
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceAntimicrobial potentials of natural products against multidrug resistance pathogens: a comprehensive review
Abeer H. Elmaidomy†
a,
Nourhan Hisham Shady†b,
Khaled Mohamed Abdeljawadc,
Mohamed Badran Elzamkanc,
Hussein Hykel Helmyc,
Emad Ashour Tarshanc,
Abanoub Nabil Adlyc,
Yasmin Hamdy Hussienc,
Nesma Gamal Sayedc,
Ahmed Zayed de and
Usama Ramadan Abdelmohsen*bf
de and
Usama Ramadan Abdelmohsen*bf
aDepartment of Pharmacognosy, Faculty of Pharmacy, Beni-Suef University, Beni-Suef 62511, Egypt
bDepartment of Pharmacognosy, Faculty of Pharmacy, Deraya University, Universities Zone, New Minia 61111, Egypt
cFaculty of Pharmacy, Deraya University, Universities Zone, New Minia 61111, Egypt
dDepartment of Pharmacognosy, College of Pharmacy, Tanta University, Elguish Street (Medical Campus), Tanta, 31527, Egypt
eInstitute of Bioprocess Engineering, Technical University of Kaiserslautern, Gottlieb-Daimler-Str. 49, Kaiserslautern 67663, Germany
fDepartment of Pharmacognosy, Faculty of Pharmacy, Minia University, Minia 61519, Egypt. E-mail: usama.ramadan@mu.edu.eg
First published on 13th October 2022
Abstract
Antibiotic resistance is one of the critical issues, describing a significant social health complication globally. Hence, the discovery of novel antibiotics has acquired an increased attention particularly against drug-resistant pathogens. Natural products have served as potent therapeutics against pathogenic bacteria since the glorious age of antibiotics of the mid 20th century. This review outlines the various mechanistic candidates for dealing with multi-drug resistant pathogens and explores the terrestrial phytochemicals isolated from plants, lichens, insects, animals, fungi, bacteria, mushrooms, and minerals with reported antimicrobial activity, either alone or in combination with conventional antibiotics. Moreover, newly established tools are presented, including prebiotics, probiotics, synbiotics, bacteriophages, nanoparticles, and bacteriocins, supporting the progress of effective antibiotics to address the emergence of antibiotic-resistant infectious bacteria. Therefore, the current article may uncover promising drug candidates that can be used in drug discovery in the future.
1. Introduction
Natural products have provided a major foundation for the development of antibiotics since ancient times (e.g., β-lactams, tetracycline, lincosamides, aminoglycosides, glycopeptides, and macrolides) (Fig. 1). Antibiotics have been shown to act on different targets within bacterial cells, including inhibition of cell wall synthesis (β-lactams: cephalosporins, carbapenems, penicillins, monobactams, glycopeptides), protein synthesis (binding to the 30S ribosomal subunit: tetracyclines, aminoglycosides, or binding to the 50S ribosomal subunit: lincosamides, chloramphenicol, macrolides, streptogramins, oxazolidinones), DNA or RNA synthesis (quinolones: fluoroquinolones, rifampin), metabolic pathways (sulfonamides: trimethoprim), or mycolic acid synthesis (isoniazid) (Fig. 1).1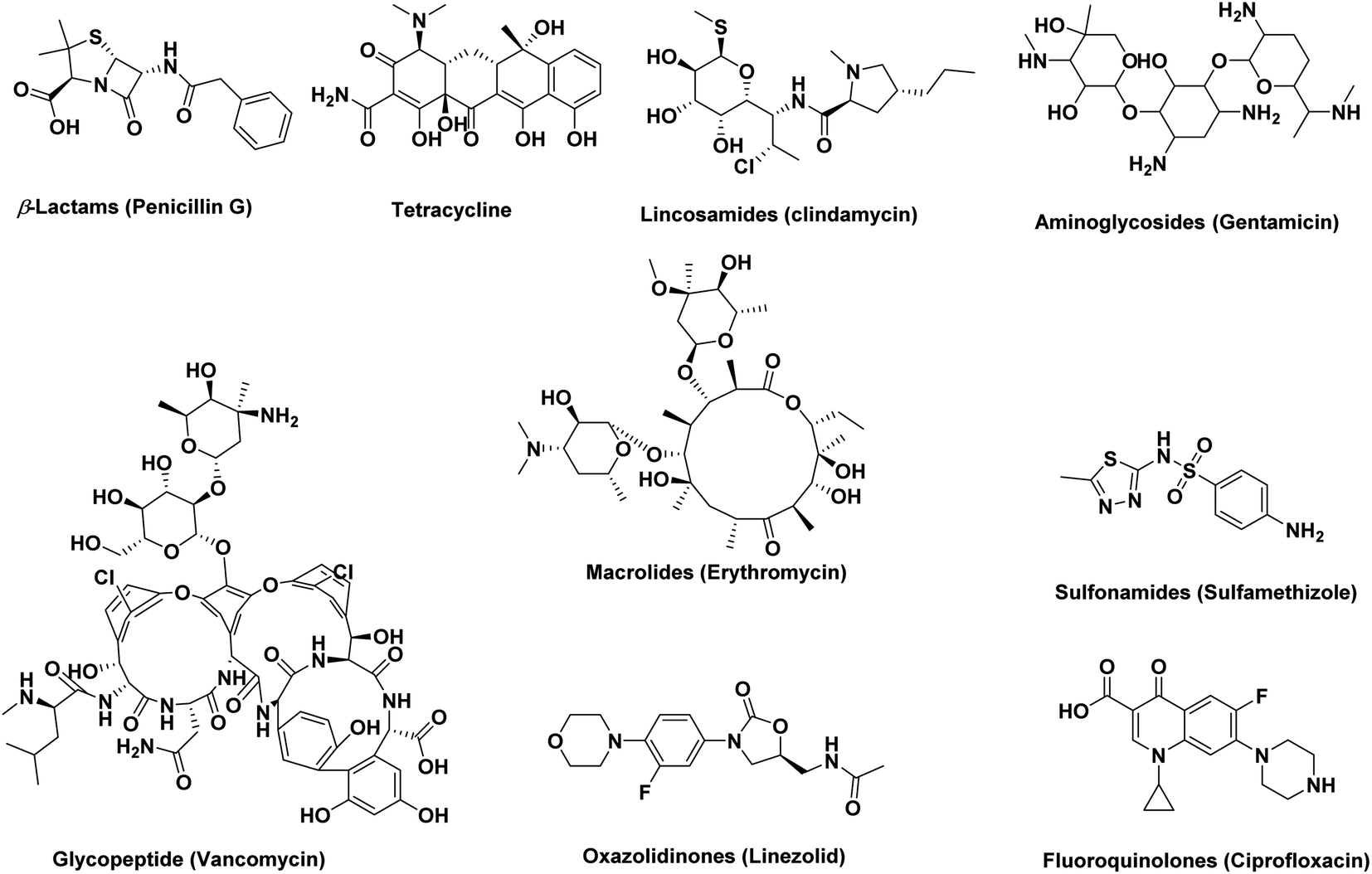 | ||
| Fig. 1 Examples of naturally occurring antibiotics classes along with three synthetic ones (sulfonamides, oxazolidinones, and fluoroquinolones). | ||
Nowadays, the rise of pathogenic different species resistant to antibiotics is one of the greatest challenges. Infections caused by multidrug-resistant (MDR) bacteria are increasingly common and represent a serious problem for the global public health. It dramatically reduces the probability of effectively treating infections and increases the morbidity and mortality associated with common bacterial diseases.2 Since the discovery of penicillin in 1928, antimicrobial resistance has been linked to antibiotic use.3 Besides, bacterial strains resistant to newly developed antibiotics have emerged recurrently.4 Therefore, antimicrobial resistance presents an ongoing challenge that requires a multifaceted approach. It is alarming since bacterial resistance continues to emerge and the rate at which antibiotics are being developed is decreasing. Antimicrobial resistance is commonly mediated through extra-chromosomal genetic elements acquisition via horizontal gene transfer.5 Low permeability of the outer membrane in Gram-negative bacteria, efflux pumps, production of degrading enzymes, biofilm formation, and modification of targets are examples of mechanisms used by bacteria to resist the toxicity of antibiotics (Table 1).5
| Drug | Drug uptake limitation | Drug target modification | Drug inactivation | Efflux pumps |
|---|---|---|---|---|
| β-Lactams | + | + | + | + |
| Carbapenems | + | |||
| Cephalosporins | + | |||
| Glycopeptides | + | + | ||
| Lipopeptides | + | |||
| Aminoglycosides | + | + | + | + |
| Tetracyclines | + | + | + | + |
| Chloramphenicol | + | + | + | |
| Lincosamides | + | + | ||
| Macrolides | + | + | ||
| Oxazolidinones | + | + | ||
| Streptogramins | + | |||
| Fluoroquinolones | + | + | + | |
| Sulfonamides | + | + | ||
| Trimethoprim | + | + |
Among the Gram-positive resistance bacteria species, Enterococcus faecium, Staphylococcus aureus, Streptococcus pneumoniae, and Enterococcus aecalis are the most frequent problem.6 While for Gram-negative resistance bacteria strains, Klebsiella pneumoniae, Escherichia coli, Acinetobacter baumannii, Pseudomonas aeruginosa, and Enterobacteria sp., have been mostly common.6 Globally, excessive use of antibiotics in animal husbandry and aquaculture, use of multiple broad-spectrum agents, and lack of good antimicrobial stewardship can be listed as the factors mostly responsible for the spread of antibiotic resistance species.7
The increase in the prevalence of antibiotic-resistant pathogens implies fewer antimicrobial agents to treat infections caused by these bacteria.8 This raises consequently the need to search for alternative drugs or methods for controlling antibiotic-resistant pathogens.
Natural products and their semisynthetic analogous have participated in a vital part in the description and expansion of antimicrobial drug, especially in the last 20 years.9 Fig. 2, where diverse terrestrial sources, including plants, fungi, lichen presented more than 80% of reported naturally derived antibiotics, Fig. 3. These products were found to act by different mechanisms controlling multi-resistant pathogens, Fig. 4. Despite the marked impact on safety, nature obtained compounds have attained specialized attention for their potential actions against diverse microorganisms. Many pure natural products along with newly synthetic analogs have confirmed their efficiencies as alternatives as antimicrobial agents against resistant infections.10 Furthermore, natural antimicrobial agents have built up considerable interest to replace the potency of non-effective antibiotics.
 | ||
| Fig. 2 Distribution of publications covering antimicrobial agents derived from natural products in the last period of research. | ||
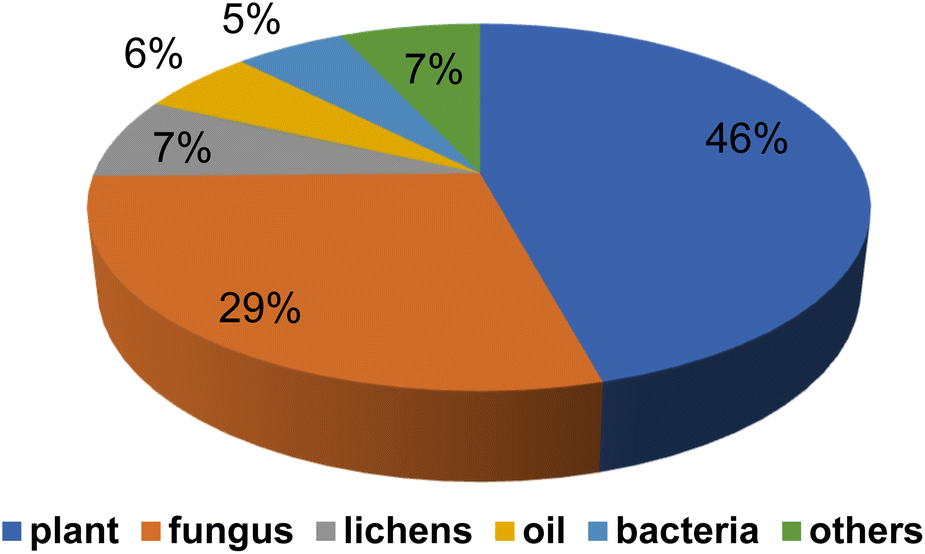 | ||
| Fig. 3 Natural products against drug resistant bacteria from diverse terrestrial sources. | ||
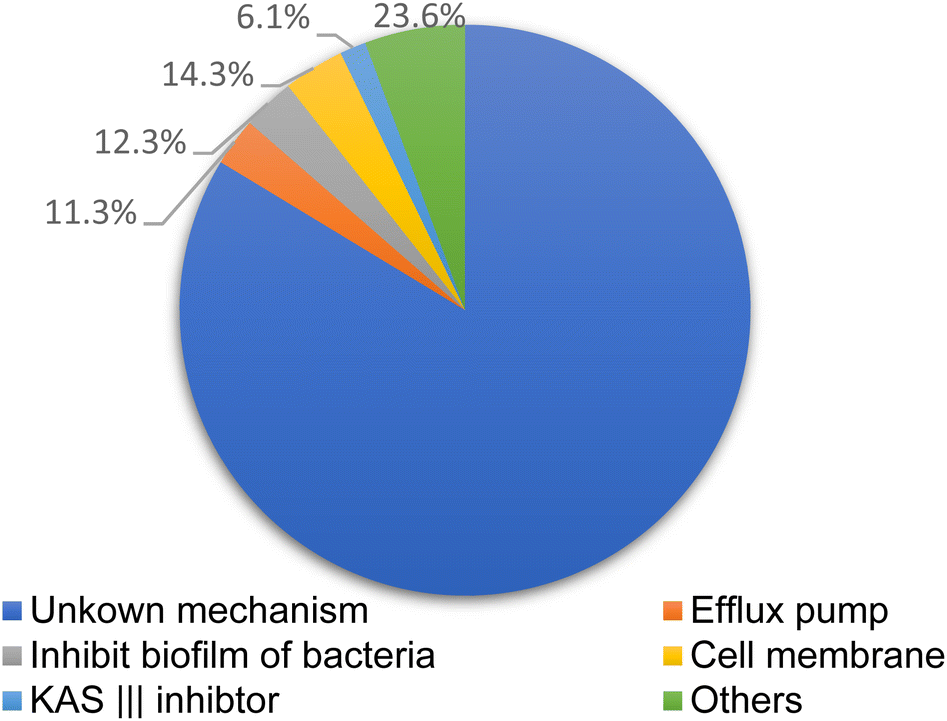 | ||
| Fig. 4 Methods for controlling multi-resistant pathogens controlling by natural products compounds. | ||
The objective of this review is to list and highlight the potential of terrestrial natural products isolated from plants, lichens, insects, animals, fungi, bacteria, mushrooms, and minerals that have been tested against the most frequent antibiotic-resistant bacteria along with describing the alternative methods that were proposed to control them. Therefore, it may reveal more drug candidates that can be used in drug industry in the near future.
2. Methods for controlling multi-resistant pathogens
2.1. Elimination of resistant plasmids
The formation, transfer, and transmission of resistant plasmids are important mechanisms that cause extensive antibiotic resistance, which play a major role in the dissemination of resistance genes.11 Consequently, one of the effective mechanisms for decreasing antibiotic resistance is inhibition of the transfer of resistant plasmids or elimination of those plasmids.2.2. Effect on the permeability of cell membrane
Since the bacterial cell membrane prevents the transport of antibiotics and consequently affects the drug efficiency, permeability of the cell bacterial membrane could be changed through changing some ion channels. By this method, the permeability of cell membranes to transport antibiotics into bacterial through bacterial cell wall may be effective against MDR pathogens overcoming antibiotic resistance.122.3. Inhibition on the efflux pump of antibiotic-resistant bacteria
Antibiotics can easily induce the overexpression of bacterial efflux pump to force bacteria pump out more antibacterial drugs to significantly decrease drug concentration at the target site, exacerbating bacterial infection. The efflux system is observed to be present in both Gram-positive and Gram-negative bacteria.13 For example, methicillin resistant Staphylococcus aureus (MRSA) up-regulates the expression of NorA gene to increase drug excretion. NorA efflux pump belongs to MFS family and is first found in clinic to be the important mechanism of bacteria resistant to quinolone and methicillin. Efflux pump inhibitors (EPI) of bacteria could eliminate antibiotic resistance.132.4. Changes in drug targets
Peptidoglycan is the major component of the cell envelope of most bacteria. In peptidoglycan synthesis, several proteins such as Mur enzymes and PBPs were found to be the targets of antibiotics. However, changes in the structure and quantity of PBPs by bacteria play important roles in bacterial drug resistance.14 Such changes involved diverse mechanisms especially ribosomal subunits via ribosomal mutation and ribosomal subunit methylation, commonly involving the erm genes interfering with drugs ability to bind to the ribosome. Additionally, for drugs that target nucleic acid synthesis, resistance is via modifications in DNA gyrase or topoisomerase IV. For the drugs that inhibit metabolic pathways, resistance is via mutations in enzymes (DHPS—dihydropteroate synthase, DHFR—dihydrofolate reductase) involved in the folate biosynthesis pathway and/or overproduction of resistant DHPS and DHFR enzymes.12.5. Inhibition on the biofilm formation
Bacterial colonization has been widely reported which is identified as the formation of a biofilm by a bacterial community. For pathogenic organisms, formation of a biofilm protects the bacteria from the host immune system, in addition to provides protection from antimicrobial agents. The thick, sticky consistency of the biofilm matrix which contains polysaccharides, proteins, and DNA from the resident bacteria, makes it difficult for antimicrobial agents to reach the bacteria. In addition, the bacterial cells in the biofilm tend to be sessile (slow metabolism rate, slow cell division), so antimicrobials that target growing, dividing bacterial cells have little effect. An important observation about biofilms is the horizontal transfer of genes facilitated by the proximity of the bacterial cells. This results in sharing of antimicrobial resistance genes among bacterial communities.12.6. Inhibition drug inactivation
Two main ways by which bacteria can inactivate antibiotics. They are either by actual degradation of the drug or drug modification through the transfer of a chemical group to the drug chemical structure (e.g., the β-lactamases are a very large group of drug hydrolyzing enzymes, hydrolyzation of tetracycline, via the tetX gene). Drug inactivation by transfer of a chemical group to the drug most commonly uses transfer of acetyl, phosphoryl, and adenyl groups. There are many transferases have been identified that can be acted on overcoming antibiotic (drugs) resistance.12.7. Bacteriocins
Bacteriocins are antimicrobial peptides ribosomally synthesized by almost all bacterial species and have a varied mechanism of action and spectrum of activity. Many bacteriocins properties as high stability, low toxicity, and broad spectra of activity, make them good alternative to antibiotics. In addition, some bacteriocins, have a dual mechanism of action, reducing the probability of selecting resistant strains. However, bacteriocin resistance in vitro is observed, and easily developed which is mostly associated with physiological adaptation. Topical, intranasal or intravenous are the available therapeutical administration ways for bacteriocin since enzymes present in the gastrointestinal tract inactivate them.62.8. Essential oils (EOs)
Another alternative tool to control MDR pathogens are essential oils (EOs). EOs have shown antimicrobial activity against MRSA, MDR-K. oxytoca, β-lactamases and carbapenemases E. coli, erythromycin-resistant Group A streptococci, and MDR-A. baumannii. EOs when blended with antimicrobial agents, their constituents could unlock the cell membrane channels, opening the passage of antimicrobial agents to reach their internal target sites. This is a great strategy to avoid selection of resistant strains in the future. However, low water-solubility/high vapor pressure are characters that limit the EOs utilization.62.9. Quorum-sensing inhibitors (QSI)
Quorum sensing (QS) is an intercellular bacterial communication used to coordinate group behaviors in a cell density-dependent manner. At high concentrations, pathogens can switch their transcription profiles to an invasive phenotype, including genes related to antibiotic tolerance and virulence determinants, and cause disease.6 In this way, QS systems constitute important ant virulence targets, as they often regulate the expression of several virulence genes simultaneously. QSI act by inhibiting cell-to-cell communications and, consequently, disease evolution, enabling the host immune system to prevent bacterial colonization and/or to clear an established infection. This antimicrobial control relies on reducing the burden of virulence rather than killing the bacteria.6 In the last two decades, various QSI from plants, animals, and microorganisms have been characterized and animal and plant infection models have demonstrated their antibacterial efficacy against QS pathogens. QSI could thus serve as a good alternative to treat infections caused by MDR pathogens. However, its application clinically still requires more research.63. Controlling multi-resistant pathogens using terrestrial/microbiota derived natural products applications
3.1. Oil derived natural products
Most reports regarding the EOs' constituents against MDR bacteria were investigated in in vitro studies. Examples include geraniol 1, which is a monoterpenoid alcohol, that was found to efficiently increased the susceptibility of MDR-Enterobacter aerogenes, E. coli, and P. aeruginosa by becoming a potent EPI.15,16 Phenol monoterpene, carvacrol 2, was reported to inhibit biofilm formation of S. aureus, and S. typhimurium.17 In other study, researchers observed heat shock protein induction in E. coli 0157![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :H7 cells treated with carvacrol 2 with flagellin synthesis inhibition, beside to the consequent production of nonmotile cells.18 Recently, monoterpene linalool 3 reported to exhibit strong antimicrobial activity against resistant K. pneumoniae through membrane disruption.19
:H7 cells treated with carvacrol 2 with flagellin synthesis inhibition, beside to the consequent production of nonmotile cells.18 Recently, monoterpene linalool 3 reported to exhibit strong antimicrobial activity against resistant K. pneumoniae through membrane disruption.19
Additionally, farnesol 4, an isoprenoid natural acyclic sesquiterpene alcohol, showed moderate effects against Streptococcus mutans and Streptococcus sobrinus biofilm formation.20 Farnesol 4 also showed antibacterial activity against S. aureus and S. epidermidis whereby it also inhibited the biofilm development.21 Two studies conducted by Masako,22 evidenced that combinations of farnesol 4 with xylitol, which is a natural sugar alcohol, have positive effects against atopic dermatitis caused by S. aureus and successfully inhibited the biofilm production of S. aureus. Study conducted by Sayout et al., 2020 (ref. 23) evidenced that camphor 5 has been shown to be active against MRSA P637, Escherichia coli P1420, Enterobacter aerogenes P1260, Pseudomonas aeruginosa P1418, Klebsiella pneumoniae LA726, Klebsiel laoxytoca BU9399, Salmonella spp., Acinetobacter baumanii PDP533, and Enterobacter cloacae P1374.
Other compounds have also been studied (α-pinene 6, camphene 7, fenchone 8, cis-verbenol 9, borneol 10, and verbenone 11). These compounds showed a strong antimicrobial activity against most of MDR strains, except camphene 7 which was not active against MRSA, and Enterobacte rcloacae, and borneol 10 which was inactive against Salmonella spp. Sayout et al., 2020,23 also conducted that β-pinene 12, myrcene 13, Δ3-carene 14, p-cymene 15, 1,8-cineole 16, limonene 17, γ-terpinene 18, terpinen-4-ol 19, and carvone 20 even if are presented in low concentrations, they have interesting antimicrobial activity against MDR bacteria.24 reported that Salmonella typhimurium when exposed to eugenol 21 at 1% and 5% (v/v), developed increased membrane permeability followed by leakage of the cell contents. In contrast,25 reported the activity of eugenol 21 (5 mM) on Listeria monocytogenes cells results in inhibition of the uptake and utilization of glucose. Salmonella enterica serovar Thompson cells were treated with a sublethal concentration of thymol 22 (0.01%), which caused overexpression of a group of molecular chaperone proteins (DnaK, GroEL, HtpG, and the Trigger factor Tf) and outer membrane-associated proteins (OmpX and two OmpA proteins), in addition to upregulation of proteins related to citrate metabolism and ATP synthesis.26 Niu et al.27 observed that cinnamaldehyde 23 affects transcription of two acyl homoserine lactones (HSLs), 3-oxo-C6-HSL and 3-oxo-C12-HSL, and the bioluminescence of V. harveyi, which is mediated by 3-hydroxy-C4-HSL and the autoinducer-2 (AI-2). The effect of several terpenes (geraniol 1, carvacrol 2, eugenol 21, and thymol 22) in combination with penicillin against MRSA ATCC 25923 and an E. coli strain was evaluated in a study of Gallucci et al.28 The MICs of carvacrol 2, eugenol 21, thymol 22 for the MRSA strain were 15.25, 133.75, and 30.15 mg mL−1, and the MICs of for geraniol 1, carvacrol 2, eugenol 21, and thymol 22 for the E. coli strain were 222.25, 7.62, 66.82, and 15.07 mg mL−1, respectively (Fig. 5).
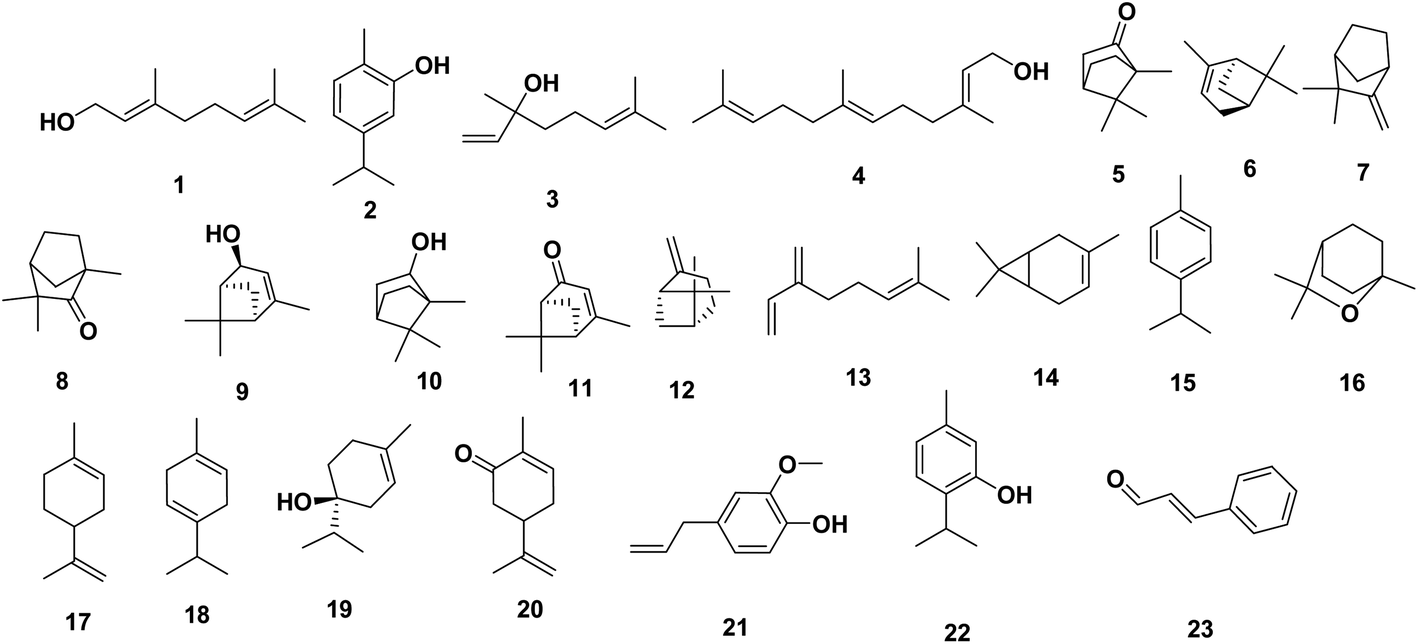 | ||
| Fig. 5 Oil derived natural products 1–23. | ||
3.2. Plants/endophytes derived natural products
Fig. 6 illustrates the chemical structure for natural derived compounds 24–110 from plants and their associated endophytes. They showed potential antibacterial activity against MDR strain mostly in in vitro studies. They may be classified phytochemically into curcuminoids (e.g., curcumin 24), chalcones and acylphloroglucinols (e.g., humulone 25, lupulone 26, xanthohumol 27, desmethylxanthohumol 28, cohumulone 29, colupulone 30), alkaloids (e.g., compounds 31–68, 160–162, and 188–189), flavonoids and isoflavonoids (e.g., 69–74, 138, and 197–200), quinonoids (e.g., 75–79), xanthones, terpenoids, and others. In the following section, such bioactive antibacterial compounds shall be discussed in detail. | ||
| Fig. 6 Plant derived natural products 24–110. | ||
Curcumin 24, a natural polyphenolic flavonoid isolated from Curcuma longa Linné., showed to have MICs against 10 MDR strains of S. aureus ranged from 125 to 250 g mL−1. In the checkerboard test, curcumin 24 markedly reduced the MICs of the antibiotics oxacillin (OXI), ampicillin (AMP), ciprofloxacin (CIP), and norfloxacin (NOR) used against MRSA. The time-kill curves showed that a combined curcumin 24 and OXI treatment reduced the bacterial counts below the lowest detectable limit after 24 h.29 Bogdanova et al., 2017,30 reported that the prenylated chalcones and acylphloroglucinols; humulone 25, lupulone 26, and xanthohumol 27, isolated from Humulus lupulus L., possessed antimicrobial properties against Staphylococcus spp., including methicillin-susceptible and resistant strains, in both planktonic and biofilm-dwelling, with no significant difference between resistant and susceptible strains. Where humulone 25, lupulone 26, and xanthohumol 27 lowered the number of bacterial cells released from the biofilm, with the strongest effect seen for lupulone 26, followed by xanthohumol 27.
Moreover, lupulone 26, and xanthohumol 27 were not only able to penetrate the biofilm and reduce the number of bacteria within it, but their higher concentrations (∼60 μg mL−1 for xanthohumol 27 and ∼125 μg mL−1 for lupulone 26) reduced the number of surviving bacterial cells to zero. Asides, humulone 25, lupulone 26, and xanthohumol 27, with desmethylxanthohumol 28, cohumulone 29, colupulone 30, reported as potent antibacterial compounds with MIC < 1 μg mL−1, against MRSA strains, through kill curves, post-antibiotic effects, anti-biofilm assays and synergy studies with antibiotics.31 Alkaloids 31–34 are β-carboline type, where berberine 31 was reported to have moderate inhibitory activity against MRSA with MIC 125 μg mL−1.32 Notable efflux inhibitory activity (ranging from two-to eightfold Ethidium Bromide MIC reduction) meanwhile was detected from quinine 32, piperine 33, and harmaline 34 using reserpine 35 as the positive control.33 Canthin-6-one 36, and 8-hydroxy-canthin-6-one 37 isolated from A. neapolitanum, displayed MICs in the range 8–64 μg mL−1 against MDR/MRSA strains.34 Three carbazole alkaloids, Clausamine A, B, F 38–40, isolated from Clausena harmandiana, where clausamine B 39 exhibited significant activity against MRSA SK1 with an MIC value of 0.25 μg mL−1 which was higher than that of standard drug, vancomycin (MIC 1 μg mL−1). While clausamine F 40, and A 38 showed strong activity with MIC 4 and 8 μg mL−1, respectively. Also, clausamine F 40 showed strong antibacterial activity against S. aureus TISTR 1466 with MIC 4 μg mL−1.35
The carbazole alkaloids, 2,7-dihydroxy-3-formyl1-(3′-methyl-2′-butenyl) carbazole 41, clausenawalline E 42, clausenawalline B 43, were isolated from Clausena wallichii, and exhibited significant activity against MRSA SK1 and S. aureus TISTR 1466 with MIC 4−16 μg mL−1.36 Clausenawalline E–K 44–48 isolated also from Clausena wallichii, showed weak antibacterial activities with MIC 64–128 μg mL−1 against S. aureus TISTR 1466 and MRSA SK1, and E. coli TISTR 780 and S. typhimurium TISTR 292.36 The antibacterial activity of lysergol 49 and its synergy with the conventional antibiotic nalidixic acid (NA) against nalidixic acid-sensitive (NASEC) and nalidixic acid-resistant (NAREC) strains of Escherichia coli were evaluated. Lysergol 49 did not possess antibacterial activity of their own, but in combination, it significantly reduced the MIC of NA. Furthermore, lysergol 49 brought down eightfold reductions in the MIC of tetracycline (TET) against MDR clinical isolate of E. coli. Additionally, lysergol 49 inhibited ATP-dependent efflux pumps, which was evident by ATPase inhibition and down-regulation of multidrug ABC transporter ATP-binding protein (yojI) gene.37 Chanoclavine 50 isolated from Ipomoea muricata, showed synergy potential against multidrug-resistant Escherichia coli (MDREC). Although chanoclavine 50 did not show antibacterial activity of its own, but in combination, it could reduce MIC of tetracycline (TET) up to 16-folds. Chanoclavine 50 was found to inhibit the efflux pumps which seem to be ATPase-dependent.38 The EPI properties of indirubin 51 isolated from Wrightia tinctoria, were investigated using S. aureus SA1199B, and its synergistic effects were tested with ciprofloxacin. Indirubin 51 showed activity against multidrug-resistant Staphylococcus aureus (MDRSA) with MIC 12.5 mg L−1 for S. aureus and 25 mg L−1 for S. epidermidis. It synergistically potentiated the activity of ciprofloxacin with a fractional inhibitory concentration index (FICI) of 0.45, may be through inhibiting the NorA efflux pump. Indirubin 51 showed to exhibit EPI activity nearly comparable to that of reserpine 35 by 4-fold reduction in ciprofloxacin MIC.39 The antimicrobial DNA-intercalating alkaloid sanguinarine 52, demonstrated a strong activity against MDR-Gram-positive and Gram-negative bacteria, with MIC 0.5–128 μg mL−1.40 6-Methoxydihydrosanguinarine (6 MS) 53, 6-acetonylhydrosanguinarine 54, and dihydrosanguinarine 55 isolated from Hylomecon hylomeconoides, showed MIC against MRSA 1.95–250 μg mL−1. Where 6 MS 53 appeared to be the most active with MICs in the range of 1.9 to 3.9 μg mL−1.41 Alkaloids bis-6-(5,6-dihydro-chelerythrinyl)-ether 56, 6-ethoxy-chelerythrine 57, 4-methoxy-N-methyl-2-quinolone 58, isolated from Zanthoxylum monophyllum exhibited strong activity against MRSA (ATCC 43300). The Compound 58 exhibited significant activity against MRSA with IC50 value of 8.0 μM.42
Furthermore, dihydrochelerythrine 59, and N-methylcanadine 60, isolated from the Zanthoxylum tingoassuiba, showed potent anti-MRSA ATCC 25923 with MIC values ranging from 85.8 to 171.7 μM and 76.9 to 307.8 μM, respectively. Nevertheless, dihydrochelerythrine 59 displayed better activity than chloramphenicol against S. aureus ATCC 25923.43 It is interesting that the 8-hydroxylated benzo[c]phenanthridine derived alkaloids, 6-hydroxy-dihydrosanguinarine 61, and 6-hydroxy-dihydrochelerythrine 62, showed potent in vitro inhibitory effects on both the methicillin sensibler Staphylococcus aureus (MSSA) and MRSA strains. The 61 and 62 minimal inhibitory concentrations/minimal bactericidal concentrations (MICs/MBCs) values against MRSA strains were as low as to be 0.49/1.95 and 0.98/7.81 μg mL−1, respectively, showing that the alkaloid 61 was demonstrated as the most potent. Its 90% MICs (1.95 μg mL−1) against MRSA were comparable to vancomycin (2.34 μg mL−1).44 The antibacterial activity of two bisbenzylisoquinoline alkaloids, tetrandrine 63 and demethyltetrandrine 64 isolated from Stephania tetrandra roots, alone and in combination with the antibiotics ampicillin (AMP), azithromycin (AZM), cefazolin (CFZ) and levofloxacin (LEV) against 10 clinical isolates of staphylococcal chromosomal cassette mec (SCCmec) III type MRSA was studied. The MICs/MBCs ranges alone were 64–128/256–1,024 μg mL−1, for both compounds. Significant synergies against 90% of the isolates were observed for the tetrandrine 63/CFZ combination, with their MICs being reduced by 75–94% FICIs ranged from 0.188 to 0.625, respectively.45 Roemerine 65, is an aporphine alkaloid isolated from Annona senegalensi, and is reported to be effective in vitro against MDR strains, as it was found to increase cell membrane permeability in a concentration-dependent manner.46 Evocarpine 66 isolated from Fructus Euodiae showed activity against MRSA with MIC 8 μg mL−1, which was equivalent to or lower than the control antibiotics, oxacillin, erythromycin, and tetracycline (MIC ≥ 128 μg mL−1).47 The anti-MRSA activity of sophoraflavanone G (SFG) 67 and synergism between SFG 67 and antibacterial agents against MRSA were evaluated. The MICs of SFG 67 against 27 strains of MRSA ranged from 3.13 to 6.25 mg mL−1. Synergism between SFG 67 and vancomycin hydrochloride (VCM) or fosfomycin (FOM) was observed (FIC indices were 0.16 and 0.48), while partial synergism was admitted between SFG 67 and other antibacterial agents such as methicillin (DMPPC), cefzonam (CZON), gentamicin (GM), minocycline (MINO) and levofloxacin (LVFX) (the FIC indices were 0.71, 0.73, 0.69, 0.65 and 0.58, respectively).48 Plumbagin 68 isolated from Plumbago zeylanica showed activity against MRSA with MIC range of 4–8 μg mL−1. Where the time-kill study revealed 99% kill of a reference MRSA strain, 8 h after exposure to plumbagin 68. In the combination MIC study using the reference MRSA strain, plumbagin 68 showed synergistic effect with ciprofloxacin and piperacillin while additive or indifference effect with other commonly used antibiotics. The transmission electron micrograph of the reference MRSA strain treated with plumbagin 68 confirmed cell wall and cytoplasmic changes.49
Asides, myricetin 69, datiscetin 70, kaempferol 71, and quercetin 72, flavone 73, and luteolin 74 exhibited inhibitory activity against MRSA. Myricetin 69 was also found to inhibit the growth of MDR Burkholderia cepacia, vancomycin-resistant enterococci (VRE) and other medically important organisms such as Klebsiella pneumoniae and Staphylococcus epidermidis. Moreover, myricetin 69 was bactericidal to B. cepacian.50 Five quinonoids, emodin 75, diospyrin 76, plumbagin 77, menadione 78, and thymoquinone 79 were evaluated against a broad panel of multi-drug and extensively drug resistant tuberculosis (M/XDR-TB) strains, rapid growing Mycobacteria, and other bacterial isolates, some of which were producers of β-lactamase, Extended-spectrum β-lactamase (ESBL), AmpC β-lactamase, metallo-betalactamase (MBL) enzymes, as well as their drug-sensitive ATCC counterparts. All the tested quinones exhibited antimycobacterial and broad-spectrum antibacterial activity, particularly against M. tuberculosis (lowest MIC 0.25 μg mL−1) and Gram-positive bacteria (lowest MIC < 4 μg mL−1) of clinical origin. Where the order of antitubercular activity of the tested quinonoids was plumbagin 77 > emodin 75 ∼ menadione 78 ∼ thymoquinone 79 > diospyrin 76, whereas their antibacterial efficacy was plumbagin 77 > menadione 78 ∼ thymoquinone 79 > diospyrin 76 > emodin 75.51 Penicillin-resistant (PRSA) and MRSA were reported to be susceptible to hyperforin 80, isolated from Hypericum perforatum.52 The prenylated xanthones isolated from Calophyllum species, calozeyloxanthone 81, and 6-deoxy-γ-mangostin 82 showed inhibition against S. aureus. However, the activity of 6-deoxy-γ-mangostin 82 was not significant. The MIC of calozeyloxanthone 81 for S. aureus (MSSA and MRSA) ranged from 4.1 to 8.1 mg mL−1.53 Three acridone alkaloids; hydroxy-1, 3-dimethoxy-10-methyl-9-acridone 83, 1-hydroxy-3-methoxy-10-methyl-9-acridone 84, and 3-hydroxy-1, 5, 6-trimethoxy-9-acridone 85, isolated from Z. leprieurii stem bark, were tested against pan sensitive (H37rv), isoniazid resistant (TMC 301) and rifampicin resistant (TMC 331) strains of M. tuberculosis using micro plate alamar blue assay. The MIC of 3-hydroxy-1, 5, 6-trimethoxy-9-acridone 85 was found to be 5.1, 4.5 and 3.9 μg mL−1 on H37rv, TMC 331 and TMC 301 while that hydroxy-1, 3-dimethoxy-10-methyl-9-acridone 83 was found to be 1.5, 8.3 and 3.5 μg mL−1 respectively.54
Additionally, the phenanthrene derivatives, i.e., dehydroeffusol 86, and juncusol 87, were isolated from the common rush, Juncus effusus L., reported to enhance the antimicrobial activities in light. The MIC for these compounds against methicillin-resistant and -sensitive Staphylococcus aureus was increased up to 16-and two-fold, respectively, by irradiation with ultraviolet A (UVA). Under UVA irradiation, dehydroeffusol 86 strongly inhibited all the restriction enzymes (KpnI, XbaI, PmeI, DraI, PacI and BciVI) that have at least one 5′-TpA sequence in their recognition sites. Weak inhibitions were found for the restriction enzymes EcoRI, SacI, BamHI, SalI, PstI and HindIII, which do not possess a 5′-TpA sequence at their restriction sites and the restriction site sequences of which consist of all bases, A, T, G and C. Weak or no inhibition was found for AscI and SmaI, the restriction site sequences of which are composed of only C and G. These results indicated the necessity of thymine (adenine) for the photosensitized DNA-binding activity of dehydroeffusol 86. A strong inhibition against SphI, which does not have a 5′-TpA sequence in the restriction sequence, indicates that there are possibly other binding sequence(s) for dehydroeffusol 86. With juncusol 87 and UVA, strong inhibitions for KpnI and BciVI and trace inhibitions for PacI, XbaI, PmeI and DraI were found. This result also showed a preference of juncusol 87 for 5′-TpA, but the preference could be more selective than that of dehydroeffusol 86 depending on the surrounding sequences of 5′-TpA in the respective restriction sites. A strong inhibition of SphI by juncusol 87 with UVA also indicated the existence of an unknown binding sequence for this compound. Generally, the DNA-binding activity of juncusol 87 was weaker than that of dehydroeffusol 86.55 Growth of two strains of MRSA was inhibited by 6.25 μg mL−1 of anacardic acid 88 isolated from the cashew Anacardium occidentale, apple, nut, and nutshell oil, and 0.78 μg mL−1 of totarol 89 isolated from the bark of Podocarpus nagi, and these two compounds were found to be bactericidal. Anacardic acid 88 was founded to be bactericidal against MRSA at any stage of growth.56 Gallic acid 90, and methyl gallate 91, isolated from Terminalia chebula, exhibited inhibitory activity against MRSA with MIC 7.9–125 μg mL−1.57 A highly potent anti-MRSA sesquiterpenoid mansonone F 92 has been isolated from Ulmus davidiana var., and showed an MIC range of 0.39–3.13 mg mL−1, compared to that of vancomycin.58 Coleon U 93, 7α-acetoxy-6βhydroxyroyleanone 94, and horminone 95, are abietanes natural products isolated from Plectranthus grandidentatus and showed MIC values ranging 0.98–15.63 mg mL−1 for MRSA, and 15.63–31.25 mg mL−1 for vancomycin-resistant Enterococcus faecalis (VRE).59
Also, one active product, α-mangostin 96, a xanthone derivative isolated from Garcinia mangostana, had MIC of 1.57–12.5 μg mL−1 against MRSA. Other related xanthones was rubraxanthone 97, which was isolated from Garcinia dioica, had the highest activity against Staphylococcal strains (MIC = 0.31–1.25 μg mL−1), an activity which was greater than that of the antibiotic vancomycin (3.13–6.25 μg mL−1). The anti-MRSA activity of α-mangostin 96 was clearly increased by the presence of vancomycin; this behavior was not observed for rubraxanthone 97.60 Xanthatin 98 a sesquiterpene lactone isolated from Xanthium sibiricum, is highly species-specific for MRSA and MSSA strains.61 Alopecurone A-C 99–101, flavanostilbenes isolated from Sophora alopecuroades, inhibited MRSA strains at concentrations of 3.13–6.25 μg mL−1.62 Oleanolic acid 102, ursolic acid 103, lupeol 104, betulinic acid 105, β-sitosterol glucoside 106, and stigmasterol 107, isolated from Psychotria sycophylla, showed MICs varied from 16 to 256 μg mL−1 against Providencia stuartii PS2636, S. aureus MRSA9, S. aureus MRSA3, and Enterobacter aerogenes EA27. The mechanistic investigations showed interference of 102–107 with bacterial growth kinetic (by extending the lag phase) and inhibition of proton pumps.63 Sesquiterpene lactones, 6-O-methylacrylylplenolin 108, 6-O-isobutyroylplenolin 109, and 6-O-angeloylplenolin 110, isolated from Centipeda minima, had activity against resistance Bacillus subtilis and S. aureus, where 6-O-isobutyroylplenolin 109 being the most active with MIC 300–600 μg mL−1 for MRSA.64
Other naturally plant-derived products were shown in Fig. 7 illustrating their chemical structures, 111–190. Guaianolide 111, secoguaianolide sesquiterpene, isolated from Artemisia gilvescens showed good MRSA inhibition activity with MIC 1.95 μg mL−1.65 Dehydroleucodine 112, sesquiterpene lactone isolated from Gynoxys verrucosa, showed moderate MRSA inhibition activity with MIC50 between 49–195 μg mL−1.66 8(17),12E,14-labdatrien-6,19-olide 113, labdane diterpenoid isolated from Salvia leriifolia showed an MIC 213 μM against MRSA.67 8(17),11(Z),13(E)-trien-15,19-dioic acid 114, is epimeric cassane-type diterpenoid, isolated from Caesalpinia decapetala displayed moderate MRSA inhibiton activity with an MIC 5.99 μg mL−1.68 (E)-8(17),12-labdadiene-15,16-dial 115, zerumbol 116, are terpenes isolated from Zingiber montanum showed MIC values 32–128 μg mL−1; 0.145–0.291 mM against MDR and MRSA different strains.69 16α-hydroxycleroda-3, 13(14)-Z-dien-15, 16-olide (CD) 117, clerodane diterpene isolated from Polyalthia longifolia, exhibited significant anti-MRSA activity (15.625–31.25 mg L−1), while time kill assays at graded MICs resulted in 2.78–9.59- and 2.9–6.18-fold reduction in growth of reference strain and clinical isolates of S. aureus, respectively. The molecule CD 117 was found to interact synergistically with clinically used antibiotics (FICI ≤ 0.5) against all clinical isolates. In infected mice, CD 117 significantly (p < 0.001) lowered the systemic microbial load in blood, liver, kidney, lung and spleen tissues and did not exhibit any significant toxicity at 100 mg kg−1 body weight.70 Rel-15,16-epoxy-7α-hydroxypimar-8,14-ene 118, a diterpenoid isolated from Plectranthus ernstii exhibited moderate anti-MRSA activity with MIC of 32 μg mL−1.71 The diterpene isopimaric acid 119, isolated from Pinus nigra exhibited anti-staphylococcal activity against a range of MDR and MRSA strains of S. aureus with MIC of 32–64 μg mL−1.72 ent-kaurenoic acid 120, and ent-pimaradienoic acid 121, diterpenoid isolated from V. arenaria exhibited activity against MDR and MRSA strains of S. aureus.73 18β-glycyrrhetinic acid 122, isolated from Glycyrrhiza glabra, enhanced the bactericidal activity of the aminoglycoside's tobramycin, gentamicin, amikacin, and of polymyxin B against two MRSA strains, reducing the MIC of these antibiotics 32–64-fold with FICI of 0.12–0.13. In an air-exposed airway epithelial cell culture, 18β-glycyrrhetinic acid 122 enhanced the bactericidal activity of tobramycin and polymyxin B against the MRSA strain. Consequently, the potential of 18β-glycyrrhetinic acid 122 to synergise with certain types of antibiotics to eliminate strains of MRSA.74
 | ||
| Fig. 7 Plant derived natural products 111–190. | ||
16R-hydroxymollic 123, 15R-hydroxymollic 124, and 7α,16αdihydroxy-1,23-dideoxyjessic acid 125, were isolated from Acalypha communis, showed better antimicrobial activity against vancomycin-resistant enterococci than penicillin G (MIC 128 μg mL−1). In addition, 16R-hydroxymollic 123 was also found active against MRSA, with a MIC 64 μg mL−1 15R-hydroxymollic 124 and penicillin G were found to be equipotent against MRSA (MIC of 128 μg mL−1).75 1′-Acetoxychavicol 126, isolated from Alpinia galanga exhibited anti-plasmid activity against Salmonella typhi, Escherichia coli and vancomycin resistant Enterococcus faecalis with an efficiency of 92%, 82% and 8% respectively at 400 μg mL−1 SIC. 1′-Acetoxychavicol 126 demonstrated the ability to cure plasmid encoded antibiotic resistance in various MDR bacterial strains of clinical isolates such as Enterococcus faecalis, Salmonella typhi, Pseudomonas aeruginosa, Escherichia coli and Bacillus cereus with curing efficiency of 66%, 75%, 70%, 32% and 6% respectively at SIC of 400–800 μg mL−1.76
In addition, isovalerylshikonin (IVS) 127, was isolated from Arnebia euchroma, exhibited marginal antibacterial activity against MRSA RN4220, with MIC 16 μg mL−1. In addition, a synergistic effect between IVS 127 and streptomycin (STM) was detected by the microdilution antimicrobial chequerboard assay, with MIC reduction for STM up to 16-fold against strain RN4220. IVS 127 also significantly inhibited bacterial efflux and expression of msrA mRNA in vitro. A murine peritonitis/sepsis model was employed to test the in vivo synergistic activity of IVS 127 and STM. IVS 127 synergistically decreased bacterial counts with STM in peritoneal, spleen and liver tissue and increased mouse survival with STM in 7 days. The acute toxicity of IVS 127 was tested and the 50% lethal dose (LD50) of IVS 127 with a single exposure was 2.584 g kg−1 in mice. Overall, IVS 127, a low-toxicity RMA, exhibited synergistic antibacterial activities in vitro and in vivo against MRSA. The effects were mediated by suppression of msrA mRNA expression and reduced bacterial efflux. In addition, these data support that IVS 127 is a potential resistance-modifying agent (RMA) against microbial resistance caused by the MsrA efflux pump.77 Glycyrrhizic acid 128, at the subinhibitory concentration of 2.4 mM was found to reduce the MIC of gentamicin in intrinsically resistant E. faecium strains down to 6.25% of MIC of gentamicin alone, whereas relatively low concentrations of glycyrrhizic acid 128 (18 μM) resulted in increased susceptibilities for some E. faecium isolates to gentamicin.78 3-geranyl-1-(2-methylpropanoyl)-phloroglucinol 129; 3-geranyl-1-(2-methylbutanoyl) phloroglucinol 130; 2-geranyloxy-1-(2-methylpropanoyl)-phloroglucinol 131; 2-geranyloxy-1-(2 methylbutanoyl)-phloroglucinol 132; 2-geranyloxy-4,6-dihydroxybenzophenone 133, isolated from Hypericum densiflorum, H. ellipticum, H. prolificum, and H. punctatum, were tested for their ability to attenuate biofilm production by S. species. The MBIC values of the Hypericum metabolites ranged from 1.95–7.81 μg mL−1 3-Geranyl-1-(2-methylbutanoyl)-phloroglucinol 130, displayed the most potent biofilm inhibition against S. aureus and S. epidermidis at an MBIC of 1.95 μg mL−1. Compounds 129–131 also inhibited biofilm formation at concentrations below their respective MIC and MBC values against some test strains. Compounds 129–131 consistently demonstrated MBIC values at or below their respective MIC values.79 Corilagin 134, and tellimagrandin I 135, are polyphenols isolated from Arctostaphylos uvaursi and Rosa canina, respectively, which reported to reduce MIC of β-lactams in MRSA. Another study investigated the effect of 134–135 on the penicillin binding protein 2′ (2a) (PBP2′ (PBP2a)) which mainly confers the resistance to β-lactam antibiotics in MRSA. These compounds when added to the culture medium were found to decrease production of the PBP2′ (PBP2a) slightly. Using Bocillin Fl, a fluorescent-labeled benzyl penicillin, it was found that PBP2′ (PBP2a) in MRSA cells that were grown in medium containing corilagin 134 or tellimagrandin I 135 almost completely lost the ability to bind Bocillin Fl. The binding activity of PBP2 and PBP3 were also reduced to some extent by these compounds. These results suggested that inactivation of PBPs, especially of PBP2′ (PBP2a), by corilagin 134 or tellimagrandin I 135 are the major reason for the remarkable reduction in the resistance level of β-lactams in MRSA.14
Silybin 136, is a flavonolignan isolated from milk thistle seed, and showed to disrupt the MRSA41577 resistance to ciprofloxacin through reducing the expression of the quinolone resistance protein NorA (norA) and quaternary ammonium resistance proteins A/B (qacA/B) efflux genes in MRSA.80 Chelerythrine 137, isolated from Toddalia asiatica showed strong antibacterial activities against MRSA, and extended spectrum β-lactamase S. aureus (ESBLs-SA) with MIC 0.156 mg mL−1, which attributed to 137 destruction of the channels across the bacterial cell membranes, causing protein leakage to the outside of the cell, and to its inhibition on protein biosynthesis.12 In S. aureus, von Willebrand factor-binding protein (vWbp) is one of the key virulence determinants because it mediates not only the activation of thrombin to convert fibrinogen to fibrin, thereby enabling S. aureus to escape from the host immune clearance, but also the adhesion of S. aureus to host cells. Thus, vWbp is regarded as a promising druggable target to treat S. aureus-associated infections. Baicalein 138, isolated from Scutellaria baicalensis, can effectively block the coagulase activity of vWbp without inhibiting the growth of the bacteria. Molecular dynamics simulations and mutagenesis assays revealed that the Asp-75 and Lys-80 residues are necessary for baicalein 138 binding to vWbp. Importantly, baicalein 138 treatment attenuates the virulence of S. aureus and protects mice from S. aureus-induced lethal pneumonia. In addition, baicalein 138 can improve the therapeutic effect of penicillin G by 75% in vivo.81 Moreover, baicalein 138, at 16 μg mL−1 could synergistically restore the antibacterial actions of ciprofloxacin against the NorA efflux pump overexpressed SA-1199B, but not with the poor NorA substrate, pefloxacin. In addition, synergistic effects were observed when baicalein 138 was combined with ciprofloxacin against 12 out of 20 clinical ciprofloxacin resistant strains. For MRSA PK studies, baicalein 138 alone could inhibit the enzymatic activity of MRSA PK in a dose-dependent manner.82 Abietane diterpenoid salvipisone 139, demonstrated a very interesting activity when its effect on 24 h-old staphylococcal biofilm cells viability was examined. It limited the survival of biofilms formed by S. aureus as well as by S. epidermidis, putting this compound to the list of potential anti-biofilm agents, better than most of known antibiotics.83 The pentacyclic triterpenoids were isolated from Callicarpa farinosa: α-amyrin 140, and betulinaldehyde 141, exhibited antimicrobial activities against MRSA and MSSA, with MIC ranging from 2 to 512 μg mL−1. From the genome-wide transcriptomic analysis to elucidate the antimicrobial effects of these compounds, multiple novel cellular targets in cell division, two-component system, ABC transporters, fatty acid biosynthesis, peptidoglycan biosynthesis, aminoacyl-tRNA synthetases, ribosomes and β-lactam resistance pathways are affected, resulting in destabilization of the bacterial cell membrane, halt in protein synthesis, and inhibition of cell growth that eventually led to cell death.84
Dehydroabietic acid (DA) 142, isolated from Pinus elliottii, showed the MIC and minimum bactericidal concentration varied between 6.25 and 50, and between 6.25 and 100 μg mL−1, respectively, against MRSA. The time-kill assay conducted with DA 142 at 6.25 μg mL−1 evidenced bactericidal activity against S. epidermidis 14990 within 24 h.85 (+)-Lyoniresinol-3α-O-β-D-glucopyranoside 143, isolated from Lycium chinense, exhibited potent anti-MRSA activity with MIC 2.5–5 14 μg mL−1.86 7,9,2′,4′-Tetrahydroxy-8-isopentenyl-5-methoxychalcone (THIPMC) 144, isolated from Sophora flavescens, was found to be active against MRSA and VRE, either alone or in combination with ampicillin (AM) or gentamicin (GM). The MIC 1–8 μg mL−1 for THIPMC 144, from 128–1024 μg mL−1 for AM, and from 128–512 μg mL−1 for GM, respectively. The combinations of THIPMC 144 plus AM or GM yielded FICI ranging from 0.188 to 0.375 μg mL−1, thereby indicating a synergistic effect.87 20-Hydroxyecdysone (20E) 145, isolated from Achyranthes japonica, was found to be active MRSA, either alone or in combination with ampicillin (AM) or gentamicin (GM). These results investigated the antibacterial activity of 20E 145, which exhibited poor antibacterial activity (MIC = 250–500 μg ML−1) against all the bacterial strains tested. But the combined activity of ampicillin (AM), gentamicin (GE) plus 20E 145 against MRSA resulted in FICs ranging from 4.00 to 0.031 μg mL−1, respectively. Meanwhile, the FIC index ranged from 0.16–4.50, indicating a marked synergistic relationship between AM, GE and 20E 145 against MRSA with enterotoxin gene in vitro.88 The seeds of Swietenia mahagoni afforded two limonoids, swietenolide 146, and 2-hydroxy-3-O-tigloylswietenolide 147, showed MDR against haemolytic S. aureus, S. aureus, S. pneumoniae, Haemophilus influenzae, E. coli, Klebsiella pneumoniae, Salmonella typhi, and Salmonella paratyphi. compound 147 displayed overall more potent activity than compound 146.89 Ellagic acid 148 from Rosa rugosa; norwogonin 149 from Scutellaria baicalensis; and chebulagic acid 150, chebulinic acid 151, corilagin 152, and terchebulin 153 from Terminalia chebula, had MDR activities. The most potent compound was norwogonin 149 with MIC 128 μg mL−1, and MBC 256 μg mL−1 against clinically relevant strains of A. baumannii.90 3β-O-p-coumaroyltormentic acid 154, isolated from Planchonia careya, showed weakly selective for VRE compared with eukaryotic cells, with MIC 59.4 μg mL−1 and IC50 of 72.0 μg mL−1 for MA104 cells.91 Ent-18-acetoxy-11α-hydroxykaur-16-en-15-one 155, ent-18-acetoxy-7β-hydroxykaur-16-en-15-one 156, ent-18-acetoxy-7β,14α-dihydroxykaur-16-en-15-one 157, isolated from Croton tonkinensis, exhibited MICs 32, 500, and 125 μg mL−1, respectively, against MRSA strains.92 9-Methoxy-tariacuripyrone 158; and aristololactam I 159; isolated from Aristolochia brevipes, demonstrated very good anti-tuberculous activity against sensitive, mono-resistant, and clinically strains, MDR, with MIC 25–50 μg mL−1, except for M. tuberculosis H37RvIr, for MIC 12.5 μg mL−1 for 158. Aristololactam I 159 demonstrated the greatest inhibitory activity against all strains assayed, with MIC 12.5–25.0 μg mL−1.93 Tiliacorinine 160, 2′-nortiliacorinine 161, and tiliacorine 162, isolated from Tiliacora triandra, were tested against 59 clinical isolates of MDR M. tuberculosis (MDR-MTB). The alkaloids 160–162 showed MIC 0.7–6.2 μg mL−1, but they exhibited the MIC 3.1 μg mL−1 against most MDR-MTB isolates.94
(−)-Licarin A (LA) 163, was isolated from Aristolochia taliscana and the antitubercular activity of LA 163 was tested in a TB murine model inducing disease with M. tuberculosis H37Rv or MDR. In animals infected with drug sensitive or MDR strains, LA 163 produced a significant decrease of pulmonary bacillary burdens at day 30 of treatment, and a significant pneumonia reduction at days 30 and 60 of treatment.95 Maritinone 164, and 3,3′-biplumbagin 165, showed the strongest activity against both MTB/H37Rv strains (MIC 1.56–3.33 μg mL−1). The bioactivity of maritinone 164 and 3,3′-biplumbagin 165 were 32 times more potent than rifampicin against the pan-resistant strain, and both dimers showed to be non-toxic against PBMC and Vero cells, with selectivity index (SI) of maritinone 164 and 3,3′-biplumbagin 165 on Vero cells was 74.34 and 194.11 against sensitive and pan-resistant MTB strains, respectively.96 Ent-18-hydroxykaur-16-en-15-one 166, ent-18-acetoxy-7α-hydroxykaur-16-en-15-one 167, ent-1β,14β-diacetoxy-7α-hydroxykaur-16-en-15-one 168, ent-1β,7α-diacetoxy-14β-hydroxykaur-16-en-15-one 169, ent-1β,7α,14β-triacetoxykaur-16-en-15-one 170, ent-1β-acetoxy-7α,14 β-dihydroxykaur-16-en-15-one 171, ent-7α,14β-dihydroxykaur-16-en-15-one 172, ent-7α,18-dihydroxykaur-16-en-15-one 173, ent-18-acetoxy-7α,14β-dihydroxykaur-16-en-15-one 174, ent-18-acetoxy-11β-hydroxykaur-16-en-15-one 175, ent-11β-acetoxy-7α-hydroxykaur-16-en-15-one 176, ent-11β-acetoxykaur-16-en-18-ol 177, ent-11β-acetoxykaur-16-en-18-oic acid 178, ent-18-acetoxy-7α-hydroxykaur-16-ene 179, ent-18-acetoxy-11α-hydroxykaur-16-ene 180, ent-16(S)-18-acetoxy-7α-hydroxykaur-15-one 181, 14α-hydroxykaur-16-en-7-one 182, 7α,10α-epoxy-14β-hydroxygrayanane-1(5),16(17)-dien-2,15-dione 183, and 7α,10α-epoxy-14β-hydroxygrayanane-1(2),16(17)-dien-15-one 184, are diterpenoids isolated from Croton tonkinensis. All diterpenoids showed high to moderate activity against Mycobacterium. The highest antituberculosis activity was observed for ent-1β,7α,14β-triacetoxykaur-16-en-15-one 170, with MIC 0.78,1.56 and 3.12–12.5 μg mL−1 against H37Ra, H37Rv and all other resistant strains of M. tuberculosis examined.97 Ethyl p-methoxycinnamate (EPMC) 185, isolated from Kaempferia galanga, was shown to inhibit M. tuberculosis H37Ra, H37Rv, drug susceptible and MDR clinical isolates (MIC 0.242–0.485 mM).98 Plumericin 186, showed better activity against pan sensitive as well as four MDR strains of M. tuberculosis with MIC values of 2.1 ± 0.12, 1.3 ± 0.15, 2.0 ± 0.07, 1.5 ± 0.13, and 2.0 ± 0.14 μg mL−1 and MBC 3.6 ± 0.22, 2.5 ± 0.18, 3.8 ± 0.27, 2.9 ± 0.20, and 3.7 ± 0.32 μg mL−1 than isoplumericin 187, respectively, isolated from Plumeria bicolor. Interestingly, both compounds showed an advantage over rifampicin (80 times) and isoniazid (8 times) by being highly active against the MDR strains.99
Conessine 188, isolated from Holarrhena antidysenterica, combined with various antibiotics for synergistic activity determination against resistance P. aeruginosa PAO1 strain K767 (wild-type), K1455 (MexAB-OprM overexpressed), and K1523 (MexB deletion). H33342 accumulation assay was used to evaluate efflux pump inhibition while NPN uptake assay was assessed membrane permeabilization. Conessine 188 significantly reduced MICs of all antibiotics by at least 8-fold in MexAB-OprM overexpressed strain. With erythromycin, novobiocin, and rifampicin, MICs were 4-8-fold < MICs of the wild-type strain. Loss of MexAB-OprM due to deletion of mexB affected susceptibility to almost all antibiotics, except novobiocin. Synergistic activities between other antibiotics (except novobiocin) and conessine 188 observed in MexB deletion strain suggested that conessine 188 might inhibit other efflux systems present in P. aeruginosa. Inhibition of H33342 efflux in the tested strains clearly demonstrated that conessine 188 inhibited MexAB-OprM pump. In contrast, the mode of action as a membrane permeabilizer was not observed after treatment with conessine 188 as evidenced by no accumulation of 1-N-phenylnaphthylamine. These results suggested that conessine 188 could be applied as a novel efflux pump inhibitor to restore antibiotic activity by inhibiting efflux pump systems in P. aeruginosa. The findings speculated that conessine 188 may also have a potential to be active against homologous resistance–nodulation–division (RND) family in other Gram-negative pathogens.100 Tomatidine (TO) 189, a steroidal alkaloid from solanaceous plants, possesses potent antibacterial activity against S. aureus small-colony variants (SCVs). Using genomic analysis of in vitro-generated TO-resistant S. aureus strains to identify mutations in genes involved in resistance, identified the bacterial ATP synthase as the cellular target.101
The growth of the majority of Pseudomonas, Streptococcus, and Staphylococcus isolates was completely inhibited by 64 μg mL−1 allicin 190. S. pyogenes SNo 67467, S. pneumoniae SNo 68668, and S. aureus ATCC 43300 were completely inhibited by 32 μg mL−1 allicin 190 and all A. baumannii isolates were completely inhibited by 16 μg mL−1. K. pneumoniae isolates were slightly more resistant, with a MIC of 128 μg mL−1. P. aeruginosa DSM2659 showed high resistance to allicin 190 (MIC 512 μg mL−1) compared to P. aeruginosa PAO1 SBUG8 and PAO25 (MIC 64 μg mL−1). MDR and non-MDR S. pneumoniae strains tested were equally susceptible to allicin 190 and showed MICs 32–64 μg mL−1 allicin 190 and MBCs from 64 to 128 μg mL−1 allicin 190, respectively. In comparison to conventional antibiotics, the MICs, and MBCs for allicin 190 were generally higher. Thus, except for the MDR strains, the clinical isolates of S. pneumoniae were susceptible to all tested antibiotics at <1 μg mL−1. The MDR S. pneumoniae isolates were resistant to erythromycin and clindamycin (MICs > 256 μg mL−1) and for these MDR strains, allicin 190, including in absolute μM terms, compared favorably with those antibiotics.102
Fig. 8 illustrates the chemical structure for plants natural derived compounds 191–265. The isoflavone biochanin A 191, exhibited ethidium bromide (EtBr) efflux pump inhibiting activity in Mycobacterium smegmatis mc2155 comparable to that of verapamil. The stilbene resveratrol 192, and formononetin 193, were less active.103 4′,6′-Dihydroxy-3′,5′-dimethyl-2′-methoxychalcone 194, 3,5,4′-trimethoxy-trans-stilbene 195, isolated from Dalea versicolor, were very weakly active alone (MICs of 250 and 500 μg mL−1, respectively), but they caused complete growth inhibition at very low concentrations (∼3.3 μg mL−1) in combination with a subinhibitory concentration of berberine 31 against NorA mutant S. aureus.104 4-((E)-5-(3,3-dimethyl-2-oxiranyl)-3-methyl-2-pentenyl)-oxy-7H-furo(3,2)-chromen-7-one 196, isolated from grape fruit oil, enhanced the susceptibility of test MRSA strains to ethidium bromide and norfloxacin.105 Sophoraflavanone B (SPF-B) 197, a prenylated flavonoid, isolated from Desmodium caudatum, showed MIC against MRSA 15.6–31.25 μg mL−1. The optical density at 600 nm of MRSA suspensions treated with a combination of detergent and SPF-B 197 reduced the MRSA by 63–73%. In the SPF-B 197 and PGN combination assay, direct binding of SPF-B 197 with PGN from S. aureus was evident.106 Naringenin 198, eriodictyol 199, and taxifolin 200, are good candidate for β-Ketoacyl acyl carrier protein synthase (KAS) III inhibitors, which is a key catalyst in bacterial fatty acid biosynthesis, and showed good binding affinities, and docked well with efKAS III, with MIC 128–512 μg mL−1.107 Galbanic acid 201, a sesquiterpene coumarin isolated from Ferula szowitsiana roots, was investigated for its antimicrobial activity as well as ethidium bromide, in six MDR clinical isolates of S. aureus. Galbanic acid 201 had inhibitory effect on none of the isolated bacteria tested (up to 800 μg mL−1). The MIC range of ciprofloxacin, tetracycline, and ethidium bromide, against all tested S. aureus were 10–80, 10–80 and 4–16 μg mL−1, respectively. These were reduced to ≤2.5–5, 2.5–5 and 0.5–2 μg mL−1 in the presence of galbanic acid 201 (300 μg mL−1) or verapamil (100 μg mL−1). The rate of ethidium bromide (2 μg mL−1) accumulation in clinical isolates was enhanced with galbanic acid 201 (300 μg mL−1). There is also a decrease in loss of ethidium bromide from bacteria in the presence of galbanic acid 201, like verapamil (100 μg mL−1).108 α-Mangostin (AMG) 202, and isobavachalcone (IBC) 203, under 8 μg mL−1 dramatically restored the activity of colistin against MDR E. coli B2 isolate, with the decreased concentration of colistin from 8 to 0.0625 μg mL−1. The MIC50 and MIC90 against MRSA and VRE were 0.5 and 4–8 μg mL−1 for AMG 202 and IBC 203, respectively. Both AMG 202 and IBC 203 display similar efficiency to vancomycin. These results indicate that AMG 202 and IBC 203 are potent antibiotic candidates to combat MDR bacteria, practically against Gram-positive pathogens.109
 | ||
| Fig. 8 Plants/endophytes derived natural products 191–265. | ||
(22E,24R)-6β-methoxyergosta-7,22-diene-3β,5α-diol 204, (22E,24R)-6β-methoxyergosta-7,22-diene-3β,5α-diol 205, isolated from a pathogenic fungus, Microdochium majus strain 99049, from wheat, showed moderate to weak anti-MRSA activity (MIC 25, 100 μg mL−1, respectively).110 The antimicrobial epidithiodioxopiperazine, gliotoxin 206, and bisdethiobis (methylthio)gliotxin 207, were isolated from the endophytic fungus Hypocrea virens, from Premna serratifolia, showed MIC 32–35 μg mL−1 against MRSA.111 Terrenolide S 208, isolated from the endophytic fungus Aspergillus terreus isolated from the roots of Carthamus lanatus, displayed a potent activity towards MRSA with IC50 2.29 μM.112 (22E, 24R)5, 8-epidioxy-5α,8α-ergosta-6,22E-dien-3β-ol 209, isolated from Chinese mangrove Ceriops tagal endophytic Cytospora sp., showed weak anti-MRSA with MIC 233 μM.113 Helvolic acid 210, isolated from endophytic fungus Xylaria sp. from Anoectochilus setaceus, showed potent anti-MRSA with MIC 4 μM.114 Cyschalasin A & B 211–212, isolated from the endophytic fungus Aspergillus micronesiensis, showed anti-MRSA with MIC 17.5, and 10.6 μM, respectively.115 (22E,24R)-stigmasta-5,7,22-trien-3β-ol 213, isolated from the endophytic fungus Aspergillus terreus, derived from Carthamus lanatus, exhibited potent antibacterial activity MRSA with IC50 0.96 μg mL−1.116 Aspergillusphenol A 214, isolated from the endophytic fungus Rhytidhysteron sp. BZM-9, showed moderate antimicrobial activity to MRSA with a MIC value of 6.25 μg mL−1.117 Cyclo-(tryptophanyl-prolyl) 215, and chloramphenicol 216, isolated from Universiti Kebangsaan 25, from Zingiber spectabile, showed anti-MRSA with a MIC 16, 8 μg mL−1, respectively.118 3-Hydroxy-1-(1,3,8-trihydroxy-6-methoxynaphthalen-2-yl) propan-1-one 217; 3-hydroxy-1-(1,8-dihydroxy-3,5-dimethoxynaphthalen-2-yl) propan-1-one 218, and 3-hydroxy-1-(1,8-dihydroxy-3,6-dimethoxynaphthalen-2-yl) propan-1-one 219, were isolated from the Phomopsis fukushii, an endophyte of plant Paris polyphylla, showed anti-MRSA activity with MIC 4, 4, and 8 mg mL−1, respectively.119 Oxysporone 220, and xylitol 221, isolated from the endophytic fungus Pestalotia sp. from Heritiera fomes, were tested against various strains of MRSA, i.e., XU212, ATCC 25923, SA-1199B, EMRSA-15, MRSA340702, and showed MIC 128, 128, 64, 64, and 128 μM, respectively, for xylitol 221, and 128, 64, 32, 32, and 64 μM, respectively, for oxysporone 220.120 Guignardone I 222, and guignardone B 223, isolated from the endophytic fungus A1 of the mangrove plant Scyphiphora hydrophyllacea exhibited zone of inhibition of 9.0 mm and 8.0 mm, respectively, against MRSA at 65 μM.121 A new fatty acid glycoside, (R)-3-hydroxy undecanoic acid methylester-3-O-α-L-rhamnopyranoside 224, was isolated from the Guignardia sp. from same plant, and zone of inhibition was found to be 10.7 mm against MRSA.122 2,3-Didehydro-19α-hydroxy-14-epicochlioquinone B 225, griseophenone C 226, and tetrahydrobostrycin 227, were isolated from the endophytic Nigrospora sp. MA75 from the mangrove plant Pongamia pinnata that exhibited anti-MRSA activity with MIC values of 16.5, 1.6, and 5.9 μM, respectively.123 2-Chloro-5-methoxy-3-methylcyclohexa-2,5-diene-1,4-dione 228, isolated from the endophytic fungus Xylaria cubensis PSU-MA34, from mangrove Bruguiera parviflora exhibited anti-MRSA with MIC 128 μg mL−1.124 Xanalteric acid I 229, xanalteric acid II 230, and altenusin 231, isolated from the endophytic Alternaria sp., from mangrove Sonneratia alba, exhibited anti-MRSA with MIC values of 125, 250, and 31.25 μg mL−1, respectively.125
A bioactive compound, equisetin 232, was isolated from endophytic Fusarium sp., and exhibited anti-MRSA activity with MIC 16 μg mL−1.126 Terretonin 233, terretonin A 234, butyrolactone VI 235, aspernolide F 236, and aspernolide G 237, were isolated from the endophytic fungus Aspergillus terreus from the roots of Carthamus lanatus. All compounds exhibited anti-MRSA activity with IC50 0.94 to <20 mg mL−1.127 Primin 238, isolated from endophytic fungus Botryosphaeria mamane PSU-M76, isolated from Garcinia mangostana leaves, exhibited anti-MRSA with MIC 8 mg mL−1.128 (3S)-lasiodiplodin 239, isolated from endophytic PSU-M35 and PSU-M11 were isolated from Garcinia mangostana leaves, exhibited anti-MRSA with MIC 128 mg mL−1.129 Beauvericin 240, isolated from endophytic fungus Fusarium oxysporum, isolated from Cinnamomum kanehirae, exhibited potent anti-MRSA with MIC 3.125 μg mL−1.130 An endophytic fungus Fusarium tricinctum isolated from Aristolochia paucinervis when cocultured with B. subtilis on solid rice medium increased the secondary metabolite production by 78-fold, i.e., increase in concentration of lateropyrone 241, lipopeptide fusaristatin A 242, and three cyclic depsipeptides of the enniatin type (enniatin B 243, enniatin B1 244, enniatin A1 245). The antibacterial activity of this compounds was tested against various pathogenic microbes including MDR S. aureus (S. aureus 25697 strain) and showed MIC 2–64 μg mL−1. The highest antimicrobial activity was shown by lateropyrone 241 with MIC 2–4 μg mL−1, followed by enniatin A1 245 with MIC 4–8 μg mL−1 and enniatin B1 244 that showed activity at MIC of 8 μg mL−1.131 Apicidin 246, was isolated from the endophytic fungi Fusarium sp. from the plant Anemopsis californica. Anti-quorum sensing inhibition of apicidin 246 was tested against MRSA. Apicidin 246 showed anti-MRSA by targeting AgrA plasmid.132 3-(2-Hydroxypropyl)-benzene-1,2-diol 247, and desoxybostrycin 248, were isolated from the PSU-N24, an endophyte of Garcinia nigrolineata. At 128 μg mL−1, the two compounds exhibited anti-MRSA activity.133 Mycoleptodiscin B 249, isolated from Mycoleptodiscus sp. isolated from Calamus thwaitesii, showed anti-MRSA activity with MIC 32 μg mL−1.134
Trichosetin 250, beauvericin A 251, beauvericin 252, enniatin I 253, and enniatin H 254, isolated from endophytic fungi Fusarium sp. TP-G1 isolated from the roots of Dendrobium officinale Kimura, showed anti-MRSA with MIC 2, 2, 8, 16, and 32, respectively.135 Skyrin 255 isolated from the Talaromyces wortmannii, an endophyte of aloe vera exhibited anti-MRSA activity with MIC 4 μg mL−1.136 Piliformic acid 256, was isolated from the Xylaria cubensis BCRC 09F 0035, an endophyte of Litsea akoensis, and exhibited MIC 200 μg mL−1 against MRSA.137 Andiconin C 257, isolated from the Aspergillus sp. TJ23, an endophyte of Hypericum perforatum, exhibited anti-MRSA with MIC > 100 μg mL−1.138 Alternariol 258, isolated from the endophytic fungus Alternaria alternata resident of plant Grewia asiatica, exhibited anti-MRSA with MIC 8 μg mL−1.139 2-Deoxy-sohirnone C 259, isolated from the Penicillium sp. GD6, an endophyte of Bruguiera gymnorrhiza showed anti-MRSA with MIC 80 μg mL−1.140 Cytosporone D 260, and cytosporone E 261, were isolated from the Cytospora sp. CR200, an endophyte collected from Conocarpus erectus, and exhibited moderate anti-MRSA with MIC 8–64 μg mL−1.141 Fusaric acid 262, was isolated from the endophytic fungi Fusarium Sp. DZ-27, isolated from the mangrove plant Kandelia candel, showed Antimycobacterial activity against clinical MDR M. tuberculosis strains, and clinical extensively drug-resistant M. avium-intracellular strains with MIC 10–60 μg mL−1.142 4-Deoxybostrycin 263, and nigrosporin 264, isolated from the endophytic fungus Nigrospora sp., showed antimycobacterial activity against clinical MDR M. tuberculosis strain (K2903531), clinical MDR M. tuberculosis strains (0907961), clinical drug-resistant M. tuberculosis strain (K0903557), and clinical extensively drug-resistant (XDR) M. avium-intracellular strain (K0803182), with MIC values in the range of 5 to >60 μg mL−1 and 15 to >60 μg mL−1, respectively.143 Vermelhotin 265, was isolated from endophytic fungus MEXU 26343, collected from the plant Hintonia latiflora,144 exhibited antimycobacterial against clinical strains of MDR-TB with MIC 1.5–12.5 g mL−1.145
Fig. 9 illustrates the chemical structure for natural derived compounds 266–307. 8-O-methylepiaustdiol 266, stemphyperylenol 267, skyrin 268, secalonic acid 269, and norlichexanthone 270, isolated from endophytic fungus Talaromyces sp. ZH-154, was isolated from Kandelia candel stem bark, exhibited antimicrobial activity against MDR P. aeruginosa with MIC 25.0, 12.5, 12.5, 12.5, and 25.0 μg mL−1, respectively.146 Neosartorin 271, derived from the endophytic fungi Aspergillus fumigatiaffinis, exhibited antimicrobial activity against resistant E. faecalis with MIC 16–32 μg mL−1.147 The polyketide setosol 272, isolated from the endophytic fungi Preussia isomera, resident of the stem of Panax notoginseng, exhibited antibacterial activity with MIC 25 μg mL−1 against MDR-resistant E. faecalis, MRSA, and MDR E. faecium.148 Equisetin 273, isolated from the endophyte Chaetomium globosum XL-1198, isolated from Salvia miltiorrhiza, exhibited antibacterial activity against MDR S. epidermidis, MDR E. faecalis, MRSA, and MDR E. faecium with MIC 3.13–6.25 μg mL−1.149 2′-acetyl-4′,4-dimethoxybiphenyl-2-carbaldehyde 274, was isolated from the endophyte Pestalotiopsis zonata resident of the plant Cyrtotachys lakka, exhibited weak anti-MRSA and vancomycin-resistant E. faecium with MIC values of 0.84 and 0.87 μm mL−1 respectively.150 Alterporriol N 275, Alterporriol D 276, were isolated from endophytic Stemphylium globuliferuman isolated from Mentha pulegium and showed anti-MRSA with MICs of 62.5, 31.25 μg mL−1, respectively.151 Indolyl-3-carboxylic acid 277, 5-acyl-2-methylpyrrole 278, isolated from the endophyte S20 of Cephalotaxus hainanensis Li. showed anti-MRSA with diameters of inhibition zones 8, 10 mm/10 μg mL−1, respectively, impregnated on sterile filter paper discs (6 mm diameter).152,153
 | ||
| Fig. 9 Endophytes derived natural products 266–307. | ||
Xiamycin A 279, indosespene 280, were produced by endophytic Streptomyces sp. HKI0595, isolated from mangrove tree Kandelia candel, showed anti-MRSA and vancomycin-resistant E. faecalis.154 Violaceol I 281, and Violaceol II 282, isolated from endophytic fungus Trichoderma polyalthiae extracted from culture broth media, showed anti-MRSA, with MIC values <9.765–156.25, <9.765–312.5 μg mL−1, respectively.155 In agar diffusion assays run on bacterial lawns, guanacastepene 283, isolated from endophytic fungus CR115, Daphnopsis americana, shows anti-MRSA and vancomycin-resistant E. faecalis. Against MRSA 100 μg of guanacastepene 283 or vancomycin produce 11-, and 17 mm zones of growth inhibition, respectively. While vancomycin is ineffective against VREF, guanacastepene 283 produced a 9 mm zone of growth inhibition.156 (2R,3S)-7-ethyl-1,2,3,4-tetrahydro-2,3,8-trihydroxy-6-methoxy-3-methyl-9,10-anthracenedione 284, isolated from the mangrove-derived fungus Phomopsis sp. PSU-MA214, showed anti-MRSA-SK1.157 Nodulisporin H 285, and 8-O-methylnodulisporin F 286, isolated from the mangrove-derived fungus Daldinia eschscholtzii HJ004, showed a moderate anti-MRSA with MIC 6.25–12.5 μg mL−1.158
Differanisole A 287, 2,6-dichloro-4-propylphenol 288, and 4,5-dimethylresorcinol 289, isolated from endophytic fungus Chaetomium sp. HQ-1, exhibited moderate anti-MRSA, with MIC 16–128 μg mL−1.159 Diaporthin 290, and orthosporin 291, isolated from endophytic fungus Diaporthe terebinthifolii, showed anti-MRSA.160 Aziridine, 1-(2-aminoethyl)-292, isolated from endophytic fungus Cochliobolus sp. APS1, from Andrographis paniculate, showed MIC 15.62 to 250 μg mL−1 against MRSA and VRSA.161 ω-Hydroxyemodin 293, emodic acid 294, (+)-2′S-isorhodoptilometrin 295, isolated from endophytic fungus Penicillium restrictum was isolated from the stems of a milk thistle plant. These compounds were quorum sensing inhibitors in a clinical isolate of MRSA, with IC50 8–120 μM.162 Pyrimidine-2,4-dion 296, isolated from endophytic fungus Bacillus sp. RD26, isolated from Phyllanthus amarus, showed anti-MRSA, with MIC 64 μg mL−1.163 Stephensiolides I, D, G, C, F 297–301, isolated from endophytic fungus Lecanicillium sp. from Sandwithia guyanensis plant. Stephensiolides I 297 showed a strong anti-MRSA with MIC ≤ 4 μg mL−1, while stephensiolides G 299 exhibited activity with MIC ≤ 16 μg mL−1, D 298 and F 301 were found to be less active with MIC ≤ 32 μg mL−1, and C 299 showed a moderate MIC of ≥128 μg mL−1.164 1-Monolinolein 302, bafilomycin D 303, nonactic acid 304, daidzein 305, 3′-hydroxydaidzein 306, isolated from endophytic actinomycete strain YBQ59 was isolated from Cinnamomum cassia, exhibited anti-MRSA ATCC 33591 and methicillin-resistant Staphylococcus epidermidis ATCC 35984 (MRSE) among which 302 revealed the strongest effects with MIC 8.5 and 14.6 μg mL−1, respectively. However, 303 showed high potential effect against MRSA (MIC of 11.1 μg mL−1) but less effect against MRSE (MIC of 30.3 μg mL−1).165 Eutyscoparin G 307, isolated from endophytic fungus Eutypella scoparia SCBG-8, displayed anti-MRSA with MIC 6.3 μg mL−1.117
The novel homicorcin peptide 308, isolated from plant endophyte S.s hominis strain MBL_AB63, displayed anti-MRSA.166 Actinomycin D 309 peptide, produced by plant endophyte S. smyrnaeus UKAQ_23, showed anti-MRSA ATCC 33591 (MIC of 2.5 μg mL−1), and MDR M. tuberculosis MDRP (IC50 of 10 μg mL−1).167 Munumbicin peptide named munumbicins C 310, produced by plant endophyte Streptomyces NRRL 30562, showed activity against drug-resistant M. tuberculosis MDR-P (IC50 > 125 μg mL−1) and vancomycin-resistant ciprofloxacin-sensitive Enterococcus faecalis ATCC 51299 (MIC of 16 μg mL−1).168 Munumbicins E-4 peptide 311, was produced by endophytic Streptomyces NRRL 30562, isolated from Kennedia nigricans, showed anti-MRSA 43000 (MIC of 16 μg mL−1).169 Kakadumycin A 312, peptide produced by Streptomyces NRRL 30566, an endophyte of the plant Grevillea pteridifolia, showed activity against MRSA ATCC 33591 (MIC of 0.5 μg mL−1).170
3.3. Lichens/endo-lichens derived natural products
Fig. 10 illustrates the chemical structure for natural derived compounds 313–341. Most reports on the activity of the following isolated compounds against MDR bacteria relate to in vitro studies. (+)-Usnic acid 313, isolated from Usnea steineri, exhibited strong activity against resistance strains of S. epidermidis (MIC 3.12 μg mL−1), S. aureus and S. haemolyticus (MIC 12.5 μg mL−1).171,172 Atranorin 314, diffractaic acid 315, sphaerophorin 316, fumarprotocetraric acid 317, psoromic acid 318, tenuiorin 319, variolaric acid 320, and vicanicin 321, isolated from lichens, collected in several Southern regions of Chile (including Antarctica), showed anti-MRSA with MIC 8–1024 μg mL−1.172,173 Thamnolic acid 322, isolated from Usnea florida showed activity against the drug-resistant M. tuberculosis H37Rv strain with MIC 250 μg mL−1.174 Salazinic acid 323 was isolated from Usnea hirta, showed anti-MRSA with MIC 7.8 μg mL−1.175 Evernic acid 324 (Evernia prunastri), hybocarpone 325 (Lecanora conizaeoides), lobaric acid 326 (Sterocaulon dactylophyllum), physodic acid 327 (H. physodes), rhizocarpic acid 328 (Psilolechia lucida), 3-hydroxyphysodic acid 329 (H. physodes), vulpinic acid 330 (Letharia vulpina), showed anti-MRSA with MIC 4–128 μg mL−1.176,177 Collatolic acid 331 from Lecanora atra, epiphorellic acid 332 from Cornicularia epiphorella, perlatolic acid 333 from Stereocaulon sp., protolichesterinic acid 334 from Cornicularia aculeata, showed anti-MRSA with MIC 4–128 μg mL−1.178 Pannarin 335 from Psoroma. dimorphum, showed anti-MRSA with MIC 4–8 μg mL−1.179 Divaricatic acid 336 was isolated from Evernia mesomorpha, showed anti-MRSA with MIC 7 μg mL−1.180 Norlichexanthone 337, isolated from Everniastrum sp., showed anti-MRSA with IC50 5.4 μg mL−1.181 Ophiobolin P& T 338–339, isolated from endo-lichen fungus Ulocladium sp. (CHMCC5507), from Everniastrum sp. lichen, showed anti-MRSA with IC50 25.1, 12.7 μg mL−1, respectively.182 Barbatic acid 340, from Cladia aggregate, showed anti-MRSA with MIC 100 μg mL−1.183 Norlichexanthone 341, isolated from endo-lichen fungus Ulocladium sp., isolated from Everniastrum sp., showed anti-MRSA with IC50 20.95 μg mL−1.181 | ||
| Fig. 10 Lichens/endo-lichens derived natural products 313–341. | ||
3.4. Insects/animal/and their associated symbiont organism derived natural products
Fig. 11 illustrates insects/animal/and their associated symbiont organism derived natural products. Most reports on the activity of the following isolated compounds against MDR bacteria relate to in vitro studies. Hexanedioic acid 342, Lauric acid 343, glycerol monolaurate 344, isolated from the edible Hermetia illucens larvae, showed anti-MRSA with MIC 137.369 μg mL−1.184,185 The prenylflavanones propolin H 345, propolin G 346, propolin D 347, and propolin C 348, isolated from Pacific propolis from bees' nest, showed anti-MRSA, where propolin D 347 and C 348 were the most active with MIC 8–16, 8–32 μg mL−1, respectively.186 Also, the propolins D 347, C 348, F 349 and G 346 from Taiwanese green propolis was obtained and showed antibacterial activity with MIC less than 2 μg mL−1 and MBC of 4 μg mL−1 against MRSA.187 | ||
| Fig. 11 Selected insects/animal/and their associated symbiont organism derived natural products. | ||
Roseoflavin 350 was isolated from Streptomyces davaonensis YH01, which was isolated from the body surface of the queen of Odontotermes formosanus, and showed potential against nine kinds of MRSA strains, with inhibition zones in the ranges of 12.7–19.7 mm under a concentration of 15 μg/6 mm discs and 18.3–22.7 mm under a concentration of 30 μg/6 mm discs.188 Actinomycin D and Actinomycin X2, isolated from endophytic Gordonia in the intestinal tract of Periplaneta americana, have anti-MRSA (ATCC 43300), with MIC 0.25 μg mL−1.189 4-Methoxy-2H-pyran-2-one 351, 4-methoxy-6-pentyl-2H-pyran-2-one 352, 6-(1-hydroxypentyl)-4-methoxy-pyran-2-one 353, 6-[8-propyloxiran-1-yl]-4-methoxypyran-2-one 354, pestalotin 355, 5,6-dihydro-4-methoxy-6-(pentanoyloxy)-2H-pyran-2-one 356, and cyclo-(L-Pro-L-Val) 357, isolated from Chrysosporium multifidum fungus isolated from Hermetia illucens gut, showed moderate anti-MRSA, where compound 354 showed the greatest activity (IC50 = 11.4 ± 0.7 μg mL−1 and MIC 62.5 μg mL−1) against MRSA.190
A number of peptides were also reported to eradicate resistant bacterial strains. For instance, a small, <500 Da factor, isolated from the blowfly, Lucilia sericata, or Maggot, showed potent, thermally stable, protease resistant antibacterial activity against MRSA.191 Mastoparan-1 peptide, isolated from Polybia paulista (Neotropical social wasp), had MIC 0.001–0.019 μM against MRSA.192 Mauriporin, isolated from Androctonus mauritanicus (Fat tailed scorpion), had MIC 5–10 μM against MRSA.193 Pro10-1D (derived from protaetiamycine), isolated from Protaetia brevitarsis (White-spotted flower chafer beetle) had MIC 4 μM against MDR.194 A3 peptide, is modified version for AamAP1 which is a novel HDP that belongs to the venom of the North African scorpion Androctonus amoeruxi. A3 with conventional antibiotics caused a synergistic antimicrobial behavior that resulted in decreasing the MIC value for A3 peptide as low as 0.125 μM against MDR.195 DLP4 peptide, isolated from Hermetia illucens larvae, had antibacterial activity against MRSA.196 Lycotoxins I, was identified from venom of the wolf spider Lycosa carolinensis, had anti-MRSA USA300.197,198 Arenicin-3, isolated from Arenicola marina (sandworm), had considerable antimicrobial activity even against XDR (extensive drug resistance) and MDR strains as Acinetobacter baumannii, Pseudomonas aeruginosa, E. coli and Klebsiella pneumoniae.199 Maculatin 1.3 peptide, isolated from frog Litoria eucnemis, showed anti-MRSA in a concentration dependent manner.198,200 IP, isolated from the hard tick Ixodes persulcatus, has MIC 0.625–2.5 μg mL−1 for MRSA.201
3.5. Bacteria, fungi, higher fungi (mushrooms) derived natural products
Fig. 12 illustrates selected bacteria, fungi, higher fungi (mushrooms) derived natural products. Most reports on the activity of the following isolated compounds against MDR bacteria relate to in vitro studies. Mollicellin S 358, mollicellin T 359, and mollicellin U 360, mollicellin D 361 and mollicellin H 362, isolated from Chaetomium brasiliense SD-596 fungus, exhibited significant anti-MRSA, with MIC 6.25–12.5 μg mL−1.202 Fusidic acid 363, the steroid-like topical antibiotics which was isolated from Fusidium coccineum or Acremonium fusidioides fungus, showed activity against MRSA.203 Pleuromutilin 364 isolated from Pleurotus mutilis fungus, showed activity against MRSA.204 Osmundalactone 365, 5-hydroxy-hex-2-en-4-olide 366, spiromentins C 367, isolated from Tapinella atrotomentosa mushroom. These compounds proved to possess significant antibacterial activity against multi-resistant Acinetobacter baumannii and extended-spectrum β-lactamase (ESBL)-producing E. coli.205 CoQ0 368, isolated from Antrodia cinnamomea fungus, showed strong MRSA growth inhibition with MIC 7.81 μg mL−1. CoQ0 368 was found to eradicate biofilm MRSA efficiently and reduce the biofilm thickness. The compound 368 has also bactericidal activity against MRSA by inhibiting DNA polymerase and topoisomerases. The proteomic assay showed that CoQ0 380 also reduced the ribosomal proteins.206 Rubellins B 369, C 370, D 371, and E 372 and caeruleoramularin 373, isolated from fungus Ramularia collo-cygni, showed activities against Gram-negative bacteria, including MDR strains, such as S. aureus (SG) 511, S. aureus 134/94 (MRSA), B. subtilis (ATCC) 6633, Mycobacterium vaccae (IMET) 10670, or Enterococcus faecalis 1528 (VRE).207 Viriditoxin 374 which is xanthoradones relative structure, produced by Penicillium radicum FKI-3765-2, exhibit anti-MRSA by inhibiting FtsZ, the bacterial tubulin homolog which is crucial in septum formation.208
 | ||
| Fig. 12 Selected bacteria, fungi, higher fungi (mushrooms) derived natural products. | ||
Moreover, the peptide antibiotics tripropeptins A, B, C, D, and Z were isolated from cultured cells and broth of Lysobacter sp. Tripropeptins are active against Gram-positive bacteria including MRSA in vitro.209 Also, serrawettin W1, cyclodepsipeptide isolated from Gram-negative bacterium Serratia marcescens, inhibited the growth of nine different MRSA isolates.210 In addition, plectasin peptide, isolated from a fungus Pseudoplectania nigrella, showed especially activity against resistance strains of S. pneumoniae, S. aureus, S. epidermidis, and S. pyogenes with MIC 32–64, 32, 8, and 0.125 μg mL−1, respectively.211
3.6. Minerals
Nanomedicine is a budding branch of medicine that uses advancement in nanotechnology for the prevention and treatment of infectious disease. The advancement in nanotechnology has become an effective approach for the fabrication of metallic/salt bulk materials in nanosized particles for the treatment against drug-resistant microbes. There are many metallic/salt nanoparticles that have efficient activity against MDR microbes with different mechanism as titanium,212 zinc,213–215 copper,216 gold,217,218 silver 398,218,219 iron 399,215 sulphur,220 etc. These nanoparticles (NPs) have developed antibacterial activities against MDR microorganisms via adhesion to the bacterial cell membrane, followed by cell penetration, causing various structural disruptions and dysfunction through reactive oxygen species (ROS) generation which induced oxidative stress and inhibition the formation of polymeric matrix from substrate in the form of biofilm.4. Alternative branches research on developing new drugs for multi-drugs resistance pathogens
In addition to natural products, several remedies have been investigated as prebiotics, probiotics, synbiotics, bacteriophages, nanoparticles, bacteriocins, antimicrobial peptides, innate defense regulator peptides, peptidomimetics, vaccines and immune stimulation etc., each one providing its own benefits and limitations. Prebiotics which are nondigestible compounds that are selectively fermented by commensal microbiota in the human gut and hold an appropriate growth habitat for commensals and raise diversification within the microbiome, with improving human health.221 Sources of prebiotics include fructose, glucose, xylo-oligosaccharide, lactulose, and inulin.222 The metabolism of prebiotics by commensal organisms provides metabolic outputs as the short-chain fatty acids (SCFAs) propionate, butyrate, and acetate. SCFAs improve the barrier function of the gut through various mechanisms, incorporating the arrangement of energy for enterocytes; upregulation of tight junctures between cells of the epithelial layer promotion of mucus manufacture; and management of regulatory T-cells and T-helper 17 cell function to reduce inflammation.223 Through these processes, prebiotics support to both build up the population of commensal organisms and reduce colonization by enteric organisms. Probiotics vary from prebiotics in that probiotics are living bacteria or fungi that are directly employed and provide a health benefit to the host. Like prebiotics, probiotics expend their effect through the manufacture of SCFAs from metabolic precursors, leading to the same downstream effects of immune modulation and raised mucosal barrier function.223 Probiotics may have the extra effect of making their own antimicrobial compounds, as well as physically covering the epithelial niche and inhibiting the qualification for other pathogens to colonize the enteric microbiome.223 Whereas prebiotics produce an indirect effect on the microbiome through metabolic routes and expansion of commensal organisms, probiotics expend a further direct effect. Other than commercialized crops, probiotics are naturally developing in fermented foods such as yogurt, cheese, kimchi, and sauerkraut.224 While synbiotics are the mix of both prebiotic and probiotic.223 As such, their technique of action incorporates both the indirect effect of the metabolic precursor (prebiotics) on SCFA, and the direct modulation of organisms (probiotics) within the enteric microbial community. Synbiotics are generally applicable over the counter in a mixture of both probiotic strains and prebiotic fibers. Probiotic strains usually covered in synbiotics include Bifidobacterium species, Lactobacilli, and S. boulardii; the prebiotic it is added to an oligosaccharide such as fructose-oligosaccharide or inulin.225 The service of microbiome manipulation with prebiotics, probiotics, and synbiotics is in its infancy related with alternative methods. A survey of the present experimental literature can give no direct conclusions concerning the effectiveness of these measures; however, as the field increases in both the recognition of the microbiome and our ability to handle it, prebiotics, probiotics, and synbiotics are expected to participate a distinguished role. For today, these supplements look safe and are well accepted in most communities. Further analysis may accurately determine their role as an option method for fighting antimicrobial resistance. These nutritionally based remedies should continue to be utilized in partnership with other demonstrated techniques, such as antibiotic management and progress in hygiene and sterilization practices, to aid in the reduction of colonization with MDROs.225Nano-formulation of natural products provided many benefits, such as targeted drug delivery, raised component solubility, diminished dose, enhanced absorption, diminished metabolism, and enhanced bioavailability.226 It serves in increasing stability and achieving rigor targeting, with raising the efficiency of phytoconstituent. This can be carried out by encapsulation of natural products in a convenient carrier system such as nanoparticles, liposomes, and nano-emulsions, which can transform an inadequately available herbal drug into a successfully bioavailable drug candidate. Despite numerous advantages, the harmful effects of nano-formulation are connected to their minor sizes and inherent toxicity to the surface. This can be overcome by the development of natural products nano-formulation.226
Undoubtedly, bacteriophages presented great diversification and have great capacity for progress as antimicrobial therapy. Bacteriophages are a virus affected bacterium. Their potential in managing MDR pathogens is owing to their specificity and efficiency in generating harmful effects in the host bacterium by cell lysis. Phage therapy employing has been introduced via intravenous, and oral passages, and for vaccine issue. Some advantages of using phage therapy cover lower developmental costs, 100% bactericidal nature, high specificity, and the demand of only a single dose at the infection site. The disadvantage for phage therapy was the genetic material in temperate phage could raise the virulence of species of bacteria through transduction of virulence genes.6 To ensure the maximum effectiveness of clinical phage therapy, fast-track investigation requires to be performed on pathogenic bacteria, followed by isolation, identification, and evaluation against individual phage strains. Looking at the narrow armamentarium of antibiotics useable and a deficiency in management of newer ones, phages are a nature's gift, which is safe and efficient method as an option remedy. To reduce individual limitations of a therapy, a combination therapy approach is developed. The prospects of phage preparations to be handled in combination with antibiotics, probiotics, and vaccines against resistant pathogenic bacteria can serve reduce illnesses significantly.6
5. Conclusion
The research aimed to unravel novel natural antibiotics therapy against MDR bacterial strains which has been recognized of high priority, particularly in the last 20 years. Following the clear knowledge of resistance mechanism, various approaches have been followed. Various phytochemical classes were successfully isolated and investigated as antibacterial candidates from plants and their associated endophytes. Natural antimicrobial agents are characterized by their structural diversity, safety, and nontoxic quality. Examples include many bioactive scaffolding's secondary metabolites as phenolic compounds, terpenoids, volatile or EOs oils, flavonoids, and sulfur-containing compounds, peptides, and polyketides. Consequently, bioactive moieties with diverse chemical designs and modes of action are promising therapeutic manifestos for the introduction of novel bioactive compounds. However, further investigations are needed to assimilate mechanisms as well as the pharmacokinetic, and pharmacodynamics aspects of the bioactive compounds. Hence, nowadays several other alternatives were developed, including the use of prebiotics, probiotics, synbiotics, bacteriophages, bacteriocins, antimicrobial peptides, innate defense regulating peptides, peptidomimetics, and others.Unfortunately, a decrease in the developmental rate of antimicrobial agents has appeared in limited approval of novel antimicrobial drugs. Hence, building up of the microbial resistance to existing drugs could not be counterbalanced and following decrease in treatment opportunities. Several reasons account for this failure in developmental rate, including covering monetary issues, trouble in arranging clinical trials, and interruptions in investigating the treatment for acute infections, in addition to exorbitant enrolment of drug approval, despite of the several measures taken by the FDA to encourage the development of antimicrobial medications so that medical practitioners have access to a better number of treatment choices.
Author contributions
U. R. A., A. H. E., N. H. S., and A. Z. conceived and designed the work. A. H. E., N. H. S., K. M. A., N. G. S., M. B. E., Y. H. M., A. N. A, H. H. H., E. A. T. collected the data. A. H. E. wrote the manuscript. All authors contributed to & revised the article and approved the submitted version.Conflicts of interest
The authors declare no conflict of interest.References
- W. C. Reygaert, AIMS Microbiol., 2018, 4, 482 CAS.
- R. Klevens, M. Morrison, J. Nadle, S. Petit, K. Gershman and S. Ray, JAMA, 2007, 298, 1763–1771 CrossRef CAS PubMed.
- N. Cassir, J.-M. Rolain and P. Brouqui, Front. Microbiol., 2014, 5, 551 Search PubMed.
- K. S. Long and B. Vester, Antimicrob. Agents Chemother., 2012, 56, 603–612 CrossRef CAS PubMed.
- H. Nikaido, Annu. Rev. Biochem., 2009, 78, 119–146 CrossRef CAS PubMed.
- R. Vivas, A. A. T. Barbosa, S. S. Dolabela and S. Jain, Microb. Drug Resist., 2019, 25, 890–908 CrossRef CAS PubMed.
- Y. K. Schneider, Antibiotics, 2021, 10, 842 CrossRef CAS PubMed.
- S. E. Rossiter, M. H. Fletcher and W. M. Wuest, Chem. Rev., 2017, 117, 12415–12474 CrossRef CAS PubMed.
- A. G. Atanasov, S. B. Zotchev, V. M. Dirsch and C. T. Supuran, Nat. Rev. Drug Discovery, 2021, 20, 200–216 CrossRef CAS PubMed.
- A. S. Abdel-Razek, M. E. El-Naggar, A. Allam, O. M. Morsy and S. I. Othman, Processes, 2020, 8, 470 CrossRef CAS.
- L. Poirel, J.-Y. Madec, A. Lupo, A.-K. Schink, N. Kieffer, P. Nordmann and S. Schwarz, Microbiol. Spectrum, 2018, 6, 6.4.14 CrossRef PubMed.
- N. He, P. Wang, P. Wang, C. Ma and W. Kang, BMC Complementary Altern. Med., 2018, 18, 1–9 Search PubMed.
- S. K. Tanaka, J. Steenbergen and S. Villano, Bioorg. Med. Chem., 2016, 24, 6409–6419 CrossRef CAS PubMed.
- S. Shiota, M. Shimizu, J. i. Sugiyama, Y. Morita, T. Mizushima and T. Tsuchiya, Microbiol. Immunol., 2004, 48, 67–73 CrossRef CAS PubMed.
- V. Lorenzi, A. Muselli, A. F. Bernardini, L. Berti, J.-M. Pagès, L. Amaral and J.-M. Bolla, Antimicrob. Agents Chemother., 2009, 53, 2209–2211 CrossRef CAS PubMed.
- E. O. Sousa, N. F. Silva, F. F. Rodrigues, A. R. Campos, S. G. Lima and J. G. M. Costa, Pharmacogn. Mag., 2010, 6, 79 CrossRef CAS PubMed.
- J. Knowles, S. Roller, D. B. Murray and A. Naidu, Appl. Environ. Microbiol., 2005, 71, 797–803 CrossRef CAS PubMed.
- S. A. Burt, R. van der Zee, A. P. Koets, A. M. de Graaff, F. van Knapen, W. Gaastra, H. P. Haagsman and E. J. Veldhuizen, Appl. Environ. Microbiol., 2007, 73, 4484–4490 CrossRef CAS PubMed.
- S.-K. Yang, K. Yusoff, M. Ajat, W. Thomas, A. Abushelaibi, R. Akseer, S.-H. E. Lim and K.-S. Lai, PloS one, 2019, 14, e0214326 CrossRef CAS PubMed.
- H. Koo, S. Pearson, K. Scott-Anne, J. Abranches, J. Cury, P. Rosalen, Y. Park, R. Marquis and W. Bowen, Oral Microbiol. Immunol., 2002, 17, 337–343 CrossRef CAS PubMed.
- F. I. Gomes, P. Teixeira, J. Azeredo and R. Oliveira, Curr. Microbiol., 2009, 59, 118–122 CrossRef CAS PubMed.
- K. Masako, K. Yusuke, I. Hideyuki, M. Atsuko, M. Yoshiki, M. Kayoko and K. Makoto, J. Dermatol. Sci., 2005, 38, 207–213 CrossRef CAS PubMed.
- A. Sayout, A. Ouarhach, R. Rabie, I. Dilagui, N. Soraa and A. Romane, Chem. Biodiversity, 2020, 17, e1900496 CrossRef CAS PubMed.
- K. P. Devi, S. A. Nisha, R. Sakthivel and S. K. Pandian, J. Ethnopharmacol., 2010, 130, 107–115 CrossRef CAS PubMed.
- A. O. Gill and R. A. Holley, Appl. Environ. Microbiol., 2004, 70, 5750–5755 CrossRef CAS PubMed.
- R. Di Pasqua, G. Mamone, P. Ferranti, D. Ercolini and G. Mauriello, Proteomics, 2010, 10, 1040–1049 CrossRef CAS PubMed.
- C. Niu, S. Afre and E. S. Gilbert, Lett. Appl. Microbiol., 2006, 43, 489–494 CrossRef CAS PubMed.
- N. Gallucci, C. Casero, M. Oliva, J. Zygadlo and M. Demo, Mol. Med. Chem., 2006, 10, 30–32 Search PubMed.
- S.-H. Mun, D.-K. Joung, Y.-S. Kim, O.-H. Kang, S.-B. Kim, Y.-S. Seo, Y.-C. Kim, D.-S. Lee, D.-W. Shin and K.-T. Kweon, Phytomedicine, 2013, 20, 714–718 CrossRef CAS PubMed.
- K. Bogdanova, M. Röderova, M. Kolar, K. Langova, M. Dusek, P. Jost, K. Kubelkova, P. Bostik and J. Olsovska, Res. Microbiol., 2018, 169, 127–134 CrossRef CAS PubMed.
- L. Bocquet, S. Sahpaz, N. Bonneau, C. Beaufay, S. Mahieux, J. Samaillie, V. Roumy, J. Jacquin, S. Bordage and T. Hennebelle, Molecules, 2019, 24, 1024 CrossRef CAS PubMed.
- H.-H. Yu, K.-J. Kim, J.-D. Cha, H.-K. Kim, Y.-E. Lee, N.-Y. Choi and Y.-O. You, J. Med. Food, 2005, 8(4), 454–461 CrossRef CAS PubMed.
- M. Mohtar, S. A. Johari, A. R. Li, M. M. Isa, S. Mustafa, A. M. Ali and D. F. Basri, Curr. Microbiol., 2009, 59, 181–186 CrossRef CAS PubMed.
- G. O'Donnell and S. Gibbons, Phytother. Res., 2007, 21, 653–657 CrossRef PubMed.
- W. Maneerat, W. Phakhodee, T. Ritthiwigrom, S. Cheenpracha, T. Promgool, K. Yossathera, S. Deachathai and S. Laphookhieo, Fitoterapia, 2012, 83, 1110–1114 CrossRef CAS PubMed.
- W. Maneerat, W. Phakhodee, S. Cheenpracha, T. Ritthiwigrom, S. Deachathai and S. Laphookhieo, Phytochemistry, 2013, 88, 74–78 CrossRef CAS PubMed.
- A. Maurya, G. R. Dwivedi, M. P. Darokar and S. K. Srivastava, Chem. Biol. Drug Des., 2013, 81, 484–490 CrossRef CAS PubMed.
- G. R. Dwivedi, A. Maurya, D. K. Yadav, V. Singh, F. Khan, M. K. Gupta, M. Singh, M. P. Darokar and S. K. Srivastava, J. Biomol. Struct. Dyn., 2019, 37, 1307–1325 CrossRef CAS PubMed.
- K. Ponnusamy, M. Ramasamy, I. Savarimuthu and M. G. Paulraj, Scand. J. Infect. Dis., 2010, 42, 500–505 CrossRef CAS PubMed.
- R. Hamoud, J. Reichling and M. Wink, Drug Metab. Lett., 2014, 8, 119–128 CrossRef CAS PubMed.
- J.-G. Choi, O.-H. Kang, H.-S. Chae, B. Obiang-Obounou, Y.-S. Lee, Y.-C. Oh, M.-S. Kim, D.-W. Shin, J.-A. Kim and Y.-H. Kim, Appl. Biochem. Biotechnol., 2010, 160, 2467–2474 CrossRef CAS PubMed.
- R. Rodríguez-Guzmán, L. C. J. Fulks, M. M. Radwan, C. L. Burandt and S. A. Ross, Planta Med., 2011, 77, 1542–1544 CrossRef PubMed.
- R. S. Costa, M. O. Lins, M. L. Hyaric, T. F. Barros and E. S. Velozo, Rev. Bras. Farmacogn., 2017, 27, 195–198 CrossRef CAS.
- G. Y. Zuo, F. Y. Meng, X. Y. Hao, Y. L. Zhang, G. C. Wang and G. L. Xu, J. Pharm. Pharm. Sci., 2008, 11, 90–94 CAS.
- G.-Y. Zuo, Y. Li, T. Wang, J. Han, G.-C. Wang, Y.-L. Zhang and W.-D. Pan, Molecules, 2011, 16, 9819–9826 CrossRef CAS PubMed.
- S. Yin, G. Rao, J. Wang, L. Luo, G. He, C. Wang, C. Ma, X. Luo, Z. Hou and G. Xu, PloS one, 2015, 10, e0143863 CrossRef PubMed.
- X. Pan, S. A. Bligh and E. Smith, Phytother. Res., 2014, 28, 305–307 CrossRef CAS PubMed.
- Y. Sakagami, M. Mimura, K. Kajimura, H. Yokoyama, M. Iinuma, T. Tanaka and M. Ohyama, Lett. Appl. Microbiol., 1998, 27, 98–100 CrossRef CAS PubMed.
- H. Periasamy, S. Iswarya, N. Pavithra, S. Senthilnathan and A. Gnanamani, Lett. Appl. Microbiol., 2019, 69, 41–49 CAS.
- H. X. Xu and S. F. Lee, Phytother. Res., 2001, 15, 39–43 CrossRef CAS PubMed.
- D. Dey, R. Ray and B. Hazra, Phytother. Res., 2014, 28, 1014–1021 CrossRef CAS PubMed.
- C. M. Schempp, K. Pelz, A. Wittmer, E. Schöpf and J. C. Simon, Lancet, 1999, 353, 2129 CrossRef CAS.
- H. Dharmaratne, W. Wijesinghe and V. Thevanasem, J. Ethnopharmacol., 1999, 66, 339–342 CrossRef CAS PubMed.
- L. Bunalema, G. W. Fotso, P. Waako, J. Tabuti and S. O. Yeboah, BMC Complementary Altern. Med., 2017, 17, 1–6 CrossRef PubMed.
- F. Hanawa, M. Okamoto and G. N. Towers, Photochem. Photobiol., 2002, 76, 51–56 CrossRef CAS PubMed.
- H. Muroi and I. Kubo, Biosci., Biotechnol., Biochem., 1994, 58, 1925–1926 CrossRef CAS.
- Y. Sato, H. Oketani, K. Singyouchi, T. Ohtsubo, M. Kihara, H. Shibata and T. Higuti, Biol. Pharm. Bull., 1997, 20, 401–404 CrossRef CAS PubMed.
- D.-Y. Shin, H.-S. Kim, K.-H. Min, S.-S. Hyun, S.-A. Kim, H. Huh, E.-C. Choi, Y. H. Choi, J. Kim and S.-H. Choi, Chem. Pharm. Bull., 2000, 48, 1805–1806 CrossRef CAS PubMed.
- C. Gaspar-Marques, P. Rijo, M. F. Simões, M. A. Duarte and B. Rodriguez, Phytomedicine, 2006, 13, 267–271 CrossRef CAS PubMed.
- M. Iinuma, H. Tosa, T. Tanaka, F. Asai, Y. Kobayashl, R. Shimano and K.-I. Miyauchi, J. Pharm. Pharmacol., 1996, 48, 861–865 CrossRef CAS PubMed.
- Y. Sato, H. Oketani, T. Yamada, K.-I. Singyouchi, T. Ohtsubo, M. Kihara, H. Shibata and T. Higuti, J. Pharm. Pharmacol., 1997, 49, 1042–1044 CrossRef CAS PubMed.
- M. Sato, H. Tsuchiya, T. Miyazaki, M. Ohyama, T. Tanaka and M. Iinuma, Lett. Appl. Microbiol., 1995, 21, 219–222 CrossRef CAS PubMed.
- O. M. Demgne, J. F. T. Mbougnia, A. J. Seukep, A. T. Mbaveng, M. Tene, P. Nayim, B. E. Wamba, M.-G. F. Guefack, V. P. Beng and P. Tane, Adv. Tradit. Med., 2021, 1–12 Search PubMed.
- R. S. Taylor and G. N. Towers, Phytochemistry, 1998, 47, 631–634 CrossRef CAS PubMed.
- K. Kawazoe, Y. Tsubouchi, N. Abdullah, Y. Takaishi, H. Shibata, T. Higuti, H. Hori and M. Ogawa, J. Nat. Prod., 2003, 66, 538–539 CrossRef CAS PubMed.
- P. E. Ordóñez, C. L. Quave, W. F. Reynolds, K. I. Varughese, B. Berry, P. J. Breen, O. Malagón, M. S. Smeltzer and C. M. Compadre, J. Ethnopharmacol., 2011, 137, 1055–1059 CrossRef PubMed.
- M. M. Farimani, A. Taleghani, A. Aliabadi, A. Aliahmadi, M. A. Esmaeili, N. N. Sarvestani, H. R. Khavasi, M. Smieško, M. Hamburger and S. N. Ebrahimi, Planta Med., 2016, 82, 1279–1285 CrossRef CAS PubMed.
- Y. Qiao, Y. Liu, X. Duan, C. Chen, J. Liu, H. Zhu, Y. Xue and Y. Zhang, Tetrahedron, 2018, 74, 3852–3857 CrossRef CAS.
- H. Siddique, B. Pendry and M. M. Rahman, Molecules, 2019, 24, 385 CrossRef PubMed.
- V. K. Gupta, S. Verma, A. Pal, S. K. Srivastava, P. K. Srivastava and M. P. Darokar, Appl. Microbiol. Biotechnol., 2013, 97, 9121–9131 CrossRef CAS PubMed.
- M. Stavri, A. Paton, B. W. Skelton and S. Gibbons, J. Nat. Prod., 2009, 72, 1191–1194 CrossRef CAS PubMed.
- E. Smith, E. Williamson, M. Zloh and S. Gibbons, Phytother. Res., 2005, 19, 538–542 CrossRef CAS PubMed.
- A. C. F. Soares, P. M. Matos, K. F. d. Silva, C. H. Martins, R. Veneziani, S. R. Ambrósio, H. J. Dias, R. A. d. Santos and V. C. Heleno, J. Braz. Chem. Soc., 2019, 30, 333–341 CAS.
- A. de Breij, T. Karnaoukh, J. Schrumpf, P. Hiemstra, P. Nibbering, J. van Dissel and P. de Visser, Eur. J. Clin. Microbiol. Infect. Dis., 2016, 35, 555–562 CrossRef CAS PubMed.
- M.-T. Gutierrez-Lugo, M. P. Singh, W. M. Maiese and B. N. Timmermann, J. Nat. Prod., 2002, 65, 872–875 CrossRef CAS PubMed.
- C. Latha, V. D. Shriram, S. S. Jahagirdar, P. K. Dhakephalkar and S. R. Rojatkar, J. Ethnopharmacol., 2009, 123, 522–525 CrossRef CAS PubMed.
- J.-M. He, S.-C. Sun, Z.-L. Sun, J.-T. Chen and Q. Mu, Int. J. Antimicrob. Agents, 2019, 53, 70–73 CrossRef CAS PubMed.
- S. Schmidt, K. Heymann, M. F. Melzig, S. Bereswill and M. M. Heimesaat, Planta Med., 2016, 82, 1540–1545 CrossRef CAS PubMed.
- S. A. Sarkisian, M. Janssen, H. Matta, G. Henry, K. L. LaPlante and D. C. Rowley, Phytother. Res., 2012, 26, 1012–1016 CrossRef CAS PubMed.
- D. Wang, K. Xie, D. Zou, M. Meng and M. Xie, Mol. Med. Rep., 2018, 18, 827–833 CAS.
- H. Zhang, Y. Luan, S. Jing, Y. Wang, Z. Gao, P. Yang, Y. Ding, L. Wang, D. Wang and T. Wang, Biochem. Pharmacol., 2020, 178, 114024 CrossRef CAS PubMed.
- B. C. Chan, M. Ip, C. B. Lau, S. Lui, C. Jolivalt, C. Ganem-Elbaz, M. Litaudon, N. E. Reiner, H. Gong and R. H. See, J. Ethnopharmacol., 2011, 137, 767–773 CrossRef CAS PubMed.
- M. Różalski, E. Walencka, B. Różalska and H. Wysokińska, Phytomedicine, 2007, 14, 31–35 Search PubMed.
- P. Y. Chung, L. Y. Chung and P. Navaratnam, Fitoterapia, 2014, 94, 48–54 CrossRef CAS PubMed.
- L. F. Leandro, M. J. O. Cardoso, S. D. C. Silva, M. G. M. Souza, R. C. S. Veneziani, S. R. Ambrosio and C. H. G. Martins, J. Med. Microbiol., 2014, 63, 1649–1653 CrossRef CAS PubMed.
- D. G. Lee, H. J. Jung and E.-R. Woo, Arch. Pharmacal Res., 2005, 28, 1031–1036 CrossRef CAS PubMed.
- G.-S. Lee, E.-S. Kim, S.-I. Cho, J.-H. Kim, G. Choi, Y.-S. Ju, S.-H. Park, S.-I. Jeong and H.-J. Kim, J. Korean Soc. Appl. Biol. Chem., 2010, 53, 290–296 CrossRef CAS.
- E. Kim, S. Jeong, J. Kim, C. Park, S. Kim, J. Kim, K. Lee, S. Lee, H. So and R. Park, J. Microbiol. Biotechnol., 2009, 19, 1576–1581 CrossRef CAS PubMed.
- A. Rahman, A. Chowdhury, H.-A. Ali, S. Z. Raihan, M. S. Ali, L. Nahar and S. D. Sarker, J. Nat. Med., 2009, 63, 41–45 CrossRef CAS PubMed.
- Y. Miyasaki, J. D. Rabenstein, J. Rhea, M.-L. Crouch, U. M. Mocek, P. E. Kittell, M. A. Morgan, W. S. Nichols, M. Van Benschoten and W. D. Hardy, PloS one, 2013, 8, e61594 CrossRef CAS PubMed.
- J. M. McRae, Q. Yang, R. J. Crawford and E. A. Palombo, J. Ethnopharmacol., 2008, 116, 554–560 CrossRef CAS PubMed.
- P. M. Giang, P. T. Son, K. Matsunami and H. Otsuka, J. Nat. Med., 2006, 60, 93–95 CrossRef CAS.
- V. M. Navarro-García, J. Luna-Herrera, M. Rojas-Bribiesca, P. Álvarez-Fitz and M. Y. Ríos, Molecules, 2011, 16, 7357–7364 CrossRef PubMed.
- S. Sureram, S. P. Senadeera, P. Hongmanee, C. Mahidol, S. Ruchirawat and P. Kittakoop, Bioorg. Med. Chem. Lett., 2012, 22, 2902–2905 CrossRef CAS PubMed.
- R. León-Díaz, M. Meckes-Fischer, L. Valdovinos-Martínez, M. G. Campos, R. Hernández-Pando and M. A. Jiménez-Arellanes, Arch. Med. Res., 2013, 44, 99–104 CrossRef PubMed.
- A. H. Uc-Cachón, R. Borges-Argáez, S. Said-Fernández, J. Vargas-Villarreal, F. González-Salazar, M. Méndez-González, M. Cáceres-Farfán and G. M. Molina-Salinas, Pulm. Pharmacol. Ther., 2014, 27, 114–120 CrossRef PubMed.
- W. S. Jang, M. Jyoti, S. Kim, K.-W. Nam, T. K. Q. Ha, W. K. Oh and H.-Y. Song, J. Nat. Med., 2016, 70, 127–132 CrossRef CAS PubMed.
- D. Lakshmanan, J. Werngren, L. Jose, K. Suja, M. S. Nair, R. L. Varma, S. Mundayoor, S. Hoffner and R. A. Kumar, Fitoterapia, 2011, 82, 757–761 CrossRef CAS PubMed.
- P. Kumar, A. Singh, U. Sharma, D. Singh, M. Dobhal and S. Singh, Pulm. Pharmacol. Ther., 2013, 26, 332–335 CrossRef CAS PubMed.
- T. Siriyong, P. Srimanote, S. Chusri, B.-e. Yingyongnarongkul, C. Suaisom, V. Tipmanee and S. P. Voravuthikunchai, BMC Complementary Altern. Med., 2017, 17, 1–7 CrossRef PubMed.
- M. Lamontagne Boulet, C. Isabelle, I. Guay, E. Brouillette, J.-P. Langlois, P.-É. Jacques, S. Rodrigue, R. Brzezinski, P. B. Beauregard and K. Bouarab, Antimicrob. Agents Chemother., 2018, 62, e02197–e02217 CrossRef PubMed.
- J. Reiter, N. Levina, M. Van der Linden, M. Gruhlke, C. Martin and A. J. Slusarenko, Molecules, 2017, 22, 1711 CrossRef PubMed.
- D. Lechner, S. Gibbons and F. Bucar, J. Antimicrob. Chemother., 2008, 62, 345–348 CrossRef CAS PubMed.
- G. Belofsky, D. Percivill, K. Lewis, G. P. Tegos and J. Ekart, J. Nat. Prod., 2004, 67, 481–484 CrossRef CAS PubMed.
- A.-N. Abulrob, M. T. Suller, M. Gumbleton, C. Simons and A. D. Russell, Phytochemistry, 2004, 65, 3021–3027 CrossRef CAS PubMed.
- S.-H. Mun, D.-K. Joung, S.-B. Kim, S.-J. Park, Y.-S. Seo, R. Gong, J.-G. Choi, D.-W. Shin, J.-R. Rho and O.-H. Kang, Foodborne Pathog. Dis., 2014, 11, 234–239 CrossRef CAS PubMed.
- K.-W. Jeong, J.-Y. Lee, D.-I. Kang, J.-U. Lee, S. Y. Shin and Y. Kim, J. Nat. Prod., 2009, 72, 719–724 CrossRef CAS PubMed.
- B. S. F. Bazzaz, Z. Memariani, Z. Khashiarmanesh, M. Iranshahi and M. Naderinasab, Braz. J. Microbiol., 2010, 41, 574–580 CrossRef CAS PubMed.
- M. Song, Y. Liu, T. Li, X. Liu, Z. Hao, S. Ding, P. Panichayupakaranant, K. Zhu and J. Shen, Adv. Sci., 2021, 8, 2100749 CrossRef CAS PubMed.
- J. Zhang, Z. Wang, Z. Song, L. Karthik, C. Hou, G. Zhu, L. Jiang, J. Han, R. Ma and L. Li, Synth. Syst. Biotechnol., 2019, 4, 173–179 CrossRef PubMed.
- P. Ratnaweera, E. D. de Silva, R. L. Wijesundera and R. J. Andersen, J. Natl. Sci. Found. Sri Lanka, 2016, 44 Search PubMed.
- E. S. Elkhayat, S. R. Ibrahim, G. A. Mohamed and S. A. Ross, Nat. Prod. Res., 2016, 30, 814–820 CrossRef CAS PubMed.
- Q. Deng, G. Li, M. Sun, X. Yang and J. Xu, Nat. Prod. Res., 2020, 34, 1404–1408 CrossRef CAS PubMed.
- P. B. Ratnaweera, D. E. Williams, E. D. de Silva, R. L. Wijesundera, D. S. Dalisay and R. J. Andersen, Mycology, 2014, 5, 23–28 CrossRef PubMed.
- Z. Wu, X. Zhang, W. H. A. Anbari, Q. Zhou, P. Zhou, M. Zhang, F. Zeng, C. Chen, Q. Tong and J. Wang, J. Nat. Prod., 2019, 82, 2653–2658 CrossRef CAS PubMed.
- S. S. El-Hawary, A. S. Moawad, H. S. Bahr, U. R. Abdelmohsen and R. Mohammed, RSC Adv., 2020, 10, 22058–22079 RSC.
- W. Zhang, X. Lu, L. Huo, S. Zhang, Y. Chen, Z. Zou and H. Tan, J. Nat. Prod., 2021, 84, 1715–1724 CrossRef CAS PubMed.
- M. M. Alshaibani, J. Jalil, N. M. Sidik, R. Edrada-Ebel and N. M. Zin, Drug Des., Dev. Ther., 2016, 10, 1817 CAS.
- H.-Y. Yang, Y.-Q. Duan, Y.-K. Yang, J. Li, X. Liu, L. Ye, Q.-L. Mi, W.-S. Kong, M. Zhou and G.-Y. Yang, Phytochem. Lett., 2017, 22, 266–269 CrossRef CAS.
- T. R. Nurunnabi, L. Nahar, S. Al-Majmaie, S. M. Rahman, M. H. Sohrab, M. M. Billah, F. M. Ismail, M. M. Rahman, G. P. Sharples and S. D. Sarker, Phytother. Res., 2018, 32, 348–354 CrossRef CAS PubMed.
- W.-L. Mei, B. Zheng, Y.-X. Zhao, H.-M. Zhong, X.-L. W. Chen, Y.-B. Zeng, W.-H. Dong, J.-L. Huang, P. Proksch and H.-F. Dai, Mar. Drugs, 2012, 10, 1993–2001 CrossRef CAS PubMed.
- Y.-B. Zeng, H. Wang, W.-J. Zuo, B. Zheng, T. Yang, H.-F. Dai and W.-L. Mei, Mar. Drugs, 2012, 10, 598–603 CrossRef CAS PubMed.
- J. Xu, RSC Adv., 2015, 5, 841–892 RSC.
- S. Klaiklay, V. Rukachaisirikul, Y. Sukpondma, S. Phongpaichit, J. Buatong and B. Bussaban, Arch. Pharmacal Res., 2012, 35, 1127–1131 CrossRef CAS PubMed.
- J. Kjer, V. Wray, R. Edrada-Ebel, R. Ebel, A. Pretsch, W. Lin and P. Proksch, J. Nat. Prod., 2009, 72, 2053–2057 CrossRef CAS PubMed.
- P. B. Ratnaweera, E. D. de Silva, D. E. Williams and R. J. Andersen, BMC Complementary Altern. Med., 2015, 15, 1–7 CrossRef CAS PubMed.
- S. R. Ibrahim, E. S. Elkhayat, G. A. Mohamed, A. I. Khedr, M. A. Fouad, M. H. Kotb and S. A. Ross, Phytochem. Lett., 2015, 14, 84–90 CrossRef CAS.
- W. Pongcharoen, V. Rukachaisirikul, S. Phongpaichit and J. Sakayaroj, Chem. Pharm. Bull., 2007, 55, 1404–1405 CrossRef CAS PubMed.
- V. Rukachaisirikul, J. Arunpanichlert, Y. Sukpondma, S. Phongpaichit and J. Sakayaroj, Tetrahedron, 2009, 65, 10590–10595 CrossRef CAS.
- Q.-X. Wang, S.-F. Li, F. Zhao, H.-Q. Dai, L. Bao, R. Ding, H. Gao, L.-X. Zhang, H.-A. Wen and H.-W. Liu, Fitoterapia, 2011, 82, 777–781 CrossRef CAS PubMed.
- A. R. Ola, D. Thomy, D. Lai, H. Brötz-Oesterhelt and P. Proksch, J. Nat. Prod., 2013, 76, 2094–2099 CrossRef CAS PubMed.
- C. P. Parlet, J. S. Kavanaugh, H. A. Crosby, H. A. Raja, T. El-Elimat, D. A. Todd, C. J. Pearce, N. B. Cech, N. H. Oberlies and A. R. Horswill, Cell Rep., 2019, 27, 187–198 CrossRef CAS PubMed.
- U. Sommart, V. Rukachaisirikul, Y. Sukpondma, S. Phongpaichit, J. Sakayaroj and K. Kirtikara, Chem. Pharm. Bull., 2008, 56, 1687–1690 CrossRef CAS PubMed.
- R. K. Dissanayake, P. B. Ratnaweera, D. E. Williams, C. D. Wijayarathne, R. L. Wijesundera, R. J. Andersen and E. D. de Silva, Mycology, 2016, 7, 1–8 CrossRef CAS PubMed.
- S. Shi, Y. Li, Y. Ming, C. Li, Z. Li, J. Chen and M. Luo, Rec. Nat. Prod., 2018, 12, 549–556 CrossRef CAS.
- R. Bara, A. H. Aly, A. Pretsch, V. Wray, B. Wang, P. Proksch and A. Debbab, J. Antibiot., 2013, 66, 491–493 CrossRef CAS PubMed.
- M. L. Macías-Rubalcava and R. E. Sánchez-Fernández, World J. Microbiol. Biotechnol., 2017, 33, 1–22 CrossRef PubMed.
- Y. Qiao, X. Zhang, Y. He, W. Sun, W. Feng, J. Liu, Z. Hu, Q. Xu, H. Zhu and J. Zhang, Sci. Rep., 2018, 8, 1–11 Search PubMed.
- R. Deshidi, S. Devari, M. Kushwaha, A. P. Gupta, R. Sharma, R. Chib, I. A. Khan, S. Jaglan and B. A. Shah, ChemistrySelect, 2017, 2, 364–368 CrossRef CAS.
- C.-S. Jiang, Z. Zhen-Fang, Y. Xiao-Hong, L. Le-Fu, G. Yu-Cheng, Y. Bo-Ping and G. Yue-Wei, Chin. J. Nat. Med., 2018, 16, 358–365 CAS.
- S. K. Deshmukh, S. A. Verekar and S. V. Bhave, Front. Microbiol., 2015, 5, 715 Search PubMed.
- J.-H. Pan, Y. Chen, Y.-H. Huang, Y.-W. Tao, J. Wang, Y. Li, Y. Peng, T. Dong, X.-M. Lai and Y.-C. Lin, Arch. Pharmacal Res., 2011, 34, 1177–1181 CrossRef CAS PubMed.
- C. Wang, J. Wang, Y. Huang, H. Chen, Y. Li, L. Zhong, Y. Chen, S. Chen, J. Wang and J. Kang, Molecules, 2013, 18, 1728–1740 CrossRef CAS PubMed.
- M. Leyte-Lugo, M. González-Andrade, M. a. d. C. González, A. E. Glenn, C. M. Cerda-García-Rojas and R. Mata, J. Nat. Prod., 2012, 75, 1571–1577 CrossRef CAS PubMed.
- D. U. Ganihigama, S. Sureram, S. Sangher, P. Hongmanee, T. Aree, C. Mahidol, S. Ruchirawat and P. Kittakoop, Eur. J. Med. Chem., 2015, 89, 1–12 CrossRef CAS PubMed.
- F. Liu, X.-L. Cai, H. Yang, X.-K. Xia, Z.-Y. Guo, J. Yuan, M.-F. Li, Z.-G. She and Y.-C. Lin, Planta Med., 2010, 76, 185–189 CrossRef CAS PubMed.
- A. R. Ola, A. Debbab, A. H. Aly, A. Mandi, I. Zerfass, A. Hamacher, M. U. Kassack, H. Brötz-Oesterhelt, T. Kurtan and P. Proksch, Tetrahedron Lett., 2014, 55, 1020–1023 CrossRef CAS.
- H.-L. Chen, W.-T. Zhao, Q.-P. Liu, H.-Y. Chen, W. Zhao, D.-F. Yang and X.-L. Yang, Fitoterapia, 2020, 141, 104475 CrossRef CAS PubMed.
- S. X. Yang, W. T. Zhao, H. Y. Chen, L. Zhang, T. K. Liu, H. P. Chen, J. Yang and X. L. Yang, Chem. Biodiversity, 2019, 16, e1900364 CrossRef PubMed.
- Y. Xiao-Long, S. Zhang, S. Shao-Jun, Y. Zhang, L. Du-Qiang and M. Zhang, Chin. J. Nat. Med., 2011, 9, 101–104 Search PubMed.
- A. Debbab, A. H. Aly, R. Edrada-Ebel, V. Wray, W. E. Müller, F. Totzke, U. Zirrgiebel, C. Schachtele, M. H. Kubbutat and W. H. Lin, J. Nat. Prod., 2009, 72, 626–631 CrossRef CAS PubMed.
- W.-j. Dai, H.-f. Dai, J. Wu, J. Liu and W.-l. Mei, Nat. Prod. Commun., 2009, 4, 1934578X0900401110 CrossRef.
- W.-J. Dai, J. Wu, Z. Han, W.-L. Mei and H.-F. Dai, J. Asian Nat. Prod. Res., 2009, 11, 704–709 CrossRef PubMed.
- L. Ding, A. Maier, H.-H. Fiebig, W.-H. Lin and C. Hertweck, Org. Biomol. Chem., 2011, 9, 4029–4031 RSC.
- K. Nuankeaw, B. Chaiyosang, T. Suebrasri, S. Kanokmedhakul, S. Lumyong and S. Boonlue, Mycoscience, 2020, 61, 16–21 CrossRef.
- S. F. Brady, M. P. Singh, J. E. Janso and J. Clardy, J. Am. Chem. Soc., 2000, 122, 2116–2117 CrossRef CAS.
- S. Klaiklay, V. Rukachaisirikul, S. Phongpaichit, C. Pakawatchai, S. Saithong, J. Buatong, S. Preedanon and J. Sakayaroj, Phytochem. Lett., 2012, 5, 738–742 CrossRef CAS.
- H.-X. Liao, T.-M. Shao, R.-Q. Mei, G.-L. Huang, X.-M. Zhou, C.-J. Zheng and C.-Y. Wang, Mar. Drugs, 2019, 17, 710 CrossRef CAS PubMed.
- P. Liu, D. Zhang, R. Shi, Z. Yang, F. Zhao and Y. Tian, 3 Biotech, 2019, 9, 1–9 Search PubMed.
- A. G. de Medeiros, D. C. Savi, P. Mitra, K. A. Shaaban, A. K. Jha, J. S. Thorson, J. Rohr and C. Glienke, Folia Microbiol., 2018, 63, 499–505 CrossRef CAS PubMed.
- H. K. Santra, S. Maity and D. Banerjee, Molecules, 2022, 27, 1459 CrossRef CAS PubMed.
- M. Figueroa, A. K. Jarmusch, H. A. Raja, T. El-Elimat, J. S. Kavanaugh, A. R. Horswill, R. G. Cooks, N. B. Cech and N. H. Oberlies, J. Nat. Prod., 2014, 77, 1351–1358 CrossRef CAS PubMed.
- N. Van Minh, N. T. Phat and D. N. Linh, Pharmacophore, 2021, 12 Search PubMed.
- P.-Y. Mai, M. Levasseur, D. Buisson, D. Touboul and V. Eparvier, Plants, 2019, 9, 47 CrossRef PubMed.
- H.-N. T. Vu, D. T. Nguyen, H. Q. Nguyen, H. H. Chu, S. K. Chu, M. V. Chau and Q.-T. Phi, Curr. Microbiol., 2018, 75, 1247–1255 CrossRef CAS PubMed.
- M. Aftab Uddin, S. Akter, M. Ferdous, B. Haidar, A. Amin, A. Shofiul Islam Molla, H. Khan and M. R. Islam, Sci. Rep., 2021, 11, 1–12 CrossRef PubMed.
- K. A. Qureshi, A. D. Bholay, P. K. Rai, H. A. Mohammed, R. A. Khan, F. Azam, M. Jaremko, A.-H. Emwas, P. Stefanowicz and M. Waliczek, Sci. Rep., 2021, 11, 1–21 CrossRef PubMed.
- U. F. Castillo, G. A. Strobel, E. J. Ford, W. M. Hess, H. Porter, J. B. Jensen, H. Albert, R. Robison, M. A. Condron and D. B. Teplow, Microbiology, 2002, 148, 2675–2685 CrossRef CAS PubMed.
- U. F. Castillo, G. A. Strobel, K. Mullenberg, M. M. Condron, D. B. Teplow, V. Folgiano, M. Gallo, R. Ferracane, L. Mannina and S. Viel, FEMS Microbiol. Lett., 2006, 255, 296–300 CrossRef CAS PubMed.
- U. Castillo, J. K. Harper, G. A. Strobel, J. Sears, K. Alesi, E. Ford, J. Lin, M. Hunter, M. Maranta and H. Ge, FEMS Microbiol. Lett., 2003, 224, 183–190 CrossRef CAS PubMed.
- M. G. Tozatti, D. S. Ferreira, L. G. B. Flauzino, T. da Silva Moraes, C. H. Martins, M. Groppo, M. L. A. e. Silva, A. H. Januario, P. M. Pauletti and W. R. Cunha, Nat. Prod. Commun., 2016, 11, 1934578X1601100419 CrossRef CAS.
- A. Pompilio, S. Pomponio, V. Di Vincenzo, V. Crocetta, M. Nicoletti, M. Piovano, J. A. Garbarino and G. Di Bonaventura, Future Microbiol., 2013, 8, 281–292 CrossRef CAS PubMed.
- G. Celenza, B. Segatore, D. Setacci, M. Perilli, F. Brisdelli, P. Bellio, M. Piovano, J. A. Garbarino, G. Amicosante and M. Nicoletti, Nat. Prod. Res., 2013, 27, 1528–1531 CrossRef CAS PubMed.
- M. Cankiliç, N. Sariozlu, M. Candan and N. Tay, Biomed. Res., 2017, 28(7), 3108–3113 Search PubMed.
- G. Shrestha, Exploring the Antibacterial, Antioxidant, and Anticancer Properties of Lichen Metabolites, Brigham Young University, 2015 Search PubMed.
- T. Kokubun, W. K. P. Shiu and S. Gibbons, Planta Med., 2007, 73, 176–179 CrossRef CAS PubMed.
- M. Lauterwein, M. Oethinger, K. Belsner, T. Peters and R. Marre, Antimicrob. Agents Chemother., 1995, 39, 2541–2543 CrossRef CAS PubMed.
- P. Bellio, B. Segatore, A. Mancini, L. Di Pietro, C. Bottoni, A. Sabatini, F. Brisdelli, M. Piovano, M. Nicoletti and G. Amicosante, Phytomedicine, 2015, 22, 223–230 CrossRef CAS PubMed.
- G. Celenza, B. Segatore, D. Setacci, P. Bellio, F. Brisdelli, M. Piovano, J. A. Garbarino, M. Nicoletti, M. Perilli and G. Amicosante, Phytomedicine, 2012, 19, 596–602 CrossRef CAS PubMed.
- J. M. Oh, Y. J. Kim, H.-S. Gang, J. Han, H.-H. Ha and H. Kim, Molecules, 2018, 23, 3068 CrossRef PubMed.
- Q.-X. Wang, L. Bao, X.-L. Yang, H. Guo, R.-N. Yang, B. Ren, L.-X. Zhang, H.-Q. Dai, L.-D. Guo and H.-W. Liu, Fitoterapia, 2012, 83, 209–214 CrossRef CAS PubMed.
- Q.-X. Wang, L. Bao, X.-L. Yang, D.-L. Liu, H. Guo, H.-Q. Dai, F.-H. Song, L.-X. Zhang, L.-D. Guo and S.-J. Li, Fitoterapia, 2013, 90, 220–227 CrossRef CAS PubMed.
- M. C. B. Martins, M. J. G. d. Lima, F. P. Silva, E. Azevedo-Ximenes, N. H. d. Silva and E. C. Pereira, Braz. Arch. Biol. Technol., 2010, 53, 115–122 CrossRef CAS.
- W. H. Choi and M. Jiang, J. Appl. Biomed., 2014, 12, 179–189 CrossRef.
- L. Borrelli, L. Varriale, L. Dipineto, A. Pace, L. F. Menna and A. Fioretti, Front. Microbiol., 2021, 12, 330 Search PubMed.
- R. Raghukumar, L. Vali, D. Watson, J. Fearnley and V. Seidel, Phytother. Res., 2010, 24, 1181–1187 CrossRef CAS PubMed.
- Y.-W. Chen, S.-R. Ye, C. Ting and Y.-H. Yu, J. Food Drug Anal., 2018, 26, 761–768 CrossRef CAS PubMed.
- L.-F. Zhou, J. Wu, S. Li, Q. Li, L.-P. Jin, C.-P. Yin and Y.-L. Zhang, ACS omega, 2021, 6, 4329–4334 CrossRef CAS PubMed.
- Y. Ma, M. Xu, H. Liu, T. Yu, P. Guo, W. Liu and X. Jin, AMB Express, 2021, 11, 1–11 CrossRef PubMed.
- Y. Correa, B. Cabanillas, V. Jullian, D. Álvarez, D. Castillo, C. Dufloer, B. Bustamante, E. Roncal, E. Neyra and P. Sheen, PloS one, 2019, 14, e0218837 CrossRef CAS PubMed.
- A. Bexfield, Y. Nigam, S. Thomas and N. A. Ratcliffe, Microbes Infect., 2004, 6, 1297–1304 CrossRef CAS PubMed.
- H. Memariani, M. Memariani, M. Shahidi-Dadras, S. Nasiri, M. M. Akhavan and H. Moravvej, Appl. Microbiol. Biotechnol., 2019, 103, 3265–3276 CrossRef CAS PubMed.
- A. Almaaytah, S. Tarazi, F. Alsheyab, Q. Al-Balas and T. Mukattash, Int. J. Pept. Res. Ther., 2014, 20, 397–408 CrossRef CAS.
- M. Krishnan, J. Choi, A. Jang and Y. Kim, Int. J. Mol. Sci., 2020, 21, 6216 CrossRef CAS PubMed.
- A. Almaaytah, A. Farajallah, A. Abualhaijaa and Q. Al-Balas, Molecules, 2018, 23, 1603 CrossRef PubMed.
- S.-I. Park, J.-W. Kim and S. M. Yoe, Dev. Comp. Immunol., 2015, 52, 98–106 CrossRef CAS PubMed.
- L. Yan and M. E. Adams, J. Biol. Chem., 1998, 273, 2059–2066 CrossRef CAS PubMed.
- J. Menousek, B. Mishra, M. L. Hanke, C. E. Heim, T. Kielian and G. Wang, Int. J. Antimicrob. Agents, 2012, 39, 402–406 CrossRef CAS PubMed.
- A. G. Elliott, J. X. Huang, S. Neve, J. Zuegg, I. A. Edwards, A. K. Cain, C. J. Boinett, L. Barquist, C. V. Lundberg and J. Steen, Nat. Commun., 2020, 11, 1–13 CrossRef PubMed.
- C. S. Brinkworth, J. H. Bowie, M. J. Tyler and J. C. Wallace, Aust. J. Chem., 2002, 55, 605–610 CrossRef CAS.
- N. Miyoshi, T. Saito, T. Ohmura, K. Kuroda, K. Suita, K. Ihara and E. Isogai, Parasites Vectors, 2016, 9, 1–11 CrossRef PubMed.
- P. Zhao, M. Yang, G. Zhu, B. Zhao, H. Wang, H. Liu, X. Wang, J. Qi, X. Yin and L. Yu, J. Antibiot., 2021, 74, 317–323 CrossRef CAS PubMed.
- M. Whitby, Int. J. Antimicrob. Agents, 1999, 12, S67–S71 CrossRef CAS PubMed.
- B. Li, Z. Zhang, J.-F. Zhang, J. Liu, X.-Y. Zuo, F. Chen, G.-Y. Zhang, H.-Q. Fang, Z. Jin and Y.-Z. Tang, Eur. J. Med. Chem., 2021, 223, 113624 CrossRef CAS PubMed.
- Z. Béni, M. Dékány, B. Kovács, B. Csupor-Löffler, Z. P. Zomborszki, E. Kerekes, A. Szekeres, E. Urbán, J. Hohmann and A. Ványolós, Molecules, 2018, 23, 1082 CrossRef PubMed.
- W.-L. Chou, T.-H. Lee, T.-H. Huang, P.-W. Wang, Y.-P. Chen, C.-C. Chen, Z.-Y. Chang, J.-Y. Fang and S.-C. Yang, Front. Pharmacol., 2019, 10, 1445 CrossRef CAS PubMed.
- S. Miethbauer, F. Gaube, U. Möllmann, H.-M. Dahse, M. Schmidtke, M. Gareis, M. Pickhardt and B. Liebermann, Planta Med., 2009, 75, 1523–1525 CrossRef CAS PubMed.
- A. Schueffler and T. Anke, Nat. Prod. Rep., 2014, 31, 1425–1448 RSC.
- H. Hashizume, M. Igarashi, S. Hattori, M. Hori, M. Hamada and T. Takeuchi, J. Antibiot., 2001, 54, 1054–1059 CrossRef CAS PubMed.
- D. E. Kadouri and R. M. Shanks, Res. Microbiol., 2013, 164, 821–826 CrossRef CAS PubMed.
- P. H. Mygind, R. L. Fischer, K. M. Schnorr, M. T. Hansen, C. P. Sönksen, S. Ludvigsen, D. Raventós, S. Buskov, B. Christensen and L. De Maria, Nature, 2005, 437, 975–980 CrossRef CAS PubMed.
- P. Sarathi and G. Thilagavathi, J. Text. Appar. Technol. Manag., 2009, 6(2), 1–8 Search PubMed.
- V. Parthasarathi and G. Thilagavathi, Int. J. Pharm. Pharm. Sci., 2011, 3, 392–398 CAS.
- E. Malka, I. Perelshtein, A. Lipovsky, Y. Shalom, L. Naparstek, N. Perkas, T. Patick, R. Lubart, Y. Nitzan and E. Banin, Small, 2013, 9, 4069–4076 CrossRef CAS PubMed.
- Q. Ye, W. Chen, H. Huang, Y. Tang, W. Wang, F. Meng, H. Wang and Y. Zheng, Appl. Microbiol. Biotechnol., 2020, 104, 5213–5227 CrossRef CAS PubMed.
- N. C. Cady, J. L. Behnke and A. D. Strickland, Adv. Funct. Mater., 2011, 21, 2506–2514 CrossRef CAS.
- W. Y. Chen, H. Y. Chang, J. K. Lu, Y. C. Huang, S. G. Harroun, Y. T. Tseng, Y. J. Li, C. C. Huang and H. T. Chang, Adv. Funct. Mater., 2015, 25, 7189–7199 CrossRef CAS.
- N. Rabiee, S. Ahmadi, O. Akhavan and R. Luque, Materials, 2022, 15, 1799 CrossRef CAS PubMed.
- M. K. Rai, S. Deshmukh, A. Ingle and A. Gade, J. Appl. Microbiol., 2012, 112, 841–852 CrossRef CAS PubMed.
- S. Roy Choudhury, S. Roy, A. Goswami and S. Basu, J. Antimicrob. Chemother., 2012, 67, 1134–1137 CrossRef CAS PubMed.
- G. R. Gibson and M. B. Roberfroid, J. Nutr., 1995, 125, 1401–1412 CrossRef CAS PubMed.
- P. Markowiak and K. Śliżewska, Nutrients, 2017, 9, 1021 CrossRef PubMed.
- R. Patel and H. L. DuPont, Clin. Infect. Dis., 2015, 60, S108–S121 CrossRef PubMed.
- M. L. Marco, D. Heeney, S. Binda, C. J. Cifelli, P. D. Cotter, B. Foligné, M. Gänzle, R. Kort, G. Pasin and A. Pihlanto, Curr. Opin. Biotechnol., 2017, 44, 94–102 CrossRef CAS PubMed.
- K. Pandey, S. Naik and B. Vakil, J. Food Sci. Technol., 2015, 52, 7577–7587 CrossRef CAS PubMed.
- S. Huang and W. H. Chang, Curr. Drug Metab., 2009, 10, 905–913 CrossRef CAS PubMed.
Footnote |
| † These authors contributed equally contribution. |
| This journal is © The Royal Society of Chemistry 2022 |