. Mateo Leach, P. Salo, S. Hägg, M. K. Matthews, J. Palmen, G. D. Norata, P. F. O'Reilly, D. Saleheen, N. Amin, A. J. Balmforth, M. Beekman, R. A. de Boer, S. Böhringer, P. S. Braund, P. R. Burton, A. J. de Craen, M. Denniff, Y. Dong, K. Douroudis, E. Dubinina, J. G. Eriksson, K. Garlaschelli, D. Guo, A. L. Hartikainen, A. K. Henders, J. J. Houwing-Duistermaat, L. Kananen, L. C. Karssen, J. Kettunen, N. Klopp, V. Lagou, E. M. van Leeuwen, P. A. Madden, R. Mägi, P. K. Magnusson, S. Männistö, M. I. McCarthy, S. E. Medland, E. Mihailov, G. W. Montgomery, B. A. Oostra, A. Palotie, A. Peters, H. Pollard, A. Pouta, I. Prokopenko, S. Ripatti, V. Salomaa, H. E. Suchiman, A. M. Valdes, N. Verweij, A. Viñuela, X. Wang, H. E. Wichmann, E. Widen, G. Willemsen, M. J. Wright, K. Xia, X. Xiao, D. J. van Veldhuisen, A. L. Catapano, M. D. Tobin, A. S. Hall, A. I. Blakemore, W. H. van Gilst, H. Zhu, J. Erdmann, M. P. Reilly, S. Kathiresan, H. Schunkert, P. J. Talmud, N. L. Pedersen, M. Perola, W. Ouwehand, J. Kaprio, N. G. Martin, C. M. van Duijn, I. Hovatta, C. Gieger, A. Metspalu, D. I. Boomsma, M. R. Jarvelin, P. E. Slagboom, J. R. Thompson, T. D. Spector, P. van der Harst and N. J. Samani, Nat. Genet., 2013, 45, 422–427 CrossRef CAS PubMed.
- B. A. Benayoun, E. A. Pollina and A. Brunet, Nat. Rev. Mol. Cell Biol., 2015, 16, 593–610 CrossRef CAS PubMed.
- D. Gems and J. P. de Magalhães, Ageing Res. Rev., 2021, 70, 101407 CrossRef PubMed.
- D. F. Lawler, B. T. Larson, J. M. Ballam, G. K. Smith, D. N. Biery, R. H. Evans, E. H. Greeley, M. Segre, H. D. Stowe and R. D. Kealy, Br. J. Nutr., 2008, 99, 793–805 CrossRef CAS PubMed.
- J. A. Mattison, R. J. Colman, T. M. Beasley, D. B. Allison, J. W. Kemnitz, G. S. Roth, D. K. Ingram, R. Weindruch, R. de Cabo and R. M. Anderson, Nat. Commun., 2017, 8, 14063 CrossRef CAS PubMed.
- M. Blüher, B. B. Kahn and C. R. Kahn, Science, 2003, 299, 572–574 CrossRef PubMed.
- H. Kurosu, M. Yamamoto, J. D. Clark, J. V. Pastor, A. Nandi, P. Gurnani, O. P. McGuinness, H. Chikuda, M. Yamaguchi, H. Kawaguchi, I. Shimomura, Y. Takayama, J. Herz, C. R. Kahn, K. P. Rosenblatt and M. Kuro-o, Science, 2005, 309, 1829–1833 CrossRef CAS PubMed.
- R. A. Miller, D. E. Harrison, C. M. Astle, J. A. Baur, A. R. Boyd, R. de Cabo, E. Fernandez, K. Flurkey, M. A. Javors, J. F. Nelson, C. J. Orihuela, S. Pletcher, Z. D. Sharp, D. Sinclair, J. W. Starnes, J. E. Wilkinson, N. L. Nadon and R. Strong, J. Gerontol., Ser. A, 2011, 66, 191–201 CrossRef PubMed.
- F. Sierra, Cold Spring Harbor Perspect. Med., 2016, 6, a025163 CrossRef PubMed.
- B. M. Egan, A. Scharf, F. Pohl and K. Kornfeld, Front. Pharmacol., 2022, 13, 938650 CrossRef CAS PubMed.
- D. J. Newman and G. M. Cragg, J. Nat. Prod., 2020, 83, 770–803 CrossRef CAS PubMed.
- Z. Wu, J. V. Smith, V. Paramasivam, P. Butko, I. Khan, J. R. Cypser and Y. Luo, Cell. Mol. Biol., 2002, 48, 725–731 CAS.
- J. A. Joseph, B. Shukitt-Hale, N. A. Denisova, R. L. Prior, G. Cao, A. Martin, G. Taglialatela and P. C. Bickford, J. Neurosci., 1998, 18, 8047–8055 CrossRef CAS PubMed.
- M. A. Wilson, B. Shukitt-Hale, W. Kalt, D. K. Ingram, J. A. Joseph and C. A. Wolkow, Aging Cell, 2006, 5, 59–68 CrossRef CAS PubMed.
- N. Saul, S. Möller, F. Cirulli, A. Berry, W. Luyten and G. Fuellen, Biogerontology, 2021, 22, 215–236 CrossRef PubMed.
- H. van Hattum and H. Waldmann, J. Am. Chem. Soc., 2014, 136, 11853–11859 CrossRef CAS PubMed.
- S. C. Sukuru, J. L. Jenkins, R. E. Beckwith, J. Scheiber, A. Bender, D. Mikhailov, J. W. Davies and M. Glick, J. Biomol. Screening, 2009, 14, 690–699 CrossRef CAS PubMed.
- B. Squiban and C. L. Kurz, Curr. Drug Targets, 2011, 12, 967–977 CrossRef CAS PubMed.
- M. P. de Torre, R. Y. Cavero, M. I. Calvo and J. L. Vizmanos, Antioxidants, 2019, 8, 142 CrossRef PubMed.
- J. Chen, J. Zhang, Y. Xiang, L. Xiang, Y. Liu, X. He, X. Zhou, X. Liu and Z. Huang, Food Funct., 2016, 7, 943–952 RSC.
- E. S. David, in Bioactive Compounds in Phytomedicine, ed. R. Iraj, IntechOpen, Rijeka, 2012, ch. 8, DOI:10.5772/26102.
- J. A. Joseph, B. Shukitt-Hale, N. A. Denisova, D. Bielinski, A. Martin, J. J. McEwen and P. C. Bickford, J. Neurosci., 1999, 19, 8114–8121 CrossRef CAS PubMed.
- S. Suter and M. Lucock, Explor. Res. Hypothesis Med., 2017, 2, 1–7 CrossRef.
- A. Argyropoulou, N. Aligiannis, I. P. Trougakos and A. L. Skaltsounis, Nat. Prod. Rep., 2013, 30, 1412–1437 RSC.
- K. Pallauf, N. Duckstein and G. Rimbach, Proc. Natl. Acad. Sci. U.S.A, 2017, 76, 145–162 Search PubMed.
- A. J. Ding, S. Q. Zheng, X. B. Huang, T. K. Xing, G. S. Wu, H. Y. Sun, S. H. Qi and H. R. Luo, Nat. Prod. Bioprospect., 2017, 7, 335–404 CrossRef CAS PubMed.
- K. Matsunami, Front. Nutr., 2018, 5, 111 CrossRef PubMed.
- D. Chattopadhyay and K. Thirumurugan, Mech. Ageing Dev., 2018, 171, 47–57 CrossRef PubMed.
- L. Wang, X. Zuo, Z. Ouyang, P. Qiao and F. Wang, J. Evidence-Based Complementary Altern. Med., 2021, 5591573 Search PubMed.
- Y. Ye, Q. Gu and X. Sun, Compr. Rev. Food Sci. Food Saf., 2020, 19, 3084–3105 CrossRef PubMed.
- J. Martel, C.-Y. Wu, H.-H. Peng, Y.-F. Ko, H.-C. Yang, J. D. Young and D. M. Ojcius, Microb. Cell, 2020, 7, 255–269 CrossRef CAS PubMed.
- M. D. Navarro-Hortal, J. M. Romero-Márquez, S. Osta, V. Jiménez-Trigo, P. Muñoz-Ollero and A. Varela-López, Diseases, 2022, 10, 28 CrossRef CAS PubMed.
- P. Shen, Y. Yue and Y. Park, Crit. Rev. Food Sci. Nutr., 2018, 58, 741–754 CrossRef PubMed.
- P. Shen, Y. Yue, J. Zheng and Y. Park, Annu. Rev. Food Sci. Technol., 2018, 9, 1–22 CrossRef PubMed.
- A. Zhu, F. Zheng, W. Zhang, L. Li, Y. Li, H. Hu, Y. Wu, W. Bao, G. Li, Q. Wang and H. Li, Antioxidants, 2022, 11, 705 CrossRef CAS PubMed.
- M. Greener, Prescriber, 2021, 32, 29–32 CrossRef.
- L. P. O'Reilly, C. J. Luke, D. H. Perlmutter, G. A. Silverman and S. C. Pak, Adv. Drug Delivery Rev., 2014, 69, 247–253 CrossRef PubMed.
- J. Zwirchmayr, B. Kirchweger, T. Lehner, A. Tahir, D. Pretsch and J. M. Rollinger, Sci. Rep., 2020, 10, 12323 CrossRef CAS PubMed.
- S. Brenner, Genetics, 1974, 77, 71–94 CrossRef CAS PubMed.
- P. M. Meneely, C. L. Dahlberg and J. K. Rose, Curr. Protoc. Essent. Lab. Tech., 2019, 19, e35 Search PubMed.
- I. Antoshechkin and P. W. Sternberg, Nat. Rev. Genet., 2007, 8, 518–532 CrossRef CAS PubMed.
- T. Kaletta and M. O. Hengartner, Nat. Rev. Drug Discovery, 2006, 5, 387–398 CrossRef CAS PubMed.
- W. Kim, R. S. Underwood, I. Greenwald and D. D. Shaye, Genetics, 2018, 210, 445–461 CrossRef CAS PubMed.
- B. Chakravarty, Nutr. Res., 2022, 106, 47–59 CrossRef CAS PubMed.
- S. E. Hulme and G. M. Whitesides, Angew. Chem., Int. Ed., 2011, 50, 4774–4807 CrossRef CAS PubMed.
- S. J. Husson, S. Moyson, D. Valkenborg, G. Baggerman and I. Mertens, Biochem. Biophys. Res. Commun., 2015, 468, 519–524 CrossRef CAS PubMed.
- K. Horvath, D. Angeletti, G. Nascetti and C. Carere, Ann. Ist. Super. Sanita, 2013, 49, 9–17 Search PubMed.
- A. Desalermos, M. Muhammed, J. Glavis-Bloom and E. Mylonakis, Expert Opin. Drug Discovery, 2011, 6, 645–652 CrossRef CAS PubMed.
- B. S. Samuel, H. Rowedder, C. Braendle, M.-A. Félix and G. Ruvkun, Proc. Natl. Acad. Sci. U.S.A, 2016, 113, E3941–E3949 CAS.
- M.-A. Félix and F. Duveau, BMC Biol., 2012, 10, 59 CrossRef PubMed.
- D. H. Hall and Z. F. Altun, C. elegans Atlas, Cold Spring Harbor Laboratory Press, 2007 Search PubMed.
- L. R. Baugh, Genetics, 2013, 194, 539–555 CrossRef CAS PubMed.
- J. J. Stastna, A. D. Yiapanas, A. A. Mandawala, K. E. Fowler and S. C. Harvey, Cryobiology, 2020, 92, 86–91 CrossRef CAS PubMed.
- M. Porta-de-la-Riva, L. Fontrodona, A. Villanueva and J. Cerón, J. Visualized Exp., 2012, 64, e4019 Search PubMed.
- H. B. Atakan, F. Ayhan and M. A. M. Gijs, Lab Chip, 2020, 20, 155–167 RSC.
- H. Xiong, C. Pears and A. Woollard, Sci. Rep., 2017, 7, 9839 CrossRef PubMed.
- A. P. Page and I. Johnstone, WormBook, 2007, DOI:10.1895/wormbook.1.138.1.
- B.-M. Song and L. Avery, Worm, 2013, 2, e21833 CrossRef PubMed.
- D. Raizen, B. M. Song, N. Trojanowski and Y. J. You, WormBook, 2012, DOI:10.1895/wormbook.1.154.1.
- K. S. Lee, S. Iwanir, R. B. Kopito, M. Scholz, J. A. Calarco, D. Biron and E. Levine, Nat. Commun., 2017, 8, 14221 CrossRef CAS PubMed.
- J. D. McGhee, Wiley Interdiscip. Rev.: Dev. Biol., 2013, 2, 347–367 CrossRef CAS PubMed.
- M. Ezcurra, A. Benedetto, T. Sornda, A. F. Gilliat, C. Au, Q. Zhang, S. van Schelt, A. L. Petrache, H. Wang, Y. de la Guardia, S. Bar-Nun, E. Tyler, M. J. Wakelam and D. Gems, Curr. Biol., 2018, 28, 2544–2556 CrossRef CAS PubMed.
- A. Buis, S. Bellemin, J. Goudeau, L. Monnier, N. Loiseau, H. Guillou and H. Aguilaniu, Cell Rep., 2019, 28, 1041–1049 CrossRef CAS PubMed.
- D. G. Moerman and A. Fire, in C. elegansII, ed. D. L. Riddle, T. Blumenthal, B. J. Meyer and J. R. Priess, Cold Spring Harbor Laboratory Press Copyright © 1997, Cold Spring Harbor Laboratory Press, Cold Spring Harbor (NY), 1997 Search PubMed.
- A. L. Kauffman, J. M. Ashraf, M. R. Corces-Zimmerman, J. N. Landis and C. T. Murphy, PLoS Biol., 2010, 8, e1000372 CrossRef PubMed.
- T. Sornda, M. Ezcurra, C. Kern, E. R. Galimov, C. Au, Y. de la Guardia and D. Gems, J. Gerontol., 2019, 74, 1180–1188 CrossRef CAS PubMed.
- C. C. Kern, S. Townsend, A. Salzmann, N. B. Rendell, G. W. Taylor, R. M. Comisel, L. C. Foukas, J. Bähler and D. Gems, Nat. Commun., 2021, 12, 5801 CrossRef CAS PubMed.
- T. Komura, M. Yamanaka, K. Nishimura, K. Hara and Y. Nishikawa, npj Aging Mech. Dis., 2021, 7, 12 CrossRef CAS PubMed.
- L. E, T. Zhou, S. Koh, M. Chuang, R. Sharma, N. Pujol, A. D. Chisholm, C. Eroglu, H. Matsunami and D. Yan, Neuron, 2018, 97, 125–138 CrossRef CAS PubMed.
- C. J. Smith, J. D. Watson, W. C. Spencer, T. O'Brien, B. Cha, A. Albeg, M. Treinin and D. M. Miller, Dev. Biol., 2010, 345, 18–33 CrossRef CAS PubMed.
- L. A. Herndon, P. J. Schmeissner, J. M. Dudaronek, P. A. Brown, K. M. Listner, Y. Sakano, M. C. Paupard, D. H. Hall and M. Driscoll, Nature, 2002, 419, 808–814 CrossRef CAS PubMed.
- D. K. Chow, C. F. Glenn, J. L. Johnston, I. G. Goldberg and C. A. Wolkow, Exp. Gerontol., 2006, 41, 252–260 CrossRef CAS PubMed.
- C. Huang, C. Xiong and K. Kornfeld, Proc. Natl. Acad. Sci. U.S.A, 2004, 101, 8084–8089 CrossRef CAS PubMed.
- J. J. Collins, C. Huang, S. Hughes and K. Kornfeld, in Wormbook, 2007, pp. 1–21 Search PubMed.
- M. D. McGee, D. Weber, N. Day, C. Vitelli, D. Crippen, L. A. Herndon, D. H. Hall and S. Melov, Aging Cell, 2011, 10, 699–710 CrossRef CAS PubMed.
- L. Herndon, C. Wolkow, M. Driscoll and D. Hall, in WormAtlas, 2017, pp. 9–39, DOI:10.1007/978-3-319-44703-2_2.
- C. Coburn, E. Allman, P. Mahanti, A. Benedetto, F. Cabreiro, Z. Pincus, F. Matthijssens, C. Araiz, A. Mandel, M. Vlachos, S. A. Edwards, G. Fischer, A. Davidson, R. E. Pryor, A. Stevens, F. J. Slack, N. Tavernarakis, B. P. Braeckman, F. C. Schroeder, K. Nehrke and D. Gems, PLoS Biol., 2013, 11, e1001613 CrossRef CAS PubMed.
- L. Herndon, C. Wolkow, M. Driscoll and D. Hall, in WormAtlas, 2018, DOI:10.3908/wormatlas.8.4.
- L. Gonzalez-Moragas, A. Roig and A. Laromaine, Adv. Colloid Interface Sci., 2015, 219, 10–26 CrossRef CAS PubMed.
- H.-E. H. Park, Y. Jung and S.-J. V. Lee, Mol. Cells, 2017, 40, 90–99 CrossRef CAS PubMed.
- F. R. G. Amrit, R. Ratnappan, S. A. Keith and A. Ghazi, Methods, 2014, 68, 465–475 CrossRef CAS PubMed.
- N. Stroustrup, B. E. Ulmschneider, Z. M. Nash, I. F. López-Moyado, J. Apfeld and W. Fontana, Nat. Methods, 2013, 10, 665–670 CrossRef CAS PubMed.
- M. A. Guerrero-Rubio, S. Hernandez-Garcia, J. Escribano, M. Jimenez-Atienzar, J. Cabanes, F. Garcia-Carmona and F. Gandia-Herrero, Food Chem., 2020, 330, 127228 CrossRef CAS PubMed.
- B. Xian, J. Shen, W. Chen, N. Sun, N. Qiao, D. Jiang, T. Yu, Y. Men, Z. Han, Y. Pang, M. Kaeberlein, Y. Huang and J.-D. J. Han, Aging Cell, 2013, 12, 398–409 CrossRef CAS PubMed.
- D. P. Felker, C. E. Robbins and M. A. McCormick, Transl. Med. Aging, 2020, 4, 1–10 CrossRef PubMed.
- J.-H. Hahm, S. Kim, R. DiLoreto, C. Shi, S.-J. V. Lee, C. T. Murphy and H. G. Nam, Nat. Commun., 2015, 6, 8919 CrossRef CAS PubMed.
- A. Jushaj, M. Churgin, B. Yao, M. De La Torre, C. Fang-Yen and L. Temmerman, PLoS One, 2020, 15, e0229583 CrossRef CAS PubMed.
- H. N. Kim, H. W. Seo, B. S. Kim, H. J. Lim, H. N. Lee, J. S. Park, Y. J. Yoon, J. W. Oh, M. J. Oh, J. Kwon, C. H. Oh, D. S. Cha and H. Jeon, Nat. Prod. Sci., 2015, 21, 128–133 CAS.
- S. M. A. Sayed, S. Alseekh, K. Siems, A. R. Fernie, W. Luyten, C. Schmitz-Linneweber and N. Saul, Nutrients, 2022, 14, 4199 CrossRef CAS PubMed.
- Q. Chen, X. Yang, E. Capanoglu, A. T. Amrouche, L. Wu, J. Luo, Y. Zhu, Y. Wang, X. Jiang, D. Zhang and B. Lu, Food Funct., 2023, 14, 457–470 RSC.
- Y.-X. Chen, P. T. N. Le, T.-T. Tzeng, T.-H. Tran, A. T. Nguyen, I. H.-J. Cheng, C.-Y. F. Huang, Y.-J. Shiao and T.-T. Ching, Nutrients, 2021, 13, 4317 CrossRef CAS PubMed.
- K. Koch, N. Weldle, S. Baier, C. Buechter and W. Waetjen, Eur. J. Nutr., 2020, 59, 137–150 CrossRef CAS PubMed.
- W. Zhang, B. Zheng, N. Deng, H. Wang, T. Li and R. H. Liu, J. Food Sci., 2020, 85, 4367–4376 CrossRef CAS PubMed.
- S. Jiang, N. Deng, B. Zheng, T. Li and R. H. Liu, Food Funct., 2021, 12, 4471–4483 RSC.
- H. Wang, S. Zhang, L. Zhai, L. Sun, D. Zhao, Z. Wang and X. Li, Food Funct., 2021, 12, 6793–6808 RSC.
- M. Tao, R. Li, T. Xu, Z. Zhang, T. Wu, S. Pan and X. Xu, Food Funct., 2021, 12, 8196–8207 RSC.
- R. J. McCloskey, A. D. Fouad, M. A. Churgin and C. Fang-Yen, J. Neurophysiol., 2017, 117, 1911–1934 CrossRef PubMed.
- S. Dernovici, T. Starc, J. A. Dent and P. Ribeiro, Dev. Neurobiol., 2007, 67, 189–204 CrossRef CAS PubMed.
- B. Kirchweger, L. C. Klein-Junior, D. Pretsch, Y. Chen, S. Cretton, A. L. Gasper, Y. V. Heyden, P. Christen, J. Kirchmair, A. T. Henriques and J. M. Rollinger, Front. Neurosci., 2022, 16, 826289 CrossRef PubMed.
- M. A. Bolanowski, R. L. Russell and L. A. Jacobson, Mech. Ageing Dev., 1981, 15, 279–295 CrossRef CAS PubMed.
- M. Scholz, D. J. Lynch, K. S. Lee, E. Levine and D. Biron, J. Neurosci. Methods, 2016, 274, 172–178 CrossRef PubMed.
- S. R. Lockery, S. E. Hulme, W. M. Roberts, K. J. Robinson, A. Laromaine, T. H. Lindsay, G. M. Whitesides and J. C. Weeks, Lab Chip, 2012, 12, 2211–2220 RSC.
- D. Ryu, L. Mouchiroud, P. A. Andreux, E. Katsyuba, N. Moullan, A. A. Nicolet-Dit-Félix, E. G. Williams, P. Jha, G. Lo Sasso, D. Huzard, P. Aebischer, C. Sandi, C. Rinsch and J. Auwerx, Nat. Med., 2016, 22, 879–888 CrossRef CAS PubMed.
- P. J. Campagnola, A. C. Millard, M. Terasaki, P. E. Hoppe, C. J. Malone and W. A. Mohler, Biophys. J., 2002, 82, 493–508 CrossRef CAS PubMed.
- L. Amigoni, M. Stuknyte, C. Ciaramelli, C. Magoni, I. Bruni, I. De Noni, C. Airoldi, M. E. Regonesi and A. Palmioli, J. Funct. Foods, 2017, 33, 297–306 CrossRef CAS.
- L. A. Harrington and C. B. Harley, Mech. Ageing Dev., 1988, 43, 71–78 CrossRef CAS PubMed.
- E. de Castro, S. Hegi de Castro and T. E. Johnson, Free Radical Biol. Med., 2004, 37, 139–145 CrossRef CAS PubMed.
- G. J. Lithgow, T. M. White, S. Melov and T. E. Johnson, Proc. Natl. Acad. Sci. U.S.A, 1995, 92, 7540–7544 CrossRef CAS PubMed.
- A. Kampkötter, T. Pielarski, R. Rohrig, C. Timpel, Y. Chovolou, W. Wätjen and R. Kahl, Pharmacol. Res., 2007, 55, 139–147 CrossRef PubMed.
- S. Murakami and T. E. Johnson, Genetics, 1996, 143, 1207–1218 CrossRef CAS PubMed.
- N. Fischer, C. Buechter, K. Koch, S. Albert, R. Csuk and W. Waetjen, J. Pharm. Pharmacol., 2017, 69, 73–81 CrossRef CAS PubMed.
- C. Büchter, D. Ackermann, S. Havermann, S. Honnen, Y. Chovolou, G. Fritz, A. Kampkötter and W. Wätjen, Int. J. Mol. Sci., 2013, 14, 11895–11914 CrossRef PubMed.
- C. F. Labuschagne and A. B. Brenkman, Ageing Res. Rev., 2013, 12, 918–930 CrossRef CAS PubMed.
- T. J. Schulz, K. Zarse, A. Voigt, N. Urban, M. Birringer and M. Ristow, Cell Metab., 2007, 6, 280–293 CrossRef CAS PubMed.
- V. I. Pérez, A. Bokov, H. Van Remmen, J. Mele, Q. Ran, Y. Ikeno and A. Richardson, Biochim. Biophys. Acta, 2009, 1790, 1005–1014 CrossRef PubMed.
- M. P. Murphy, H. Bayir, V. Belousov, C. J. Chang, K. J. A. Davies, M. J. Davies, T. P. Dick, T. Finkel, H. J. Forman, Y. Janssen-Heininger, D. Gems, V. E. Kagan, B. Kalyanaraman, N.-G. Larsson, G. L. Milne, T. Nyström, H. E. Poulsen, R. Radi, H. Van Remmen, P. T. Schumacker, P. J. Thornalley, S. Toyokuni, C. C. Winterbourn, H. Yin and B. Halliwell, Nat. Metab., 2022, 4, 651–662 CrossRef PubMed.
- D.-E. Jeong, Y. Lee and S.-J. V. Lee, in Hypoxia: Methods and Protocols, ed. L. E. Huang, Springer New York, New York, 2018, pp. 213–225 Search PubMed.
- W. N. Tawe, M.-L. Eschbach, R. D. Walter and K. Henkle-Dührsen, Nucleic Acids Res., 1998, 26, 1621–1627 CrossRef CAS PubMed.
- P. Link and M. Wink, Phytomedicine, 2019, 55, 119–124 CrossRef CAS PubMed.
- P. Link, K. Roth, F. Sporer and M. Wink, Molecules, 2016, 21, 871 CrossRef PubMed.
- Z.-Z. Yang, Y.-T. Yu, H.-R. Lin, D.-C. Liao, X.-H. Cui and H.-B. Wang, Free Radical Biol. Med., 2018, 129, 310–322 CrossRef CAS PubMed.
- D. Pretsch, J. M. Rollinger, A. Schmid, M. Genov, T. Woehrer, L. Krenn, M. Moloney, A. Kasture, T. Hummel and A. Pretsch, Sci. Rep., 2020, 10, 11707 CrossRef CAS PubMed.
- S. T. Henderson and T. E. Johnson, Curr. Biol., 2001, 11, 1975–1980 CrossRef CAS PubMed.
- S.-Q. Zheng, A.-J. Ding, G.-P. Li, G.-S. Wu and H.-R. Luo, PLoS One, 2013, 8, e56877 CrossRef CAS PubMed.
- A. P. García-González, A. D. Ritter, S. Shrestha, E. C. Andersen, L. S. Yilmaz and A. J. M. Walhout, Cell, 2017, 169, 431–441 CrossRef PubMed.
- S. Beydoun, H. S. Choi, G. Dela-Cruz, J. Kruempel, S. Huang, D. Bazopoulou, H. A. Miller, M. L. Schaller, C. R. Evans and S. F. Leiser, Commun. Biol., 2021, 4, 258 CrossRef CAS PubMed.
- D. H. Kim, Annu. Rev. Genet., 2013, 47, 233–246 CrossRef CAS PubMed.
- I. Lenaerts, G. A. Walker, L. Van Hoorebeke, D. Gems and J. R. Vanfleteren, J. Gerontol., Ser. A, 2008, 63, 242–252 CrossRef PubMed.
- L. Larigot, D. Mansuy, I. Borowski, X. Coumoul and J. Dairou, Biomolecules, 2022, 12, 342 CrossRef CAS PubMed.
- T. H. Lindblom and A. K. Dodd, J. Exp. Zool., 2006, 305, 720–730 CrossRef PubMed.
- A. R. Burns, I. M. Wallace, J. Wildenhain, M. Tyers, G. Giaever, G. D. Bader, C. Nislow, S. R. Cutler and P. J. Roy, Nat. Chem. Biol., 2010, 6, 549–557 CrossRef CAS PubMed.
- A. AlOkda and J. M. Van Raamsdonk, MicroPubl., Biol., 2022 DOI:10.17912/micropub.biology.000634.
- X. Wang, X. Wang, L. Li and D. Wang, Biochem. Biophys. Res. Commun., 2010, 400, 613–618 CrossRef CAS PubMed.
- N. Wu, Y.-C. Ma, X.-Q. Gong, P.-J. Zhao, Y.-J. Jia, Q. Zhao, J.-H. Duan and C.-G. Zou, Nat. Commun., 2023, 14, 240 CrossRef CAS PubMed.
- H. Peixoto, M. Roxo, S. Krstin, T. Roehrig, E. Richling and M. Wink, J. Agric. Food Chem., 2016, 64, 1283–1290 CrossRef CAS PubMed.
- L. Gao, R. Zhang, J. Lan, R. Ning, D. Wu, D. Chen and W. Zhao, J. Nat. Prod., 2016, 79, 3039–3046 CrossRef CAS PubMed.
- T. E. Johnson, Proc. Natl. Acad. Sci. U. S. A., 1987, 84, 3777–3781 CrossRef CAS PubMed.
- Y. Liu, Z. Zhou, L. Yin, M. Zhu, F. Wang, L. Zhang, H. Wang, Z. Zhou, H. Zhu, C. Huang and S. Fan, BioFactors, 2022, 48, 442–453 CrossRef CAS PubMed.
- B. Gerstbrein, G. Stamatas, N. Kollias and M. Driscoll, Aging Cell, 2005, 4, 127–137 CrossRef CAS PubMed.
- Q. Ruan, Y. Qiao, Y. Zhao, Y. Xu, M. Wang, J. Duan and D. Wang, J. Ethnopharmacol., 2016, 177, 101–110 CrossRef CAS PubMed.
- Z. Pincus, T. C. Mazer and F. J. Slack, Aging, 2016, 8, 889–898 CrossRef CAS PubMed.
- G. Di Rosa, G. Brunetti, M. Scuto, A. T. Salinaro, E. J. Calabrese, R. Crea, C. Schmitz-Linneweber, V. Calabrese and N. Saul, Int. J. Mol. Sci., 2020, 21, 3893 CrossRef CAS PubMed.
- S. Li, H. A. Stone and C. T. Murphy, Lab Chip, 2015, 15, 524–531 RSC.
- D. S. Yoon, M.-H. Lee and D. S. Cha, Bio-Protoc., 2018, 8, e2774 Search PubMed.
- P. Back, B. P. Braeckman and F. Matthijssens, Oxid. Med. Cell. Longevity, 2012, 608478 Search PubMed.
- G. J. Lithgow and G. A. Walker, Mech. Ageing Dev., 2002, 123, 765–771 CrossRef PubMed.
- J. R. Cypser, P. Tedesco and T. E. Johnson, Exp. Gerontol., 2006, 41, 935–939 CrossRef CAS PubMed.
- T. E. Johnson, E. de Castro, S. Hegi de Castro, J. Cypser, S. Henderson and P. Tedesco, Exp. Gerontol., 2001, 36, 1609–1617 CrossRef CAS PubMed.
- K. M. Van Pelt and M. C. Truttmann, Transl. Med. Aging, 2020, 4, 60–72 CrossRef PubMed.
- R. Keowkase and N. Weerapreeyakul, Planta Med., 2016, 82, 516–523 CrossRef CAS PubMed.
- W. Cai, J. Wu, X. Wang, J. Huang, Z. Shen and X. Chen, Tradit. Med. Mod. Med., 2019, 2, 19–25 CrossRef CAS.
- S. Govindan, M. Amirthalingam, K. Duraisamy, T. Govindhan, N. Sundararaj and S. Palanisamy, Biomed. Pharmacother., 2018, 102, 812–822 CrossRef CAS PubMed.
- Y. Chen, B. Onken, H. Chen, S. Xiao, X. Liu, M. Driscoll, Y. Cao and Q. Huang, J. Agric. Food Chem., 2014, 62, 3422–3431 CrossRef CAS PubMed.
- A. Upadhyay, J. Chompoo, N. Taira, M. Fukuta and S. Tawata, Biosci., Biotechnol., Biochem., 2013, 77, 217–223 CrossRef CAS PubMed.
- J. Li, D. Liu, D. Li, Y. Guo, H. Du and Y. Cao, Chem. Biodiversity, 2022, 19, e202100685 CAS.
- W. Chen, L. Rezaizadehnajafi and M. Wink, J. Pharm. Pharmacol., 2013, 65, 682–688 CrossRef CAS PubMed.
- M. Lucanic, W. T. Plummer, E. Chen, J. Harke, A. C. Foulger, B. Onken, A. L. Coleman-Hulbert, K. J. Dumas, S. Guo, E. Johnson, D. Bhaumik, J. Xue, A. B. Crist, M. P. Presley, G. Harinath, C. A. Sedore, M. Chamoli, S. Kamat, M. K. Chen, S. Angeli, C. Chang, J. H. Willis, D. Edgar, M. A. Royal, E. A. Chao, S. Patel, T. Garrett, C. Ibanez-Ventoso, J. Hope, J. L. Kish, M. Guo, G. J. Lithgow, M. Driscoll and P. C. Phillips, Nat. Commun., 2017, 8, 14256 CrossRef CAS PubMed.
- J. Lee, G. Kwon, J. Park, J.-K. Kim and Y.-H. Lim, Exp. Biol. Med., 2016, 241, 1757–1763 CrossRef CAS PubMed.
- J. Li, Z. Lin, X. Tang, G. Liu, Y. Chen, X. Zhai, Q. Huang and Y. Cao, Food Funct., 2020, 11, 6595–6607 RSC.
- P. R. Hunt, T. G. Son, M. A. Wilson, Q.-S. Yu, W. H. Wood, Y. Zhang, K. G. Becker, N. H. Greig, M. P. Mattson, S. Camandola and C. A. Wolkow, PLoS One, 2011, 6, e21922 CrossRef CAS PubMed.
- A. S. Arampatzis, O. Tsave, B. Kirchweger, J. Zwirchmayr, V. P. Papageorgiou, J. M. Rollinger and A. N. Assimopoulou, Front. Pharmacol., 2022, 13, 909285 CrossRef CAS PubMed.
- S. Schmeisser, K. Schmeisser, S. Weimer, M. Groth, S. Priebe, E. Fazius, D. Kuhlow, D. Pick, J. W. Einax, R. Guthke, M. Platzer, K. Zarse and M. Ristow, Aging Cell, 2013, 12, 508–517 CrossRef CAS PubMed.
- Q. Zhu, Y. Qu, X.-G. Zhou, J.-N. Chen, H.-R. Luo and G.-S. Wu, Oxid. Med. Cell. Longevity, 2020, 6069354 CAS.
- B. A. Akhoon, S. Pandey, S. Tiwari and R. Pandey, Exp. Gerontol., 2016, 78, 47–56 CrossRef CAS PubMed.
- Y. Qu, L. Shi, Y. Liu, L. Huang, H.-R. Luo and G.-S. Wu, Oxid. Med. Cell. Longevity, 2022, 8878923 CAS.
- K. Pietsch, N. Saul, S. Chakrabarti, S. R. Stürzenbaum, R. Menzel and C. E. W. Steinberg, Biogerontology, 2011, 12, 329–347 CrossRef CAS PubMed.
- J. Nance and C. Frøkjær-Jensen, Genetics, 2019, 212, 959–990 CrossRef CAS PubMed.
- L. R. Girard, T. J. Fiedler, T. W. Harris, F. Carvalho, I. Antoshechkin, M. Han, P. W. Sternberg, L. D. Stein and M. Chalfie, Nucleic Acids Res., 2007, 35, D472–D475 CrossRef CAS PubMed.
- WormAtlas, ed. Z. F. Altun, L. A. Herndon, C. A. Wolkow, C. Crocker, R. Lints and D. H. Hall, pp. 2002–2023, https://www.wormatlas.org Search PubMed.
- Caenorhabditis Genetics Center (CGC), https://cgc.umn.edu, accessed 05.03.2023 Search PubMed.
- Worm Base, https://wormbase.org/#012-34-5, accessed 05.03.2023 Search PubMed.
- O. Thompson, M. Edgley, P. Strasbourger, S. Flibotte, B. Ewing, R. Adair, V. Au, I. Chaudhry, L. Fernando, H. Hutter, A. Kieffer, J. Lau, N. Lee, A. Miller, G. Raymant, B. Shen, J. Shendure, J. Taylor, E. H. Turner, L. W. Hillier, D. G. Moerman and R. H. Waterston, Genome Res., 2013, 23, 1749–1762 CrossRef CAS PubMed.
- X. Zhao, L. Lu, Y. Qi, M. Li and L. Zhou, Biosci., Biotechnol., Biochem., 2017, 81, 1908–1916 CrossRef CAS PubMed.
- K. Filippopoulou, N. Papaevgeniou, M. Lefaki, A. Paraskevopoulou, D. Biedermann, V. Kren and N. Chondrogianni, Free Radical Biol. Med., 2017, 103, 256–267 CrossRef CAS PubMed.
- W.-Y. Zeng, L. Tan, C. Han, Z.-Y. Zheng, G.-S. Wu, H.-R. Luo and S.-L. Li, Oxid. Med. Cell. Longevity, 2021, 7656834 CAS.
- J. H. Kim, I. H. Bang, Y. J. Noh, D. K. Kim, E. J. Bae and I. H. Hwang, Int. J. Mol. Sci., 2020, 21, 2212 CrossRef CAS PubMed.
- C. W. Yu, C. C. Wei and V. H. C. Liao, Free Radical Res., 2014, 48, 371–379 CrossRef CAS PubMed.
- A. Le Bras, Lab. Anim., 2020, 49, 311 CrossRef.
- J. S. Watts, H. F. Harrison, S. Omi, Q. Guenthers, J. Dalelio, N. Pujol and J. L. Watts, G3: Genes, Genomes, Genet., 2020, 10, 4167–4176 CrossRef CAS PubMed.
- P. Shah, Z. Bao and R. Zaidel-Bar, Genetics, 2022, 221, iyac068 CrossRef PubMed.
- Y. Wang, S. Luo, Z. Xu, L. Liu, S. Feng, T. Chen, L. Zhou, M. Yuan, Y. Huang and C. b. Ding, Arabian J. Chem., 2021, 14, 103149 CrossRef CAS.
- Y. Gong, Y. Luo, J.-A. Huang, J. Zhang, Y. Peng, Z. Liu and B. Zhao, J. Funct. Foods, 2012, 4, 988–993 CrossRef CAS.
- J. H. An and T. K. Blackwell, Genes Dev., 2003, 17, 1882–1893 CrossRef CAS PubMed.
- J. M. A. Tullet, M. Hertweck, J. H. An, J. Baker, J. Y. Hwang, S. Liu, R. P. Oliveira, R. Baumeister and T. K. Blackwell, Cell, 2008, 132, 1025–1038 CrossRef CAS PubMed.
- S. Havermann, R. Rohrig, Y. Chovolou, H. U. Humpf and W. Wätjen, J. Agric. Food Chem., 2013, 61, 2158–2164 CrossRef CAS PubMed.
- J. Nass, S. Abdelfatah and T. Efferth, Phytomedicine, 2021, 84, 153482 CrossRef CAS PubMed.
- Y. Niu, J. Liao, H. Zhou, C.-C. Wang, L. Wang and Y. Fan, Molecules, 2022, 27, 4952 CrossRef CAS PubMed.
- J. Liu, J. Meng, J. Du, X. Liu, Q. Pu, D. Di and C. Chen, Molecules, 2020, 25, 3511 CrossRef CAS PubMed.
- K. Koch, N. Weldle, S. Baier, C. Büchter and W. Wätjen, Eur. J. Nutr., 2020, 59, 137–150 CrossRef CAS PubMed.
- X. Wang, X. Li, L. Li, X. Yang, J. Wang, X. Liu, J. Chen, S. Liu, N. Zhang, J. Li and H. Wang, Food Funct., 2022, 13, 10680–10694 RSC.
- F. Yan, Y. Yang, L. Yu and X. Zheng, J. Agric. Food Chem., 2017, 65, 7457–7466 CrossRef CAS PubMed.
- H. Wang, J. Liu, T. Li and R. H. Liu, Food Funct., 2018, 9, 5273–5282 RSC.
- C. Duangjan, P. Rangsinth, X. Gu, M. Wink and T. Tencomnao, Oxid. Med. Cell. Longevity, 2019, 9012396 CAS.
- S. Pandey, S. Tiwari, A. Kumar, A. Niranjan, J. Chand, A. Lehri and P. S. Chauhan, Ind. Crops Prod., 2018, 120, 113–122 CrossRef CAS.
- A. L. Hsu, C. T. Murphy and C. Kenyon, Science, 2003, 300, 1142–1145 CrossRef CAS PubMed.
- J. F. Morley and R. I. Morimoto, Mol. Biol. Cell, 2004, 15, 657–664 CrossRef CAS PubMed.
- J. Brunquell, S. Morris, A. Snyder and S. D. Westerheide, Cell Stress Chaperones, 2018, 23, 65–75 CrossRef CAS PubMed.
- W. Liu, Y. Guan, S. Qiao, J. Wang, K. Bao, Z. Mao, L. Liao, A. Moskalev, B. Jiang, J. Zhu, C. Xia, J. Li and Z. Hu, Oxid. Med. Cell. Longevity, 2021, 9942090 CAS.
- Y.-P. Zhang, W.-H. Zhang, P. Zhang, Q. Li, Y. Sun, J.-W. Wang, S. O. Zhang, T. Cai, C. Zhan and M.-Q. Dong, Nat. Commun., 2022, 13, 6339 CrossRef CAS PubMed.
- B. Song, B. Zheng, T. Li and R. H. Liu, Food Funct., 2020, 11, 3598–3609 RSC.
- H. Jeon and D. S. Cha, Chin. J. Nat. Med., 2016, 14, 335–342 CAS.
- F. Yan, X.-A. Chen and X. Zheng, Food Res. Int., 2017, 102, 213–224 CrossRef CAS PubMed.
- S. Ogg, S. Paradis, S. Gottlieb, G. I. Patterson, L. Lee, H. A. Tissenbaum and G. Ruvkun, Nature, 1997, 389, 994–999 CrossRef CAS PubMed.
- K. Lin, J. B. Dorman, A. Rodan and C. Kenyon, Science, 1997, 278, 1319–1322 CrossRef CAS PubMed.
- V. Shukla, D. Yadav, S. C. Phulara, M. M. Gupta, S. K. Saikia and R. Pandey, Free Radical Biol. Med., 2012, 53, 1848–1856 CrossRef CAS PubMed.
- J. M. Romero-Marquez, M. D. Navarro-Hortal, V. Jimenez-Trigo, P. Munoz-Ollero, T. Y. Forbes-Hernandez, A. Esteban-Munoz, F. Giampieri, I. Delgado Noya, P. Bullon, L. Vera-Ramirez, M. Battino, C. Sanchez-Gonzalez and J. L. Quiles, Antioxidants, 2022, 11, 629 CrossRef CAS PubMed.
- M. Roxo, H. Peixoto, P. Wetterauer, E. Lima and M. Wink, Oxid. Med. Cell. Longevity, 2020, 7590707 CAS.
- C. Duangjan, P. Rangsinth, X. Gu, S. Zhang, M. Wink and T. Tencomnao, Phytomedicine, 2019, 64, 153061 CrossRef CAS PubMed.
- W. B. Bakrim, A. D. R. Nurcahyanti, M. Dmirieh, I. Mahdi, A. M. Elgamal, M. A. El Raey, M. Wink and M. Sobeh, Oxid. Med. Cell. Longevity, 2022, 3486257 CAS.
- N. M. Hegazi, M. Sobeh, S. Rezq, M. A. El-Raey, M. Dmirieh, A. M. El-Shazly, M. F. Mahmoud and M. Wink, Sci. Rep., 2019, 9, 1–12 CrossRef CAS PubMed.
- M. A. O. Abdelfattah, M. Dmirieh, W. Ben Bakrim, O. Mouhtady, M. A. Ghareeb, M. Wink and M. Sobeh, J. Ethnopharmacol., 2022, 292, 115187 CrossRef CAS PubMed.
- S. Thabit, H. Handoussa, M. Roxo, B. Cestari de Azevedo, N. S. E. El Sayed and M. Wink, Molecules, 2019, 24, 2633 CrossRef CAS PubMed.
- A. L. Tambara, L. de Los Santos Moraes, A. H. Dal Forno, J. R. Boldori, A. T. Goncalves Soares, C. de Freitas Rodrigues, L. R. B. Mariutti, A. Z. Mercadante, D. S. de Avila and C. C. Denardin, Food Chem. Toxicol., 2018, 120, 639–650 CrossRef CAS PubMed.
- J. Zheng, D. Heber, M. Wang, C. Gao, S. B. Heymsfield, R. J. Martin, F. L. Greenway, J. W. Finley, J. H. Burton, W. D. Johnson, F. M. Enright, M. J. Keenan and Z. Li, Int. J. Vitam. Nutr. Res., 2017, 87, 149–158 CrossRef CAS PubMed.
- M. S. Bonkowski and D. A. Sinclair, Nat. Rev. Mol. Cell Biol., 2016, 17, 679–690 CrossRef CAS PubMed.
- H. A. Tissenbaum and L. Guarente, Nature, 2001, 410, 227–230 CrossRef CAS PubMed.
- M. Maulik, S. Mitra, S. Hunter, M. Hunstiger, S. R. Oliver, A. Bult-Ito and B. E. Taylor, Sci. Rep., 2018, 8, 1–13 CAS.
- L.-G. Xiong, J.-A. Huang, J. Li, P.-H. Yu, Z. Xiong, J.-W. Zhang, Y.-S. Gong, Z.-H. Liu and J.-H. Chen, J. Agric. Food Chem., 2014, 62, 11163–11169 CrossRef CAS PubMed.
- D. J. Deusing, S. Winter, A. Kler, E. Kriesl, B. Bonnlaender, U. Wenzel and E. Fitzenberger, Fitoterapia, 2015, 102, 163–170 CrossRef CAS PubMed.
- S. Robida-Stubbs, K. Glover-Cutter, D. W. Lamming, M. Mizunuma, S. D. Narasimhan, E. Neumann-Haefelin, D. M. Sabatini and T. K. Blackwell, Cell Metab., 2012, 15, 713–724 CrossRef CAS PubMed.
- V. T. Cuong, W. Chen, J. Shi, M. Zhang, H. Yang, N. Wang, S. Yang, J. Li, P. Yang and J. Fei, Exp. Gerontol., 2019, 117, 99–105 CrossRef CAS PubMed.
- J. Apfeld, G. O'Connor, T. McDonagh, P. S. DiStefano and R. Curtis, Genes Dev., 2004, 18, 3004–3009 CrossRef CAS PubMed.
- B. Onken and M. Driscoll, PLoS One, 2010, 5, e8758 CrossRef PubMed.
- H. J. Weir, P. Yao, F. K. Huynh, C. C. Escoubas, R. L. Goncalves, K. Burkewitz, R. Laboy, M. D. Hirschey and W. B. Mair, Cell Metab., 2017, 26, 884–896 CrossRef CAS PubMed.
- L. Chen, H. Cao, Q. Huang, J. Xiao and H. Teng, Crit. Rev. Food Sci. Nutr., 2022, 62, 7730–7742 CrossRef CAS PubMed.
- B. Spanier, R. Lang, D. Weber, A. Lechner, T. Thoma, M. Rothner, K. Petzold, T. Lang, A. Beusch, M. Bösl, V. Schlagbauer, H. Daniel and T. Hofmann, J. Agric. Food Chem., 2019, 67, 4774–4781 CrossRef CAS PubMed.
- M. Dueñas, F. Surco-Laos, S. González-Manzano, A. M. González-Paramás, E. Gómez-Orte, J. Cabello and C. Santos-Buelga, Pharmacol. Res., 2013, 76, 41–48 CrossRef PubMed.
- F. A. Tomás-Barberán, R. García-Villalba, A. González-Sarrías, M. V. Selma and J. C. Espín, J. Agric. Food Chem., 2014, 62, 6535–6538 CrossRef PubMed.
- EFSA, Novel Food Authorisation, https://open.efsa.europa.eu/question/EFSA-Q-2018-00724_2023, accessed 03.08.2023 Search PubMed.
- P. A. Andreux, W. Blanco-Bose, D. Ryu, F. Burdet, M. Ibberson, P. Aebischer, J. Auwerx, A. Singh and C. Rinsch, Nat. Metab., 2019, 1, 595–603 CrossRef CAS PubMed.
- J. G. Wood, B. Rogina, S. Lavu, K. Howitz, S. L. Helfand, M. Tatar and D. Sinclair, Nature, 2004, 430, 686–689 CrossRef CAS PubMed.
- S. Feng, C. Zhang, T. Chen, L. Zhou, Y. Huang, M. Yuan, T. Li and C. Ding, Antioxidants, 2021, 10, 1697 CrossRef CAS PubMed.
- A. Matencio, M. A. Guerrero-Rubio, F. Caldera, C. Cecone, F. Trotta, F. Garcia-Carmona and J. M. Lopez-Nicolas, Int. J. Pharm., 2020, 589, 119862 CrossRef CAS PubMed.
- T. M. Bass, D. Weinkove, K. Houthoofd, D. Gems and L. Partridge, Mech. Ageing Dev., 2007, 128, 546–552 CrossRef CAS PubMed.
- R. Strong, R. A. Miller, C. M. Astle, J. A. Baur, R. de Cabo, E. Fernandez, W. Guo, M. Javors, J. L. Kirkland, J. F. Nelson, D. A. Sinclair, B. Teter, D. Williams, N. Zaveri, N. L. Nadon and D. E. Harrison, J. Gerontol., Ser. A, 2013, 68, 6–16 CrossRef CAS PubMed.
- J. A. Baur, K. J. Pearson, N. L. Price, H. A. Jamieson, C. Lerin, A. Kalra, V. V. Prabhu, J. S. Allard, G. Lopez-Lluch, K. Lewis, P. J. Pistell, S. Poosala, K. G. Becker, O. Boss, D. Gwinn, M. Wang, S. Ramaswamy, K. W. Fishbein, R. G. Spencer, E. G. Lakatta, D. Le Couteur, R. J. Shaw, P. Navas, P. Puigserver, D. K. Ingram, R. de Cabo and D. A. Sinclair, Nature, 2006, 444, 337–342 CrossRef CAS PubMed.
- K. J. Pearson, J. A. Baur, K. N. Lewis, L. Peshkin, N. L. Price, N. Labinskyy, W. R. Swindell, D. Kamara, R. K. Minor, E. Perez, H. A. Jamieson, Y. Zhang, S. R. Dunn, K. Sharma, N. Pleshko, L. A. Woollett, A. Csiszar, Y. Ikeno, D. Le Couteur, P. J. Elliott, K. G. Becker, P. Navas, D. K. Ingram, N. S. Wolf, Z. Ungvari, D. A. Sinclair and R. de Cabo, Cell Metab., 2008, 8, 157–168 CrossRef CAS PubMed.
- D. Chen and L. Guarente, Trends Mol. Med., 2007, 13, 64–71 CrossRef CAS PubMed.
- N. L. Price, A. P. Gomes, A. J. Ling, F. V. Duarte, A. Martin-Montalvo, B. J. North, B. Agarwal, L. Ye, G. Ramadori, J. S. Teodoro, B. P. Hubbard, A. T. Varela, J. G. Davis, B. Varamini, A. Hafner, R. Moaddel, A. P. Rolo, R. Coppari, C. M. Palmeira, R. de Cabo, J. A. Baur and D. A. Sinclair, Cell Metab., 2012, 15, 675–690 CrossRef CAS PubMed.
- B. P. Hubbard, A. P. Gomes, H. Dai, J. Li, A. W. Case, T. Considine, T. V. Riera, J. E. Lee, S. Y. E, D. W. Lamming, B. L. Pentelute, E. R. Schuman, L. A. Stevens, A. J. Y. Ling, S. M. Armour, S. Michan, H. Zhao, Y. Jiang, S. M. Sweitzer, C. A. Blum, J. S. Disch, P. Y. Ng, K. T. Howitz, A. P. Rolo, Y. Hamuro, J. Moss, R. B. Perni, J. L. Ellis, G. P. Vlasuk and D. A. Sinclair, Science, 2013, 339, 1216–1219 CrossRef CAS PubMed.
- K. Pallauf, I. Günther, G. Kühn, D. Chin, S. de Pascual-Teresa and G. Rimbach, Adv. Nutr., 2021, 12, 995–1005 CrossRef PubMed.
- C. Burnett, S. Valentini, F. Cabreiro, M. Goss, M. Somogyvári, M. D. Piper, M. Hoddinott, G. L. Sutphin, V. Leko, J. J. McElwee, R. P. Vazquez-Manrique, A. M. Orfila, D. Ackerman, C. Au, G. Vinti, M. Riesen, K. Howard, C. Neri, A. Bedalov, M. Kaeberlein, C. Soti, L. Partridge and D. Gems, Nature, 2011, 477, 482–485 CrossRef CAS PubMed.
- M. Kaeberlein, T. McDonagh, B. Heltweg, J. Hixon, E. A. Westman, S. D. Caldwell, A. Napper, R. Curtis, P. S. DiStefano, S. Fields, A. Bedalov and B. K. Kennedy, J. Biol. Chem., 2005, 280, 17038–17045 CrossRef CAS PubMed.
- C. Brenner, Life Metab., 2022, 1, 122–133 CrossRef PubMed.
- S. T. Soukup, B. Spanier, G. Grünz, D. Bunzel, H. Daniel and S. E. Kulling, PLoS One, 2012, 7, e46914 CrossRef CAS PubMed.
- S. Ehala, M. Vaher and M. Kaljurand, J. Agric. Food Chem., 2005, 53, 6484–6490 CrossRef CAS PubMed.
- S. Weiskirchen and R. Weiskirchen, Adv. Nutr., 2016, 7, 706–718 CrossRef CAS PubMed.
- H. Yuan and R. Marmorstein, Science, 2013, 339, 1156–1157 CrossRef CAS PubMed.
- T. Nunes, L. Almeida, J.-F. Rocha, A. Falcão, C. Fernandes-Lopes, A. I. Loureiro, L. Wright, M. Vaz-da-Silva and P. Soares-da-Silva, J. Clin. Pharmacol., 2009, 49, 1477–1482 CrossRef CAS PubMed.
- R. Zamora-Ros, N. G. Forouhi, S. J. Sharp, C. A. González, B. Buijsse, M. Guevara, Y. T. van der Schouw, P. Amiano, H. Boeing, L. Bredsdorff, G. Fagherazzi, E. J. Feskens, P. W. Franks, S. Grioni, V. Katzke, T. J. Key, K. T. Khaw, T. Kühn, G. Masala, A. Mattiello, E. Molina-Montes, P. M. Nilsson, K. Overvad, F. Perquier, M. L. Redondo, F. Ricceri, O. Rolandsson, I. Romieu, N. Roswall, A. Scalbert, M. Schulze, N. Slimani, A. M. Spijkerman, A. Tjonneland, M. J. Tormo, M. Touillaud, R. Tumino, A. D. van der, G. J. van Woudenbergh, C. Langenberg, E. Riboli and N. J. Wareham, J. Nutr., 2014, 144, 335–343 CrossRef CAS PubMed.
- M. G. Hertog, E. J. Feskens, D. Kromhout, P. Hollman and M. Katan, Lancet, 1993, 342, 1007–1011 CrossRef CAS PubMed.
- M. G. Hertog, D. Kromhout, C. Aravanis, H. Blackburn, R. Buzina, F. Fidanza, S. Giampaoli, A. Jansen, A. Menotti and S. Nedeljkovic, Arch. Intern. Med., 1995, 155, 381–386 CrossRef CAS PubMed.
- T. M. Holland, P. Agarwal, Y. Wang, K. Dhana, S. E. Leurgans, K. Shea, S. L. Booth, K. B. Rajan, J. A. Schneider and L. L. Barnes, Neurology, 2023, 100, e694 CrossRef CAS PubMed.
- A. Kampkötter, C. Timpel, R. F. Zurawski, S. Ruhl, Y. Chovolou, P. Proksch and W. Wätjen, Comp. Biochem. Physiol., Part B, 2008, 149, 314–323 CrossRef PubMed.
- S. Havermann, H.-U. Humpf and W. Waetjen, Fitoterapia, 2016, 113, 123–127 CrossRef CAS PubMed.
- B. Ayuda-Durán, E. Sánchez-Hernández, S. González-Manzano, C. Santos-Buelga and A. M. González-Paramás, Front. Nutr., 2022, 9, 989427 CrossRef PubMed.
- G. Grünz, K. Haas, S. Soukup, M. Klingenspor, S. E. Kulling, H. Daniel and B. Spanier, Mech. Ageing Dev., 2012, 133, 1–10 CrossRef PubMed.
- F. Surco-Laos, J. Cabello, E. Gómez-Orte, S. González-Manzano, A. M. González-Paramás, C. Santos-Buelga and M. Dueñas, Food Funct., 2011, 2, 445–456 RSC.
- M. F. M. Sciacca, V. Romanucci, A. Zarrelli, I. Monaco, F. Lolicato, N. Spinella, C. Galati, G. Grasso, L. D'Urso, M. Romeo, L. Diomede, M. Salmona, C. Bongiorno, G. Di Fabio, C. La Rosa and D. Milardi, ACS Chem. Neurosci., 2017, 8, 1767–1778 CrossRef CAS PubMed.
- G. Buckland and C. A. Gonzalez, Br. J. Nutr., 2015, 113, S94–S101 CrossRef CAS PubMed.
- R. Estruch, E. Ros, J. Salas-Salvadó, M.-I. Covas, D. Corella, F. Arós, E. Gómez-Gracia, V. Ruiz-Gutiérrez, M. Fiol and J. Lapetra, N. Engl. J. Med., 2018, 378, e34 CrossRef CAS PubMed.
- M. A. Martinez-Gonzalez, M. Bes-Rastrollo, L. Serra-Majem, D. Lairon, R. Estruch and A. Trichopoulou, Nutr. Rev., 2009, 67, S111–S116 CrossRef PubMed.
- L. G. Gillingham, S. Harris-Janz and P. J. Jones, Lipids, 2011, 46, 209–228 CrossRef CAS PubMed.
- S. Han, E. A. Schroeder, C. G. Silva-García, K. Hebestreit, W. B. Mair and A. Brunet, Nature, 2017, 544, 185–190 CrossRef CAS PubMed.
- J. Goudeau, S. Bellemin, E. Toselli-Mollereau, M. Shamalnasab, Y. Chen and H. Aguilaniu, PLoS Biol., 2011, 9, e1000599 CrossRef CAS PubMed.
- S.-J. Lee, A. B. Hwang and C. Kenyon, Curr. Biol., 2010, 20, 2131–2136 CrossRef CAS PubMed.
- R. Ratnappan, F. R. G. Amrit, S.-W. Chen, H. Gill, K. Holden, J. Ward, K. R. Yamamoto, C. P. Olsen and A. Ghazi, PLoS Genet., 2014, 10, e1004829 CrossRef PubMed.
- J. C. Garcia-Moreno, M. Porta de la Riva, E. Martínez-Lara, E. Siles and A. Cañuelo, Neurobiol. Aging, 2019, 82, 60–68 CrossRef CAS PubMed.
- A. Cañuelo, B. Gilbert-López, P. Pacheco-Liñán, E. Martínez-Lara, E. Siles and A. Miranda-Vizuete, Mech. Ageing Dev., 2012, 133, 563–574 CrossRef PubMed.
- G. Di Rosa, N. Saul, C. Schmitz-Linneweber and V. Calabrese, Pathophysiology, 2018, 25, 203 CrossRef.
- L. Diomede, S. Rigacci, M. Romeo, M. Stefani and M. Salmona, PLoS One, 2013, 8, e58893 CrossRef CAS PubMed.
- M. Katsiki, N. Chondrogianni, I. Chinou, A. J. Rivett and E. S. Gonos, Rejuvenation Res., 2007, 10, 157–172 CrossRef CAS PubMed.
- T. Nikou, V. Liaki, P. Stathopoulos, A. D. Sklirou, E. N. Tsakiri, T. Jakschitz, G. Bonn, I. P. Trougakos, M. Halabalaki and L. A. Skaltsounis, Food Chem. Toxicol., 2019, 125, 403–412 CrossRef CAS PubMed.
- M. A. Guerrero-Rubio, S. Hernandez-Garcia, F. Garcia-Carmona and F. Gandia-Herrero, Food Chem., 2019, 274, 840–847 CrossRef CAS PubMed.
- M. H. Land, M. L. Toth, L. MacNair, S. A. Vanapalli, T. W. Lefever, E. N. Peters and M. O. Bonn-Miller, Cannabis Cannabinoid Res., 2020, 6, 522–527 CrossRef PubMed.
- Z. Wang, P. Zheng, Y. Xie, X. Chen, N. Solowij, K. Green, Y. L. Chew and X. F. Huang, FASEB J., 2021, 35, e21537 CAS.
- A. P. Vanin, W. A. Tamagno, C. Alves, L. Mesacasa, L. F. Santin, N. T. Sutorillo, D. Bilibio, C. Müller, L. Galon and R. R. Kaizer, Sci. Rep., 2022, 12, 15376 CrossRef CAS PubMed.
- Z. Wang, P. Zheng, X. Chen, Y. Xie, K. Weston-Green, N. Solowij, Y. L. Chew and X.-F. Huang, Geroscience, 2022, 44, 1505–1524 CrossRef CAS PubMed.
- Z. Pandelides, C. Thornton, A. S. Faruque, A. P. Whitehead, K. L. Willett and N. M. Ashpole, Geroscience, 2020, 42, 785–800 CrossRef CAS PubMed.
- D. O. Mitchell, L. Wen Jing, C. Tobias, A. B. Bruce and K. Richard, J. Neurosci., 2017, 37, 2859 CrossRef PubMed.
- A. Levichev, S. Faumont, R. Z. Berner, Z. Purcell, A. M. White, K. Chicas-Cruz and S. R. Lockery, Curr. Biol., 2023, 33, 1625–1639 CrossRef CAS PubMed.
- S. I. Pastuhov, K. Matsumoto and N. Hisamoto, Genes Cells, 2016, 21, 696–705 CrossRef CAS PubMed.
- N. Harrison, M. A. Lone, T. K. Kaul, P. Reis Rodrigues, I. V. Ogungbe and M. S. Gill, PLoS One, 2014, 9, e113007 CrossRef PubMed.
- M. Lucanic, J. M. Held, M. C. Vantipalli, I. M. Klang, J. B. Graham, B. W. Gibson, G. J. Lithgow and M. S. Gill, Nature, 2011, 473, 226–229 CrossRef CAS PubMed.
- S.-C. Cheng, W.-H. Li, Y.-C. Shi, P.-L. Yen, H.-Y. Lin, V. H.-C. Liao and S.-T. Chang, J. Agric. Food Chem., 2014, 62, 4159–4165 CrossRef CAS PubMed.
- A. Takahashi, T. Watanabe, T. Fujita, T. Hasegawa, M. Saito and M. Suganuma, Biosci., Biotechnol., Biochem., 2014, 78, 1206–1211 CrossRef CAS PubMed.
- P. Yuan, L.-Y. Pan, L.-G. Xiong, J.-W. Tong, J. Li, J.-A. Huang, Y.-S. Gong and Z.-H. Liu, Food Funct., 2018, 9, 3798–3806 RSC.
- S. Abbas and M. Wink, Antioxidants, 2014, 3, 129 CrossRef PubMed.
- S. Zhang, C. Duangjan, T. Tencomnao, J. Liu, J. Lin and M. Wink, Food Funct., 2020, 11, 8179–8192 RSC.
- C.-C. Wei, C.-W. Yu, P.-L. Yen, H.-Y. Lin, S.-T. Chang, F.-L. Hsu and V. H.-C. Liao, J. Agric. Food Chem., 2014, 62, 10701–10707 CrossRef CAS PubMed.
- G. Dominguez-Rodriguez, D. Ramon Vidal, P. Martorell, M. Plaza and M. L. Marina, J. Agric. Food Chem., 2022, 70, 7993–8009 CrossRef CAS PubMed.
- J. S. Sangha, D. Fan, A. H. Banskota, R. Stefanova, W. Khan, J. Hafting, J. Craigie, A. T. Critchley and B. Prithiviraj, J. Funct. Foods, 2013, 5, 1180–1190 CrossRef CAS.
- N. Shen, W. Zeng, F. Leng, J. Lu, Z. Lu, J. Cui, L. Wang and B. Jin, Food Funct., 2021, 12, 12395–12406 RSC.
- L. Xiong, N. Deng, B. Zheng, T. Li and R. H. Liu, Food Funct., 2021, 12, 7851–7866 RSC.
- S. Guha, O. Nataraja, C. G. Murbach, J. Dinh, E. C. Wilson, M. Cao, S. Zou and Y. Dong, Nutrients, 2014, 6, 911–921 CrossRef CAS PubMed.
- S. Guha, M. Cao, R. M. Kane, A. M. Savino, S. Zou and Y. Dong, Age (Dordr), 2013, 35, 1559–1574 CrossRef PubMed.
- C. Scerbak, E. Vayndorf, A. Hernandez, C. McGill and B. Taylor, GeroScience, 2018, 40, 151–162 CrossRef CAS PubMed.
- M. Sobeh, A. Esmat, G. Petruk, M. A. O. Abdelfattah, M. Dmirieh, D. M. Monti, A. B. Abdel-Naim and M. Wink, J. Funct. Foods, 2018, 41, 223–231 CrossRef CAS.
- M. I. Prasanth, J. M. Brimson, S. Chuchawankul, M. Sukprasansap and T. Tencomnao, Oxid. Med. Cell. Longevity, 2019, 7024785 CAS.
- A.-G. Wu, J.-F. Teng, V. K.-W. Wong, X.-G. Zhou, W.-Q. Qiu, Y. Tang, J.-M. Wu, R. Xiong, R. Pan, Y.-L. Wang, B. Tang, T.-Y. Ding, L. Yu, W. Zeng, D.-L. Qin and B. Y.-K. Law, Phytomedicine, 2019, 65, 153088 CrossRef CAS PubMed.
- B. Tsolmon, Y. Fang, T. Yang, L. Guo, K. He, G.-Y. Li and H. Zhao, Food Chem., 2021, 343, 128392 CrossRef CAS PubMed.
- W. Jia, Q. Peng, L. Su, X. Yu, C. W. Ma, M. Liang, X. Yin, Y. Zou and Z. Huang, Mar. Drugs, 2018, 16, 444 CrossRef CAS PubMed.
- E. B. Lee, M. M. Xing and D. K. Kim, Arch. Pharmacal Res., 2017, 40, 825–835 CrossRef CAS PubMed.
- F. Zhang, M. Berg, K. Dierking, M.-A. Félix, M. Shapira, B. S. Samuel and H. Schulenburg, Front. Microbiol., 2017, 8, 485 Search PubMed.
- C. Heintz and W. Mair, Cell, 2014, 156, 408–411 CrossRef CAS PubMed.
- I. Gusarov, L. Gautier, O. Smolentseva, I. Shamovsky, S. Eremina, A. Mironov and E. Nudler, Cell, 2013, 152, 818–830 CrossRef CAS PubMed.
- Q. Sun, N. M. Vega, B. Cervantes, C. P. Mancuso, N. Mao, M. N. Taylor, J. J. Collins, A. S. Khalil, J. Gore and T. K. Lu, Mol. Syst. Biol., 2022, 18, e9933 CrossRef CAS PubMed.
- S. Urquiza-Zurich, V. A. Garcia-Angulo, P. Burdisso, M. F. Palominos, L. Fernandez-Hubeid, P. A. Harcha, J. P. Castillo and A. Calixto, mBio, 2023, 14, e0340222 CrossRef PubMed.
- R. Saito, N. Sato, Y. Okino, D. S. Wang and G. Seo, Biosci. Microbiota, Food Health, 2023, 42, 124–130 CrossRef PubMed.
- A. L. Gaeta, K. Willicott, C. W. Willicott, L. E. McKay, C. M. Keogh, T. J. Altman, L. C. Kimble, A. L. Yarbrough, K. A. Caldwell and G. A. Caldwell, iScience, 2023, 26, 106859 CrossRef CAS PubMed.
- V. Donato, F. R. Ayala, S. Cogliati, C. Bauman, J. G. Costa, C. Leñini and R. Grau, Nat. Commun., 2017, 8, 14332 CrossRef CAS PubMed.
- M. C. Rodríguez-Daza, E. C. Pulido-Mateos, J. Lupien-Meilleur, D. Guyonnet, Y. Desjardins and D. Roy, Front. Nutr., 2021, 8, 689456 CrossRef PubMed.
- E.-M. Pferschy-Wenzig, M. R. Pausan, K. Ardjomand-Woelkart, S. Röck, R. M. Ammar, O. Kelber, C. Moissl-Eichinger and R. Bauer, Nutrients, 2022, 14, 2111 CrossRef CAS PubMed.
- M. Shapira, Curr. Opin. Microbiol., 2017, 38, 142–147 CrossRef CAS PubMed.
- P. Dirksen, A. Assié, J. Zimmermann, F. Zhang, A. M. Tietje, S. A. Marsh, M. A. Félix, M. Shapira, C. Kaleta, H. Schulenburg and B. S. Samuel, G3: Genes, Genomes, Genet., 2020, 10, 3025–3039 CrossRef CAS PubMed.
- R. Van Assche, V. Broeckx, K. Boonen, E. Maes, W. De Haes, L. Schoofs and L. Temmerman, J. Mol. Biol., 2015, 427, 3441–3451 CrossRef CAS PubMed.
- L. K. Caesar and N. B. Cech, Nat. Prod. Rep., 2019, 36, 869–888 RSC.
- A. Panossian, E.-J. Seo and T. Efferth, Phytomedicine, 2018, 50, 257–284 CrossRef PubMed.
- Y. Fu and W. Zhao, J. Nat. Prod., 2020, 83, 505–515 CrossRef CAS PubMed.
- O. Potterat and M. Hamburger, Planta Med., 2014, 80, 1171–1181 CrossRef CAS PubMed.
- A. E. Fox Ramos, L. Evanno, E. Poupon, P. Champy and M. A. Beniddir, Nat. Prod. Rep., 2019, 36, 960–980 RSC.
- U. Grienke, P. A. Foster, J. Zwirchmayr, A. Tahir, J. M. Rollinger and E. Mikros, Sci. Rep., 2019, 9, 11113 CrossRef PubMed.
- J. Langeder, K. Doring, H. Schmietendorf, U. Grienke, M. Schmidtke and J. M. Rollinger, J. Nat. Prod., 2022, 86, 8–17 CrossRef PubMed.
- J. J. Kellogg, D. A. Todd, J. M. Egan, H. A. Raja, N. H. Oberlies, O. M. Kvalheim and N. B. Cech, J. Nat. Prod., 2016, 79, 376–386 CrossRef CAS PubMed.
- J. Zwirchmayr, C. D. Cruz, U. Grienke, P. Tammela and J. M. Rollinger, iScience, 2023, 107523 CrossRef.
- M. Heinrich, B. Jalil, M. Abdel-Tawab, J. Echeverria, Ž. Kulić, L. J. McGaw, J. M. Pezzuto, O. Potterat and J.-B. Wang, Front. Pharmacol., 2022, 13, 953205 CrossRef PubMed.
- J. Bisson, J. B. McAlpine, J. B. Friesen, S. N. Chen, J. Graham and G. F. Pauli, J. Med. Chem., 2016, 59, 1671–1690 CrossRef CAS PubMed.
- J. B. Baell, J. Nat. Prod., 2016, 79, 616–628 CrossRef CAS PubMed.
- T. T. Cushnie and A. J. Lamb, Int. J. Antimicrob. Agents, 2011, 38, 99–107 CrossRef CAS PubMed.
- K.-T. Chung, Z. Lu and M. Chou, Food Chem. Toxicol., 1998, 36, 1053–1060 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: https://doi.org/10.1039/d3np00021d |
| This journal is © The Royal Society of Chemistry 2023 |