
Biocompatibility assessment of type-II collagen and its polypeptide for tissue engineering: effect of collagen's molecular weight and glycoprotein content on tumor necrosis factor (Fas/Apo-1) receptor activation in human acute T-lymphocyte leukemia cell line
E. Jeevithana,
Z. Jingyia,
B. Baoa,
W. Shujunb,
R. JeyaShakila*c and
W. H. Wu*ab
aDepartment of Marine Pharmacology, College of Food Science and Technology, Shanghai Ocean University, Shanghai 201306, China. E-mail: whwu@shou.edu.cn; srijeevithan@gmail.com
bCo-Innovation Center of Jiangsu Marine Bio-industry Technology, Huaihai Institute of Technology, Lianyungang, 222005, China
cDepartment of Fish Quality Assurance and Management, Fish Quality Monitoring and Certification Centre, Fisheries College and Research Institute, Tamil Nadu Fisheries University, Tuticorin 628 008, India. E-mail: jeyashakila@gmail.com
First published on 3rd February 2016
Abstract
Low molecular weight (LMW) (57, 40 and 25 kDa) collagens were separated and Fas cell surface death receptor activation were investigated based on its MW and glycoprotein content. The biochemical properties such as amino acid profile, FTIR and glycoprotein content of LMW collagens were slightly different and confirmed that limited digestion by proteolytic enzyme disrupted the triple helical structure of collagen. Cell adhesion pattern between collagens and immune cells were dose dependent manner. Better active cellular suppression of fish collagen than mammalian collagen was confirmed by increased secretion of immunological indexes (FAS/APO-1, cytokines (IL-2, IL-6) and caspases (casp-3, casp-8)) in T cell. Apoptotic regulatory gene expression (FAS, FAS-L, CASp-3, CASP-8, IL-2 and IL-6) of 6T-CEM cells was further confirmed the biocompatibility of fish collagens. Fluorescence microstructure revealed destruction of immune cells morphology and nuclei due to apoptosis by fish collagens. Consequently, this study disclosed that the apoptosis induction in T cells was based on the MW of collagens and also attachment of glycoprotein with tumor necrosis factor receptor (TNFRSF6) in cell membrane, which further activates DD, FAD, DED and caspase cascade pathway. Accordingly, this study created new platform for the better use of fish collagen in biomedical industries for the treatment of autoimmune diseases.
1. Introduction
For a decade, fish collagen has been extensively studied for biomedical aspects due to its peculiar properties such as formation of tissues, organs and in various functional expressions of cells in order to replace the mammalian collagen because of a possibility of animal diseases transmission to humans.1,2 The structure and function of collagens are varied based on its existence in mammals' body. Up to now, there are 25 types of collagen identified in vertebrates, mainly found in fibrous tissues such as tendons, ligaments and skin, and also abundant in corneas, cartilage, blood vessels, gut and intervertebral discs. In general, collagen present in skin, tendon, bone, cornea, fibrocartilage, large vessels, intestine, uterus and dermis is characterized as type I collagen consisting α1, α2, β and γ chains, whereas collagen present in hyaline cartilages, vitreous, nucleus pulposus and notochord is type-II collagen with three identical α chains. The fish collagen isolated from several marine species was characterized by many researchers.3–56T-CEM cell lines are an immortalized human acute T-lymphocyte leukemia cells. In tissue engineering, it is mainly used to investigate the signalling pathway, genetic expression and receptor mechanism of immune cells against foreign materials. These cell lines are generally isolated from virulent tumours such as acute T-cell lymphoblastic leukemia and T-cell lymphoma, which mainly infect blood cells, bone marrow, and tissues. Due to the potential properties of type-II collagen, many researchers have recently focused to investigate about its oral tolerance effect to treat autoimmune diseases.3,6 Oral tolerance is simply defined as inability of antibody production by immune systems against external materials that enter the body through digestive track. Glycosylated polypeptide plays a main role in autoimmune disease for the recognition of T cells located in specific target tissues. Type-II collagen is a well-defined tissue-specific protein which is glycosylated about 5–30%.7 Glycosylated peptides could facilitate the binding mechanism of protein to immune cell lines with the help of MHC class 2 molecules.8–10 The glycoprotein content of collagen may vary based on types and molecular weight of collagen from different species.
Recently, there has been reinstated interest in type-II collagen autoimmunity as a cause of arthritis. The immune response against type-II collagen is considered as a vital factor for the treatment of autoimmune disease such as arthritis. It was reported that immunization of rats with native type-II collagen could consequence in an inflammatory polyarthritis.11 It is believed that both cell-mediated and humoral immunity to collagen is accountable for the development of arthritis. Though the immune response of collagen in rats has been studied in some extend, the pathogenesis of the arthritis is still not understood. Indeed, some researchers have emphasized cell-mediated immune mechanisms of type-II collagen in arthritis etiology,12 while others have highlighted humoral mechanisms.13 The arthritogenicity and tolerogenicity effect of marine low MW type-II collagens have been investigated recently by few researchers.3,14 In 1982, the first collagen polypeptide was established with a MW of 59 kDa for bone calcification.15 Later, Xi et al.14 reported that low MW collagen with 28 kDa could easily recognized by T cells and induction of immune tolerance by collagen epitopes was closely related with degree of hydroxylation and percentage of glycosylation. Furthermore, Myers et al.16 reported that low MW polypeptides from the T-cell determinant of human collagen possess the ability to suppress T-cell activation in rheumatoid arthritis. Recently, some other studies have been initiated to investigate the biomedical application of type-II collagen peptides from different fish species.17,18 However, up to our knowledge, no studies of fish type-II collagen and its low MW collagens immunity in terms of in vitro tolerance and apoptotic regulatory gene expression of immune cells (T-leukemia cells) have been reported. Therefore, as a first step, we undertook a detailed study of the specific immune response of fish type-II collagen in T cell lines in order to carry out the further experiment in collagen immunization for the treatment of arthritis in vivo. With the above connections, the present study was designed to investigate the impact of molecular weight and glycoprotein content of collagens for tumour necrosis factor receptor activation in terms of extrinsic apoptotic pathway mechanism.
2. Experimental section
2.1. Materials
Whale shark cartilage (Rhincodon typus) was procured from M/s. Yueqing Ocean Biological Health Care Product Co. Ltd. Zhejiang, China. They were then washed with tap water, adherent muscle removed manually, cut in to small size and used for the collagen isolation. All the chemicals and reagents used in this study were analytical grade and purchased from commercial vendors.2.2. Preparation of collagen and low MW polypeptide
Pepsin soluble type-II collagen (CII) from the shark cartilage was extracted and purified according to our previous methods.4 Three millilitres of collagen sample (100 mg ml−1) in 0.5 M acetic acid was used for the enzymatic hydrolysis and the reaction was carried out by an addition of enzyme, thermolysin from Bacillus thermoproteolyticusrokko (EC number: 3.4.24.27) (Sigma-Aldrich, Shanghai, China) at a level of 0.1% (w/w) of the protein content. The enzymatic reaction was performed with continuous stirring with optimum temperature (70 °C) and pH (8.0) for 60 min. The enzyme activity was terminated by addition of EDTA (20 mM). After enzyme inactivation, the hydrolysate was applied to a gel filtration column using Sephadex G-50 coarse and the separation was followed as per our previous method.72.3. Characterization of low molecular weight collagenous polypeptide
The protein profile was analysed using SDS-PAGE according to the method of Laemmli.19 Amino acid profile, Fourier transform infrared spectroscopy (FTIR), DPPH radical scavenging activity and reducing power were determined as per our earlier reported method.72.4. Cell culture
Human acute T-lymphocyte leukemia cell lines (6T-CEM) were from Cell Resource Center of Shanghai Institute of Biological Sciences, Chinese Academy of Science (Shanghai, China). The cell lines were cultured in RPMI1640 medium (Gibco) supplemented with 10% fetal bovine serum (Gibco) at 37 °C in a humidified atmosphere with 5% CO2. Cell viability was estimated by trypan blue dye exclusion.2.5. Cytotoxicity
Microtiter plates (96-well, Costar) were coated overnight at 4 °C with different concentration of collagen (0.01, 0.1, 1, 10, 50 and 100 μg). Controls consisted in uncoated wells. Wells for each condition were done in triplicates. Cells were then added at 1 × 105 cells per well and incubated at 37 °C for 2 days. Viable cells were detected by adding MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) to a final concentration of 0.5 mg ml−1 for 2 h at 37 °C. The supernatant was eliminated and 250 μl of dimethyl sulfoxide (DMSO) was added to each well in order to solubilize the blue crystals of formazan that had been formed. The absorbance was then measured at 570 nm using a plate reader.2.6. Cell adhesion assay
Cell adhesion to collagen adsorbed to microtiter plates was performed as previously described with slight modification.20 Briefly, 96-well plates (Costar, flat bottom) were coated overnight at 4 °C with native collagens. Dose–response curves were obtained from coating with dilution series of collagen solutions (0.01, 0.1, 1, 10, 50 and 100 μg). Wells were then saturated with 1% heat denatured BSA for 30 min. Cells suspended in serum-free medium were added to the wells (2 × 105 cells) and incubated for 4 h at 37 °C. The non-adherent and loosely attached cells were removed by gently washing the wells with aliquots of 1× PBS. Adherent cells were fixed with 5% glutaraldehyde for 20 min and washed with distilled water. Cells were stained with 0.1% (w/v) crystal violet for 60 min at room temperature and the wells were washed with distilled water. The dyes attached in the cells were solubilized with 10% acetic acid at 150 rpm for 5 min and the absorbance read at 570 nm using a plate reader.2.7. Quantification of FAS/APO-1, IL-2, IL-6, Casp-3 and Casp-8 level
Microtiter plates (96-well, Costar) were coated with series dilution of collagen (0.01, 0.1, 1, 10, 50 and 100 μg). Controls consisted in uncoated wells. Wells for each condition were done in triplicates. Cells were then added at 1 × 105 cells per well and incubated at 37 °C in a humidified atmosphere with 5% CO2 for 24 h. Then the level of apoptosis regulatory receptor (FAS/APO-1), cytokines (IL-2 and IL-6) and crucial mediators (caspase-3 and caspase-8) were measured by ELISA according to protocols of the respective ELISA kits (Shanghai Blue-Gene Biotech Co. Ltd. Shanghai, China).2.8. Apoptosis regulatory gene expression
2.9. Fluorescence microscopy
3. Results and discussion
3.1. Biochemical characterizations
Table 1 shows amino acid composition of the isolated native and low MW collagens. Glycine, alanine and proline are major amino acid constituent for the collagens, which were similar characteristic profile to other collagens reported by several researchers.4,22,23 The native collagen had highly conserved hydrophobic amino acid such as isoleucine, leucine, phenylalanine and valine than low MW collagens. On the other hand, the above mentioned hydrophobic amino acid levels were higher in 40 kDa collagens than that of other low MW collagens. Variation of amino acid composition between native and low MW collagens might be due to proteolytic digestion by thermolysin and also removal of certain amino acid residues during the purification process involved in this study. In 1980, a study revealed that the hydrophobic characteristic of a protein could vary based on its size.24 It is believed that highly conserved hydrophobic amino acids play a key role in the structure and functional properties of protein.| Amino acids content | CII | 57 kDa | 40 kDa | 25 kDa |
|---|---|---|---|---|
| Asp | 49.07 | 50.70 | 67.62 | 49.15 |
| Thr | 26.28 | 17.60 | 24.33 | 17.94 |
| Ser | 32.83 | 30.13 | 53.52 | 30.38 |
| Glu | 87.43 | 87.40 | 79.96 | 81.04 |
| Gly | 289.84 | 329.90 | 341.92 | 283.73 |
| Ala | 96.51 | 97.47 | 98.49 | 91.35 |
| Cys | 2.36 | 8.18 | 0.13 | 7.81 |
| Val | 31.39 | 23.62 | 23.24 | 20.75 |
| Met | 18.35 | 6.75 | 3.93 | 3.97 |
| Ile | 25.70 | 12.75 | 18.23 | 10.71 |
| Leu | 81.67 | 51.43 | 64.11 | 59.84 |
| Tyr | 8.04 | 23.52 | 48.55 | 38.24 |
| Phe | 28.06 | 17.95 | 20.58 | 16.56 |
| Lys | 25.40 | 17.13 | 16.61 | 16.99 |
| His | 12.42 | 10.20 | 5.36 | 5.43 |
| Arg | 43.12 | 36.83 | 46.73 | 60.23 |
| HYP | 58.03 | 66.23 | 56.87 | 59.75 |
| Pro | 98.83 | 100.11 | 88.04 | 99.89 |
| Total | 1000 | 1000 | 1000 | 1000 |
Substantially, it mainly contributes for gene activation, G protein coupling, and receptor oligomerization. In another study, it was reported that polypeptide with above 50% hydrophobic amino acids possessed antimicrobial and hemolytic activities.25 Moreover, Wieprecht et al.26 disclosed the importance of hydrophobicity for enhanced biding of the peptide helix core to the lipid acyl chains of the cell membrane. From the above debate, it was expected that collagen with 40 kDa may possess better bioactivity than other low MW collagens. The protein pattern and molecular weight collagen extracted from shark cartilage was analysed by SDS-PAGE, showing the typical patterns of a type-II collagen with three identical chains (α1)3. Further, the electrophoretic protein pattern confirmed that three different low molecular weight collagen polypeptides were successfully separated in this study with molecular weight of 57, 40 and 25 kDa (Fig. 1). The band intensity of 40 kDa collagen was relatively higher than that of other low MW collagens, which reveals that proteolytic digestion of shark type-II collagen by thermolysin enzyme under present experimental conditions was effectively produced the collagen with 40 kDa.
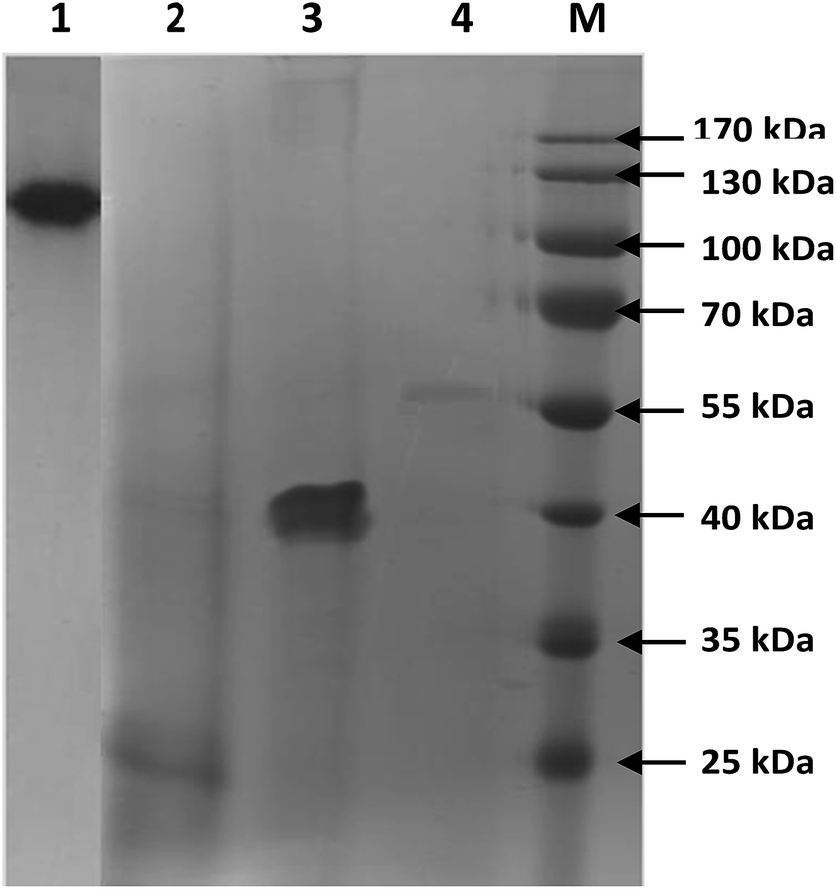 | ||
| Fig. 1 Molecular weight distribution of type-II collagens. Lane 1: type-II collagen; lane 2: 25 kDa collagen; lane 3: 40 kDa collagen; lane 4: 57 kDa collagen; lane M: marker. | ||
3.2. Functional groups and secondary structure characterization
The FTIR peak absorption pattern of amide I, II and III are directly associated with the shape and structural configuration of a polypeptide. The FTIR spectra of native and low MW collagens had different absorption peak in major amide bands due to changes in the crosslinks between peptide and functional groups as a result of hydrolysis process by thermolysin. Amide-A peak of type-II, 57, 40 and 25 kDa collagens was observed at 3368, 3392, 3393 and 3399 cm−1 respectively (Fig. 2), which was generally connected with N–H stretching coupled with the hydrogen bond of a carbonyl group in a peptide chain. In general, N–H stretching vibration occurs in the range of 3400–3440 cm−1.5 Major peaks in the spectra of collagens from shark cartilage were similar to other reports.4,5,27 Amide-B peak was observed at 2997, 2997, 2928 and 2946 cm−1 for type-II, 57, 40 and 25 kDa collagens, respectively. This peak represents the asymmetric stretching vibration of![[double bond, length as m-dash]](https://www.rsc.org/images/entities/char_e001.gif) C–H as well as NH3+. Thermolysin hydrolysis caused major shifts in amide-B peaks between native collagen and its polypeptides due to the changes in the CO, C–H and NH2 stretching in the collagen molecules.
C–H as well as NH3+. Thermolysin hydrolysis caused major shifts in amide-B peaks between native collagen and its polypeptides due to the changes in the CO, C–H and NH2 stretching in the collagen molecules.
 | ||
| Fig. 2 Fourier transform infrared spectra of type-II collagen and low molecular weight collagenous polypeptide. CII: type-II collagen; 57, 40 and 25 kDa: low MW collagens. | ||
Amide-I peak was appeared at 1666, 1653, 1644 and 1631 cm−1 for type-II, 57, 40 and 25 kDa collagens, respectively. Amide-I band is associated with stretching vibrations of carbonyl groups in peptides, being the most important factor in investigating the secondary structure of a protein. Muyonga et al.28 reported that amide-I region is generally used for the analysis of the secondary structure of proteins, which is primarily due to the CO stretching vibration of the peptide linkages. The amide-I peak of native collagen was shifted to lower frequency in low MW collagens, which was due to alteration in the triple helical structure by CO stretching vibration of the peptide linkages because of thermolysin hydrolysis process and changes in the amino acid composition.29
The characteristic peak of the amide-II region for type-II, 57, 40 and 25 kDa collagens was noticed at 1550, 1546, 1546 and 1542 cm−1, respectively, which are connected to the C–N stretching vibration and the N–H in-plane bend. Similar to the present study, Muyonga et al.28 observed the amide-II peak for Nile perch skin collagen at 1540–1558 cm−1. The above results clearly established that the triple helix structure, intermolecular cross-linkages and molecular order of native and low MW collagens varied based on crosslinking pattern of amino acid residues and the level of amino acid composition.
3.3. Glycoprotein composition
As shown in Table 2, glycoprotein content of native collagen was ranged between 5.10–16.1%, which was higher than low MW collagens. It has been reported that collagen isolated from shark cartilages had the carbohydrate contents ranged from 5.36 to 32.30%.4 The glycoprotein content of low MW collagens such as 57, 40 and 25 kDa was ranged between 2.4–3.3, 1.6–2.3, and 1.4–2.0, respectively. Among the low MW collagens, 57 kDa collagen had high content of glycoprotein than others. One of the primary physiological functions of glycoproteins in tissues is to protect the organism against toxins and they are act as a barrier in capillary endothelial cells which form the brush border of the intestine and the blood–brain barrier.30| Samples | Absorbance at 550 nm | CHO content (%) |
|---|---|---|
| Lysozyme | 0.112 | 0 |
| BSA | 0.170 | Trace amount |
| Ovalbumin | 0.397 | 3.20 |
| Apo-transferrin | 0.468 | 5.80 |
| Fetuin | 3.073 (10×) | 22.90 |
| Alpha acid glycoprotein | 2.473 (10×) | 41.40 |
| Type-II (0.25 mg ml−1 and 2.5 mg ml−1) | 0.130/0.414 | 5.10–16.17 |
| 57 kDa (0.25 mg ml−1 and 2.5 mg ml−1) | 0.043/0.295 | 2.43/3.39 |
| 40 kDa (0.25 mg ml−1 and 2.5 mg ml−1) | 0.029/0.205 | 1.65/2.36 |
| 25 kDa (0.25 mg ml−1 and 2.5 mg ml−1) | 0.027/0.177 | 1.43/2.04 |
Glycoproteins were described as the ATP-dependent transporter responsible for efflux of chemotherapeutic mediators from resilient cancer cells.31 It has also been established that glycoprotein is a crucial agent for the identification and recognition of T cell in order to treat rheumatoid arthritis in animal models.32 Furthermore, Merly and Smith33 demonstrated that the relationships between glycosylation pattern of collagen and the development of arthritis are crucial factors assumed the high number of potential glycosylation sites found in collagen. According to the substantiation described above, the higher glycoprotein content of collagens is considered as potential factors for effective cellular response and possible toxicity effect in human immune cell lines.
3.4. Scavenging and reducing power activity
The oxidation protective capabilities of native and low MW collagens were analyzed by determining the DPPH radical scavenging capacity and the reducing power. As shown in Fig. 3a, native and low MW collagens showed the radical scavenging activity in dose-dependent manner, which were lower than that of commercial antioxidants such as BHT and vitamin C. The native collagen had higher scavenging power than other low MW collagens and among the collagen polypeptide, 40 kDa collagen showed better scavenging power. The scavenging power of collagen and collagen polypeptide prepared from Spanish mackerel skin, hemp abalone foot and scallop adductor muscle have been previously documented by several researchers.34,35 | ||
| Fig. 3 Antioxidant (DPPH (A) reducing power (B)) activity of whale shark type-II collagen and low MW collagenous polypeptide. | ||
The reducing power of native collagen, 57, 40 and 25 kDa collagen was ranged between 0.07–0.135, 0.06–0.07, 0.059–0.099 and 0.06–0.071 at 700 nm, respectively (Fig. 3b), which was similar to the reducing power reported for chickpea and hemp protein hydrolysates.35,36 In the present study, the reducing power of collagens was dose-dependent manner and the values were increased with increasing the collagen concentration from 6.25 to 25 μg. It has been reported that certain specific amino acid residues such as Pro, Lys His, Met, Cys, Ala or Gly and other hydrophobic amino acids together with the higher glycopeptides content of collagen might be responsible for the higher antioxidant effects in comparison with other antioxidant peptide.37–39 Therefore, the higher content of glycoprotein and conserved hydrophobic amino acids such as isoleucine, leucine, phenylalanine and valine in native collagen than low MW collagens might be a possible substantiation for the higher antioxidant activity of native collagen.
However, the above justification of higher glycoprotein content could not claim the higher antioxidant activity of 40 kDa collagen than that of 57 kDa collagen. This might be due to variation in the amino acid sequence and composition between collagens.40 It was reported that squid hydrolysate expressed the highest antioxidant activity in spite of the lower content in hydrophobic amino acids and some amino acids such as aromatic residues, His, Met, Cys, Pro, Ala or Gly are believed to enhance radical scavenging activities of antioxidant peptides.3 Therefore, in conclusion we revealed that the antioxidant activity of native and low MW collagens may not only influence by their glycoprotein content, hydrophobic and aromatic amino acids, and also by their amino acid sequence and compositions.
3.5. Cytotoxicity and FAS/APO-1 level in T-cells
As a main objective of the present study, we determined the in vitro tolerance effect for the possible use of collagens in biomedical aspects to replace mammalian collagen. In order to confirm the toxicity, human acute T-lymphocyte leukemia cell lines (6T-CEM) were cultured for 2 days with different concentration of collagens and its activity was compared with mammalian collagen as positive control. The result shows that the total cell viability of both fish and mammalian collagen treated immune cell lines was rapidly decreased compared to control cell lines (without any treatment) and the pronounced effect was achieved with mammalian and 40 kDa collagens (Fig. 4). The cell inhibition rate were increased with collagen concentration from 0.01 to 10 μg after that it was slightly increased.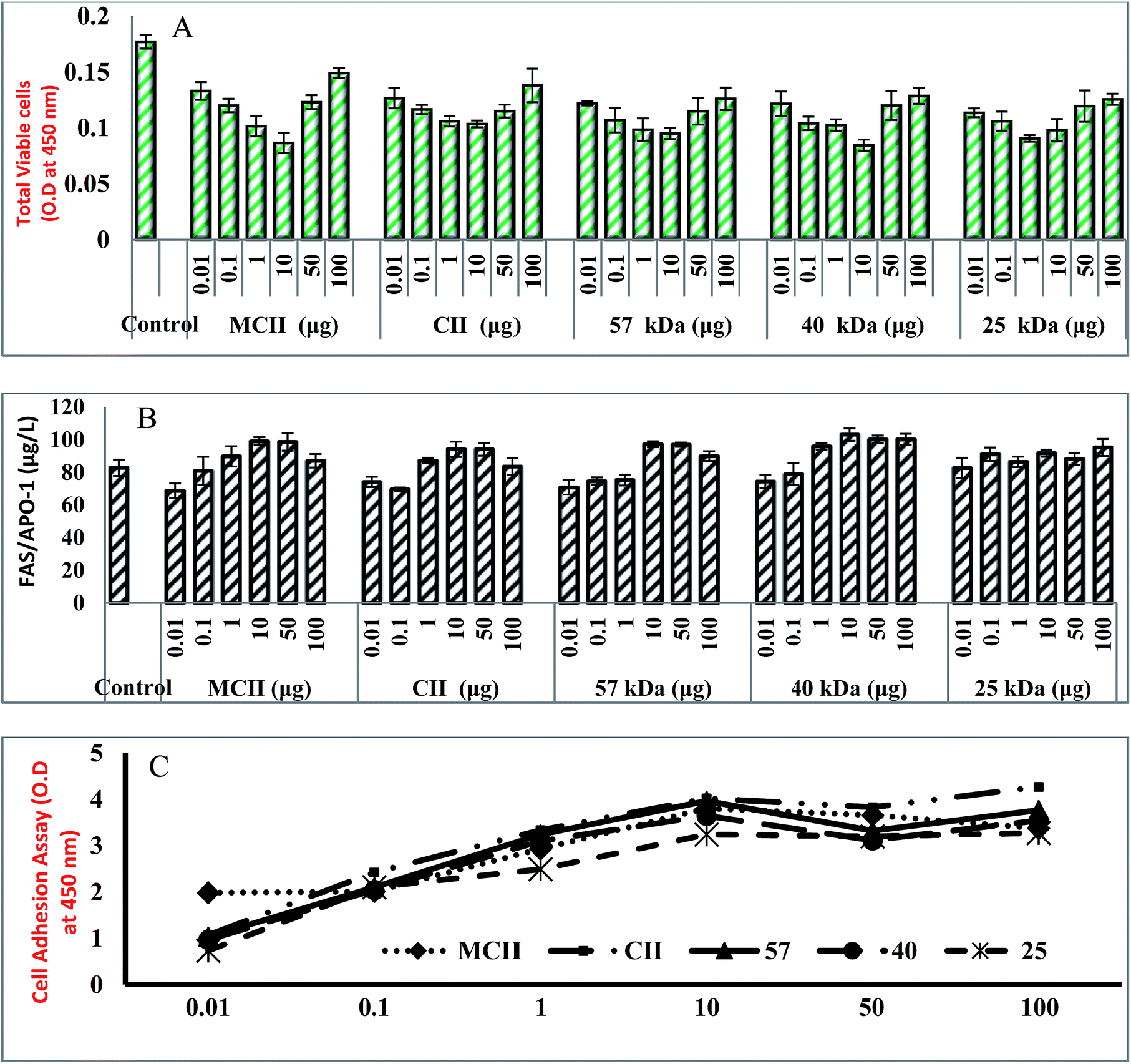 | ||
| Fig. 4 Cytotoxicity (A), Fas/Apo-1 (B) and cell adhesion (C) activity of whale shark type-II collagen and low molecular weight collagenous polypeptide. | ||
The possible reason might be that the increased concentration of collagens after certain level might competitively bind with immune cell receptors leads to improper activation of cellular death (FAS) receptor. Fas/APO-1 (apoptosis antigen-1) is a cell-surface protein capable of inducing apoptosis in many cell types through specific antibody entrapment. The immune cell lines treated with 0.01 and 0.1 μg of collagens could not produce high level of FAS/Apo-1 than control cell lines (Fig. 4). But, the level of FAS/APO-1 was high in cells treated with 1 to 50 μg collagens. The higher Fas/Apo-1 level was achieved with 10 μg of collagen, which reveals the capability of collagens to induce apoptosis in T cell lines. The present observation was in accordance with the cellular toxicity effect of collagens prepared from Blue shark cartilage.3 In the present study, the level of FAS/APO-1 was quite higher in 40 kDa collagen treated cell lines than mammalian collagen treated cell lines. From the above findings, it was concluded that induction of apoptosis and an activation of apoptotic cellular receptors in immune cell lines might be influenced by the molecular size, weight and concentration of collagen.
3.6. Cell adhesion assay
To adapt the approach of cell toxicity and the cellular surface receptors interaction, the cell adhesion assay was performed with different collagen concentrations. The different binding properties of collagen on immune cell lines showed that collagens efficiently adhere to T cells and the attachment of collagens with T cellular receptors was dose dependent-manner (Fig. 4). When compared to all collagens at same concentration, the native collagen could efficiently adhere to the immune cell receptors due to high concentration of glycoprotein content which is a fundamental mediator for the recognition of immune cellular receptor (Fig. 5). Half-maximal cell attachment was obtained with 10 μg collagens at which cells reached maximum binding. In accordance with the present study, it was stated that jellyfish and rat tail collagen had optimum concentrations of 20 μg and 5 μg against osteoblastic cells or fibroblasts, respectively.41 A detailed study about the effect of different concentration of type-II collagen (0.13–270 μg ml−1) and different number of chondrocyte cell line (2500–20![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 000 cells per wells) on cell adhesion assay was performed by Kuschel et al.42 and they stated that the number of cell binding increased with increasing coating concentrations of collagen and an optimum cell attachment was attained at 4.5 μg per 2500 cells per well, 2.9 μg per 5000 cells per well, 3.1 μg per 10000 cells per well, and 2.5 μg per 20000 cells per well coating concentrations, which was possibly due to either saturation of nitrocellulose surface with protein or cells that could not increase their avidity to the protein at these specific high coating concentrations. A group of certain specific integrin type cell receptor plays a vital role for the identification of specific recognition sites on collagen triple helical structures.43 The above results concluded that the different adhesion pattern of collagens could be attained by specific binding mechanism between immune cell receptors and collagens.
000 cells per wells) on cell adhesion assay was performed by Kuschel et al.42 and they stated that the number of cell binding increased with increasing coating concentrations of collagen and an optimum cell attachment was attained at 4.5 μg per 2500 cells per well, 2.9 μg per 5000 cells per well, 3.1 μg per 10000 cells per well, and 2.5 μg per 20000 cells per well coating concentrations, which was possibly due to either saturation of nitrocellulose surface with protein or cells that could not increase their avidity to the protein at these specific high coating concentrations. A group of certain specific integrin type cell receptor plays a vital role for the identification of specific recognition sites on collagen triple helical structures.43 The above results concluded that the different adhesion pattern of collagens could be attained by specific binding mechanism between immune cell receptors and collagens.
 | ||
| Fig. 5 A schematic diagram for receptor binding mechanism of collagen to induce apoptosis in T-lymphocyte leukemia cell line. | ||
3.7. Level of cytokines and caspases in T-cells
Cytokines are smaller proteins released from immune cells under specific condition triggered by antigen, which are mainly involved in cell signalling pathway where it transfer the signal to other cells in order to initiate the defence system. The immune cell lines treated with above 1 μg of mammalian and fish collagens could produce high level of cytokines such as interleukin-2 (IL-2) and interleukin-6 (IL-6) when compared to control cell lines (Fig. 6A and B). The concentration of IL-2 was more or less similar between mammalian and fish collagens at 10 μg, however, IL-6 level was significantly increased in cell lines treated with 40 kDa at 10 μg than that of other collagens and mammalian collagen treated cell lines. The main principle for biocompatibility is suppression of cellular response or less production of antibody against orally administered fish collagen. One of the crucial mechanisms of active cellular suppression is the secretion of down regulating or suppressive cytokines such as IL-2, IL-4, IL-6 and TGF-β.44 It has been reported that, the delivery of soluble peptides adsorbed to chitosan increases antigen release and maintains a sustained production of IL-10 in respiratory mucosa, assisting stimulation of oral tolerance.45 | ||
| Fig. 6 Level of cytokines (IL-2 (A), IL-6 (B)) and caspases (caspase-3 (C) and caspase-8 (D)) in T cell lines after the treatment with type-II collagen and low molecular weight collagenous polypeptide. | ||
Caspases play a pivotal role in the stimulation and maintenance of apoptosis, necrosis, and inflammation in immune cells. The level of caspases (caspase-3 and caspase-8) was higher in collagen treated cell lines when compared to control cells, and the higher level of caspases production achieved with the concentration of 10 μg (Fig. 6C and D). Interestingly, the level of caspase-3 production was high in those cell lines treated with fish collagen than that of mammalian collagen, which reveals that the fish collagen could efficiently establish cellular suppression than mammalian collagen. The collagen binding with FAS receptors may establish a series of immune response such as activation of caspase cascade pathway where caspase-8 activates caspase-3, a vital factor for cellular apoptosis. Caspase-3, caspase-8, and caspase-9 are the major enzymes which promote cell death via DNA fragmentation and inactivation of the proteins that protect cells against apoptosis.46,47 From the above results, it was established that fish collagen possess more or less similar or even better active cellular suppression than mammalian collagen and the tolerance could be achieved with 10 μg of collagens.
3.8. Genetic expression of immune cells and fluorescence microscopy
In general, the Ct (cycle threshold) value of RT-PCR is used to measure the target desired genetic expression in biological systems. The Ct value is simply defined as the number of thermal cycles required to exceed background fluorescent signal (threshold value) and it is inversely proportional to the amount of target nucleic acid in the sample, which means the lower Ct value represents higher expression of target gene. The fish and mammalian collagen treated cell lines had higher apoptotic gene such as FAS, FAS-L, CASP-3, CASP-8, IL-2 and IL-6 expressions than control cell lines (Fig. 7). The present observation was in accordance to the previous studies carried out with acute myeloid leukemia (K562, U937, and HL-60 cell lines of Mcl-1 gene) cell lines by lycorine treatment and T-leukemia (MOLT-4 and 6T-CEM) or lymphoma cell lines (H9 and HUT102) by bortezomib treatment.48,49 Similar to our study, Chaoqin et al.50 used leukemia cell lines (6T-CEM) in order to investigate the apoptosis promoting gene (ICE, bax, bcl-XL and bcl-Xs) and anti-apoptotic gene (bcl-2) expression induced by allicin. In the present study, the gene expressions of Casp-3 (Ct-25.3), IL-2 (Ct-29.6) and IL-6 (Ct-32.1) were high in fish collagen treated cell lines than mammalian collagen. On the other hand, among the low MW collagens, the apoptotic gene expression of FAS (Ct-30.9), Casp-3 (Ct-26.2), Casp-8 (Ct-28.4), and IL-6 (Ct-32.3) was high in 40 kDa collagen treated sample. The higher apoptotic gene expression by 40 kDa collagen might be related to the specific apoptotic regulatory membrane receptor binding mechanism in immune cells. The above results clearly demonstrated that administration of collagens could possibly induce apoptosis regulatory genetic expression which leads to down-regulation of immune responses. | ||
| Fig. 7 Apoptosis regulatory gene expression of 6T-CEM cells by real-time PCR. | ||
In order to substantiate the above findings, we further carried out fluorescence microscopy to identify the structural changes of immune cell membrane and its nuclei upon treatment with collagens. The fluorescence microscopic pictures clearly demonstrated that the apoptosis rate of 6T-CEM cell lines increased significantly with the treatment of collagens than control (Fig. 8). The apoptotic rate was higher in CII treated cell lines than that of cells treated with mammalian collagen. Collectively, these results further confirmed that the higher apoptosis gene expressions of fish collagens in 6T-CEM cell lines. Chen et al.3 opined that the apoptosis of 6T-CEM cell lines induced by CII may result in the depletion of antigen-specific reactive T cells, which is also vital in inducing specific immune tolerance. Michaelsson et al.51 demonstrated that the presence of a glycosylated immunodominant T cell determinant is strictly dependent on the presence of carbohydrates. Further they suggested that carbohydrates on post-translationally modified tissue-specific proteins might be a critical factor for development of autoimmune disease. It has been reported that T cells are capable to recognize glycosylated peptides bound to MHC class II.8,10 In the present study, higher content of glycoprotein in native collagen than that of low MW collagens supports the enhanced apoptosis regulatory gene expression of collagen. From the above findings, we concluded that the apoptosis of T cells was initiated by triggering membrane death (Fas) receptor through collagen binding which leads to establish caspase cascade pathway.
 | ||
| Fig. 8 Fluorescence microscopic structure of T cells stained with FITC and DAIP. The cell specified by arrows is apoptotic cell. | ||
4. Conclusions
To sum up, the isolated collagen from shark cartilage was characterized as type-II and low molecular weight collagen was successfully separated under present experimental conditions. The biochemical properties such as amino acid composition and FTIR spectra of native and low MW collagens were not similar due to changes in their primary and secondary structure by proteolytic hydrolysis. The higher antioxidant properties exposed that collagen polypeptide with MW of 40 kDa obtained by thermolysin hydrolysis may be considered potential alternative biomaterial to synthetic antioxidants. In vitro tolerance and apoptosis regulatory gene expression of fish collagen were investigated as an intension of to replace the use of mammalian collagen in biomedical applications. Immune cellular apoptosis and cell focal adhesion were dose dependent manner, the optimal coating concentration being approximately 10 μg. The increased focal adhesions of immune cells treated with highly glycosylated fish native and low MW collagens revealed that the recognizing patter of immune cellular receptors was mainly influenced with glycoprotein content and molecular weight of collagen. The activation of Fas cell surface death receptor was highly influenced by 40 kDa collagen. The extrinsic apoptosis induction in T cells was based on the MW of collagens and uniting mechanism between glycoprotein and death (Fas/CD95) receptor. Accordingly, the extracted native collagen could possess less toxicity in terms of active immune cellular suppression than mammalian collagen and also by understanding the better biocompatibility properties and apoptotic regulatory gene expression profile of low MW collagens could disclose new therapeutic biomaterials for the treatment of inflammatory autoimmune disease, rheumatoid arthritis.Acknowledgements
This work received financial support from Shanghai High Technology Research and Development Program (No. 14431906000 and No. 15410722500).References
- A. Jongjareonrak, S. Benjakul, W. Visessanguan, T. Nagai and M. Tanaka, Food Chem., 2005, 93, 475–484 CrossRef CAS.
- P. M. Gilsenan and S. B. Ross-Murphy, Int. J. Biol. Macromol., 2001, 29, 53–61 CrossRef CAS PubMed.
- L. Chen, B. Bao, N. Wang, J. Xie and W. H. Wu, Pharmaceutical, 2012, 5, 339–352 CAS.
- E. Jeevithan, B. Bao, Y. Bu, Y. Zhou, Q. Zhao and W. H. Wu, Mar. Drugs, 2014, 12, 3852–3873 CrossRef CAS PubMed.
- A. Veeruraj, M. Arumugam and T. Balasubramanian, Process Biochem., 2013, 48, 1592–1602 CrossRef CAS.
- A. R. Poole, Cartilage in Health and Disease, in Arthritis and Allied Conditions: A Textbook of Rheumatology, ed. W. J. Koopman and L. W. Moreland, Williams and Wilkins, Baltimore, MD, USA, 15th edn 2005, pp. 223–269 Search PubMed.
- E. Jeevithan, Z. Jingyi, N. Wang, L. He, B. Bao and W. H. Wu, Process Biochem., 2015, 50, 463–472 CrossRef CAS.
- R. M. Chicz, R. G. Urban, J. C. Gorga, D. A. A. Vignali, W. S. Lane and J. L. Strominger, J. Exp. Med., 1993, 178, 27–47 CrossRef CAS PubMed.
- C. V. Harding, J. Kihlberg, M. Elofsson, G. Magnusson and E. R. Unanue, J. Immunol., 1993, 151, 2419–2425 CAS.
- G. Y. Ishioka, A. G. Lamont, D. Thomson, N. Bulbow, F. C. A. Gaeta and A. Sette, J. Immunol., 1992, 148, 2446–2451 CAS.
- D. E. Trentham, A. S. Townes and A. H. Kang, J. Exp. Med., 1977, 146, 857–868 CrossRef CAS PubMed.
- D. E. Trentham, R. A. Dynesius and J. R. David, J. Clin. Invest., 1978, 62, 359–366 CrossRef CAS PubMed.
- J. M. Stuart, M. A. Cremer, A. S. Townes and A. H. Kang, Arthritis Rheum., 1979, 12, 1344–1351 CrossRef.
- C. Xi, L. Tan, Y. Sun, F. Liang, N. Liu and H. Xue, Mol. Immunol., 2009, 46, 729–737 CrossRef CAS PubMed.
- T. M. Schmid and H. E. Conrad, J. Biol. Chem., 1982, 257, 12444–12450 CAS.
- L. K. Myers, Y. Sakurai, E. F. Rosloniec, J. M. Stuart and A. H. Kang, Am. J. Med. Sci., 2004, 327, 212–216 CrossRef PubMed.
- H. Ehrlich, Biol. Mater. Mar. Orig., 2015, 4, 69–89 Search PubMed.
- J. Xie, H. Y. Ye and X. F. Luo, Int. Food Res. J., 2014, 21, 1171–1175 CAS.
- U. K. Laemmli, Nature, 1970, 227, 680–685 CrossRef CAS PubMed.
- F. Elefteriou, J. Y. Exposito, R. Garrone and C. Lethias, Eur. J. Biochem., 1999, 26, 3840–3848 Search PubMed.
- S. Hattori, H. Fujisaki, T. Kiriyama, Y. T. okoyama and S. Irie, Anal. Biochem., 2002, 301, 27–34 CrossRef CAS PubMed.
- I. Bae, K. Osatomi, A. Yoshida, K. Osako, A. Yamaguchi and K. Hara, Food Chem., 2008, 108, 49–54 CrossRef CAS.
- M. Ogawa, M. W. Moody, R. J. Portier, J. Bell, M. A. Schexnayder and J. N. Losso, J. Agric. Food Chem., 2003, 51, 8088–8092 CrossRef CAS PubMed.
- H. Meirovitch, S. Rackovsky and H. A. Scheraga, Macromolecules, 1980, 13, 1398–1405 CrossRef CAS.
- R. E. W. Hancock and D. S. Chapple, Chemotherapia, 1999, 43, 1317–1323 CAS.
- T. Wieprecht, M. Dathe, E. Krause, M. Beyermann, W. L. D. L. Molloy and M. Donald, FEBS Lett., 1997, 417, 135–140 CrossRef CAS PubMed.
- P. Kittiphattanabawon, S. Benjakul, W. Visessanguan and F. Shahidi, LWT--Food Sci. Technol., 2010, 43, 792–800 CrossRef CAS.
- J. H. Muyonga, C. G. B. Cole and K. G. Duodu, Food Chem., 2004, 85, 81–89 CrossRef CAS.
- E. Jeevithan, B. Bao, Z. Jingyi, S. Hong and W. H. Wu, J. Food Sci. Technol., 2015, 52, 6312–6322 CrossRef CAS PubMed.
- A. Sparreboom, J. V. Asperen, U. Mayer, A. H. Schinkel, J. W. S. Dirk and K. F. Meijer, Proc. Natl. Acad. Sci. U. S. A., 1997, 94, 2031–2035 CrossRef CAS.
- V. J. Wacher, J. A. Silverman, Y. Zhang and L. Z. Benet, J. Pharm. Sci., 1998, 87, 1322–1330 CrossRef CAS PubMed.
- J. Backlund, A. Treschow, R. Bockermann, B. Holm, L. Holm and S. Issazadeh-Navikas, Eur. J. Immunol., 2002, 32, 3776–3784 CrossRef CAS PubMed.
- L. Merly and S. L. Smith, Int. Immunopharmacol., 2013, 15, 309–315 CrossRef CAS PubMed.
- C. F. Chi, Z. H. Cao, B. Wang, F. Y. Hu, Z. R. Li and B. Zhang, Molecule, 2014, 19, 11211–11230 CrossRef PubMed.
- C. H. Tang, X. S. Wang and X. Q. Yang, Food Chem., 2009, 114, 1484–1490 CrossRef CAS.
- Y. Li, B. Jiang, T. Zhang, W. Mu and J. Liu, Food Chem., 2008, 106, 444–450 CrossRef CAS.
- B. Zhu, X. Dong, D. Zhou, Y. Gao, J. Yang and D. Li, Food Hydrocolloids, 2012, 28, 182–188 CrossRef CAS.
- E. Mendis, N. Rajapakse, H. G. Byun and S. K. Kim, Life Sci., 2005, 77, 2166–2178 CrossRef CAS PubMed.
- B. Gimenez, J. Gomez-Estaca, A. Aleman, M. C. Gómez-Guillen and M. P. Montero, Food Chem., 2009, 114, 976–983 CrossRef CAS.
- H. M. Chen, K. Muramoto, F. Yamaguchi, K. Fujimoto and K. Nokihara, J. Agric. Food Chem., 1998, 46, 49–53 CrossRef CAS PubMed.
- S. Addad, J. Y. Exposito, C. Faye, S. Ricard-Blum and C. Lethias, Mar. Drugs, 2011, 9, 967–983 CrossRef CAS PubMed.
- C. Kuschel, H. Steuer, A. N. Maurer, B. Kanzok, R. Stoop and B. Angres, BioTech, 2006, 40, 523–531 CrossRef CAS PubMed.
- J. Emsley, C. Knight, R. Farndale, M. Barnes and R. C. Liddington, Cell, 2000, 101, 47–56 CrossRef CAS PubMed.
- H. L. Weiner, Immunol. Today, 1997, 18, 335–343 CrossRef CAS PubMed.
- G. Hall, L. Lund, J. R. Lamb and E. R. Jarman, J. Allergy Clin. Immunol., 2002, 110, 883–890 CrossRef CAS PubMed.
- S. Nagata, Cell, 1997, 88, 355–365 CrossRef CAS PubMed.
- N. Thornberry and Y. A. Lazebnik, Science, 1998, 281, 1312–1316 CrossRef CAS PubMed.
- X. S. Liu, J. Jiang, X. Jiao, Y. Wu, J. Lin and Y. Cai, Cancer Lett., 2009, 274, 16–24 CrossRef CAS PubMed.
- Q. L. Zhang, L. Wang, Y. W. Zhang, X. X. Jiang, F. Yang and W. L. Wu, Leukemia, 2009, 23, 1507–1514 CrossRef CAS PubMed.
- Y. Chaoqin, L. Changquan, H. Xueqiang, P. Ruiping and Z. Denghai, Chin. J. Integr. Med., 1999, 5, 126–130 Search PubMed.
- E. Michaelsson, V. Malmström, S. Reis, E. Michaëlsson, V. Malmström and S. Reis, J. Exp. Med., 1994, 1, 745–749 CrossRef.
| This journal is © The Royal Society of Chemistry 2016 |