
Recent advances in spheroid-based microfluidic models to mimic the tumour microenvironment
Jooyoung
Ro
ab,
Junyoung
Kim
a and
Yoon-Kyoung
Cho
 *ab
*ab
aDepartment of Biomedical Engineering, Ulsan National Institute of Science and Technology (UNIST), Ulsan, Korea. E-mail: ykcho@unist.ac.kr
bCenter for Soft and Living Matter, Institute for Basic Science (IBS), Ulsan, Korea
First published on 21st April 2022
Abstract
Three-dimensional (3D) multicellular spheroid models can recapitulate the human tumour microenvironment with more accuracy than conventional cell culture models, as they include complex architectural structures and dynamic cellular interactions. Among the diverse platforms for spheroid formation, microfluidic platforms have been extensively applied to study spheroids because they can mimic the in vivo microenvironment. This review provides an overview of the advantages of 3D spheroid cultures with a summary of the recent applications for tumour microenvironment-focused cellular interactions, as well as the studies on spheroids and external stimuli. These 3D tumour spheroid-based microfluidic devices will provide a platform for a better understanding of cellular and external interactions, as well as the discovery of cancer therapeutics.
Introduction
As model platforms for human cancer and other diseases, two-dimensional (2D) monolayer in vitro and in vivo models are presently used for drug screening, basic biology studies, and tissue engineering.1 Although 2D monolayer models are more cost-effective and convenient, they cannot mimic the complex physiological environment of the human body.2In vivo animal studies can emulate actual physiological conditions better than 2D cultures. However, they still have several drawbacks, such as ethical concerns, cost issues, and different immune results compared to those for humans.3 Therefore, neither approach is ideal for modelling diseases and routine applications in therapeutic studies. Considering the importance of the tumour microenvironment in creating tumour models in vitro, three-dimensional (3D) cell culture has emerged as an alternative. 3D cultures can provide a physiologically relevant microenvironment compared to 2D cultures and are more cost-effective compared to in vivo cultures, with no associated ethical issues. Thus, the use of 3D tumour models could help overcome the limitations of 2D cultures and in vivo experiments while providing results consistent with those of in vivo studies (Fig. 1).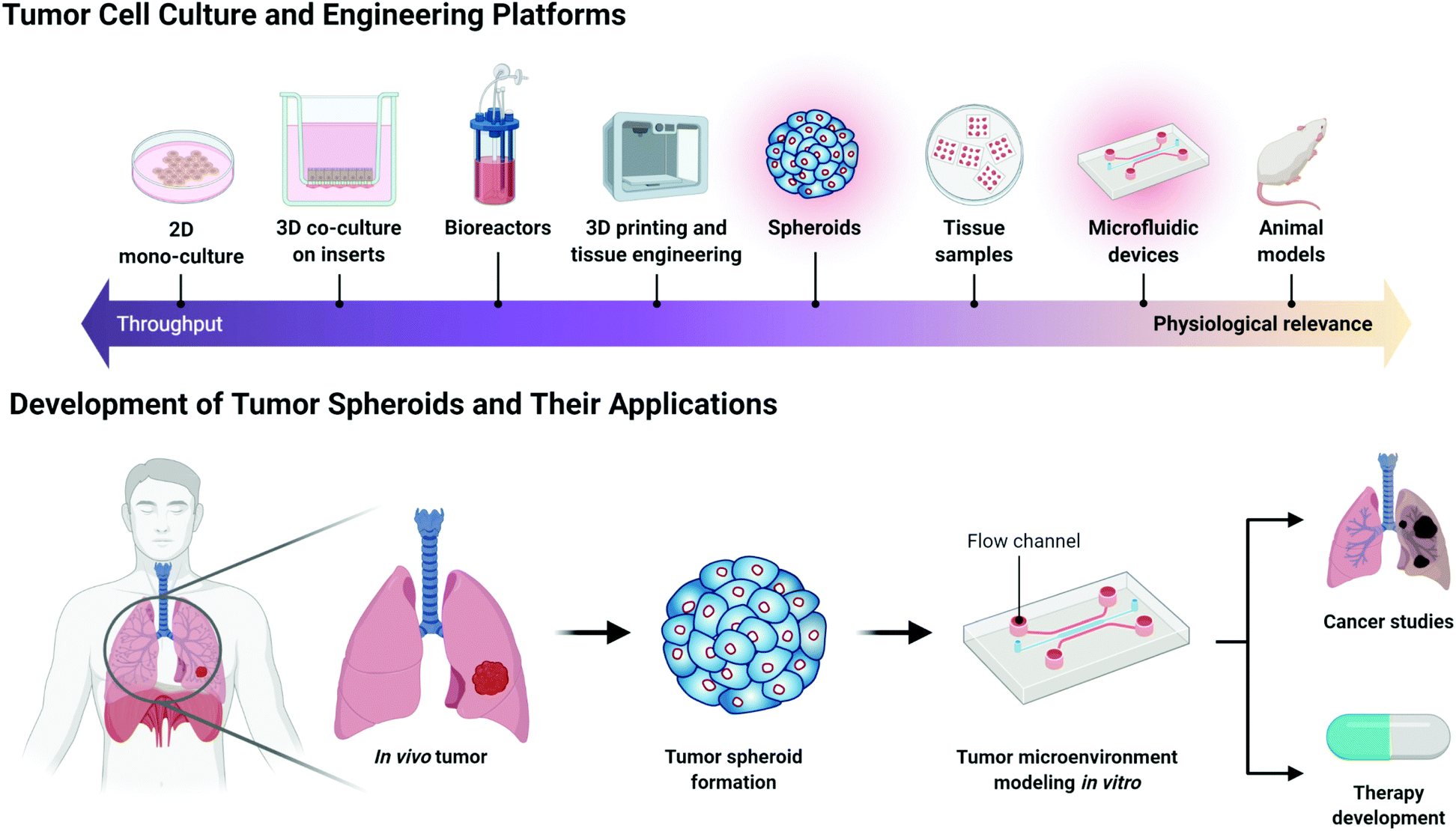 | ||
| Fig. 1 Scheme of tumour cell engineering platforms and the development of tumour spheroids and their applications. Various methods for modelling tumour cell cultures are introduced depending on their throughput and physiological relevance (top). Overall introduction to the development of tumour spheroids including what they look like, where they can be cultured, and how they can be used as an in vitro study model (bottom). | ||
3D in vitro culture models have been widely used in tissue engineering, biomedical engineering, disease studies, and drug discovery.4 Spheroids are cellular aggregates with a small, tightly-bound spherical shape grown in suspension or embedded in a 3D matrix using 3D culture methods.5 Cancer cell spheroids, also known as multicellular tumour spheroids, exhibit avascular tumour nodules or micrometastasis.6 3D tumour spheroids can mimic cell–cell and cell–matrix interactions between tumour cells and the tumour microenvironment, as well as physiological transport characteristics. Larger-sized spheroids are able to sustain oxygen and nutrient gradients, leading to the formation of a necrotic core that can be found in poorly vascularised tumours in vivo.7 Moreover, 3D tumour spheroids better resemble clinical tumours in terms of protein and gene expression profiles compared to 2D culture models.8
Several 3D cell culture methods have been developed for the formation of 3D spheroids. The most suitable technique can be chosen depending on the type of cell, tissue origin, aim of the study, and need for high-throughput capabilities. Several culture tools are available for 3D spheroid formation, including bioreactors, gel-embedded systems, hanging-drops, droplet reactors, microfluidics, and 3D bioprinting.9 Along with the 3D cell culture techniques, well-established techniques must be adapted to thoroughly characterise these complex cellular aggregates. Such characterisation is required to extract relevant biological data and standardise techniques and protocols for spheroid validation in preclinical assays.
This article reviews the 3D spheroid formation methods and highlights their applications in studying cell–cell interactions and external biophysical interactions in the tumour microenvironment. Furthermore, the review also discusses the limitations of the current spheroid-based in vitro microfluidic models and proposes future research opportunities.
Spheroid formation platforms
Bioreactors
Bioreactors are widely used to prepare spheroids with precise control and reproducibility (Fig. 2A). Cells with optimal density are placed in the bioreactor chamber under continuous agitation using a pump system through a scaffold. Bioreactors are equipped with systems to provide efficient nutrient circulation, waste removal, and physical and chemical homogeneity.10 Bioreactor spheroid formation models are more suited for large-scale spheroid production at an industrial level than other spheroid formation methods.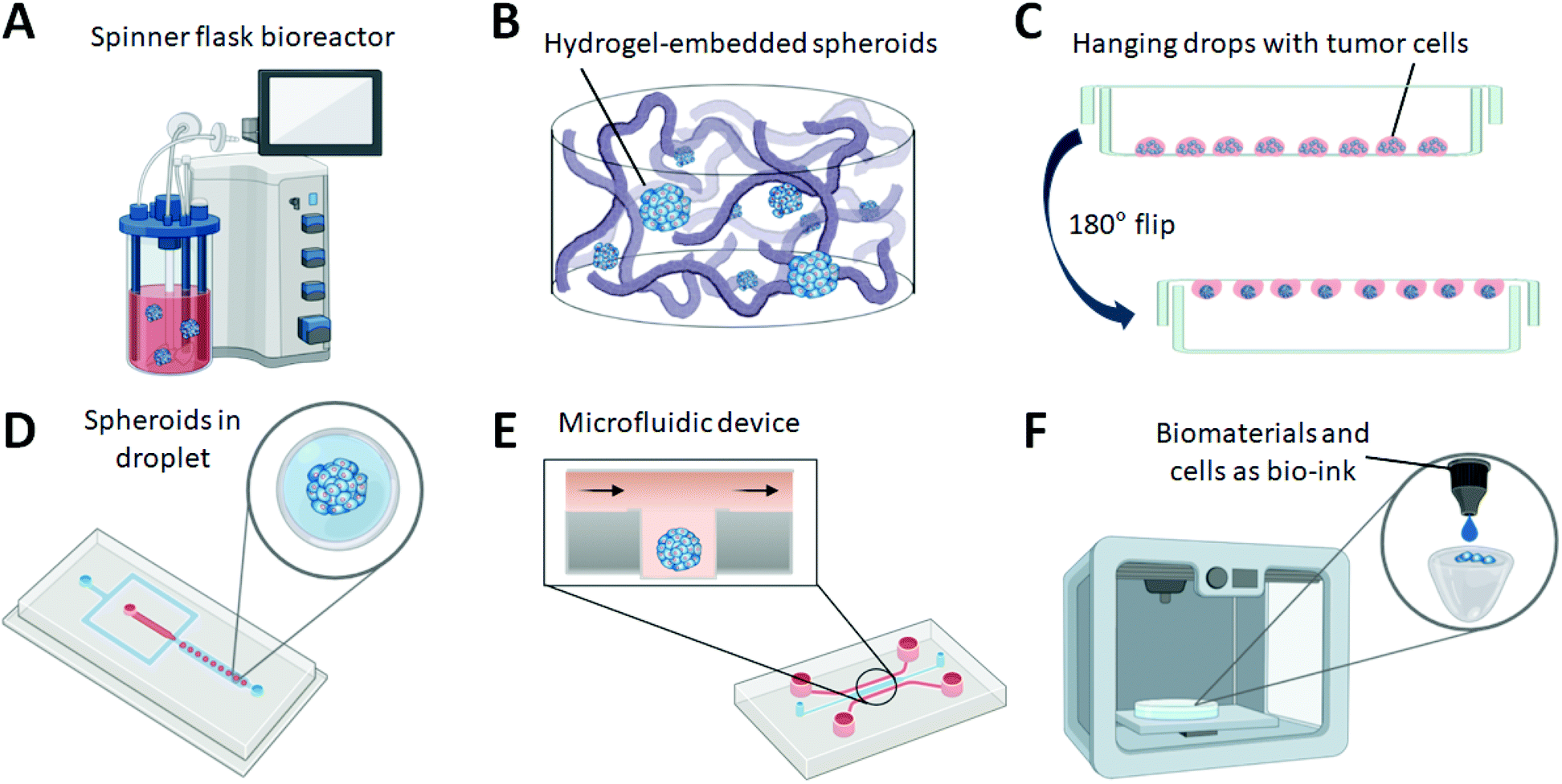 | ||
| Fig. 2 Different formation platforms for 3D spheroids. (A) Spinner flask bioreactor, (B) hydrogel-embedded spheroids, (C) hanging drops, (D) droplet generation and cell encapsulation, (E) microfluidic device, and (F) 3D bioprinting. | ||
Massai et al. developed a versatile bioreactor designed for a dynamic suspension cell culture to study the effect of shear stress conditions on tumour spheroids.11 They were able to tune the shear stress of the bioreactor and demonstrated that the ultralow shear suspension provided by the bioreactor was beneficial for spheroid culturing. Additionally, Zhang et al. used a microgravity bioreactor for developing biomaterial-free-mediated spheroid formation to maintain the properties of adipose-derived stem cells.12 They were able to form spheroids with enhanced stemness properties using only the microgravity bioreactor, without any additional biomaterials. For encapsulation of human pluripotent stem cells (hPSCs) and formation of spheroids in scale-up, Fattahi et al. developed a bioreactor model where microcapsules shield cell spheroids against shear stress in the bioreactor.13 The use of hydrogel microcapsules can prevent damage to the cells compared with bare cells in the bioreactor. Moreover, the encapsulation did not affect the pluripotency state and differential potential of the hPSC spheroids.
Bioreactors are advantageous because they can produce a large number of heterogeneous spheroids; however, the produced spheroids are randomly distributed in size and cell population, which requires manual selection steps for spheroid placement onto a dish.
Gel-embedded platforms
The formation of spheroids in gel-embedded structures is a widely used method (Fig. 2B). Hydrogels are commonly used for cell culture because they can mimic the elements of the native extracellular matrix (ECM), support cell adhesion, and exhibit mechanical properties similar to those of soft tissues.14 The protein-based biocompatible hydrogels, such as collagen, Matrigel, or fibrin, are most widely used for cell culture.15 The gel-embedded spheroids are suitable for studying chemotaxis and the metastatic behaviour of spheroids, as well as the co-culture of stromal cells with spheroids.16Charoen et al. described a simple method for embedding spheroids within collagen gels followed by whole spheroid and single-cell analysis to evaluate the efficacy of anticancer therapeutics.17 Spheroids were externally grown in agarose-coated well plates and seeded into collagen gels. As the collagen-based ECM structure surrounds the spheroids, spheroids respond differently to chemotherapeutics and drug delivery than 2D monolayer cultures, mirroring the results observed in a murine xenograft model. Similarly, Shin et al. reported a microwell platform fabricated in a hydrogel scaffold for studying chemotherapy using Matrigel-embedded 3D spheroids.18 Uniform-sized spheroids were grown in the microwells of the hydrogel scaffold and directly used to study the therapeutic efficiency of an anticancer drug. Morello et al. developed a thermo-sensitive, natural polymer-based hydrogel consisting of chitosan and pectin. The hydrogel exists in a gel state at room temperature and in a liquid state at higher temperatures, and supports the culture of tumour spheroids up to 44 days of culture.19
These findings highlight the benefits of gel-embedded spheroid formation, as covalently cross-linked hydrogels have enabled advanced cancer research, providing a natural tumour microenvironment for spheroid cultures. However, gel-embedded spheroids have limitations such as high cost and the need for manual operation.
Hanging drop plates
The hanging drop method for producing spheroids provides simplicity and reproducibility in generating uniform-sized spheroids (Fig. 2C). In this method, cells are placed in a hanging drop culture plate and incubated until the cells form 3D spheroids.20 As the hanging drop method requires relatively less specialised equipment compared with other spheroid culture methods and can be adapted to general culture conditions with high throughput, it is preferred as a simple culture method for the formation of spheroids.21While most hanging drop culture plates are made of plastic materials, Michael et al. demonstrated a hanging drop system for spheroid culture using surface-engineered paper.22 They utilised the porous properties of papers, combined with wax-patterned microfluidic channel networks, and performed in situ analyses such as drug testing, time-dependent detection of secreted proteins, and fluorescence staining without disturbing the spheroids. Zhao et al. developed a 3D printed hanging drop dripper for efficient spheroid culture and analysis without retrieval.23 Utilising a benchtop 3D printer, they printed an artefact that enables hanging drop formation and avoids the tedious fabrication process based on micromechanical systems. This system allows the long-term culture of spheroids and subsequent analysis as the spheroid drips from the array, eliminating the need for retrieval.
Liu et al. developed a polydimethylsiloxane (PDMS) platform for spheroid formation, culture, and drug response analysis.24 The platform was developed by punching holes on a 3 mm-thick PDMS layer with a flat needle. Due to PDMS deformation, the punched holes had wide ends and a narrow middle. Droplets were formed through the microholes, and tumour spheroids were cultured in the droplets.
Cui et al. applied the hanging drop method to develop a droplet microarray.25 This platform is based on a droplet fusion technique, where individual cell spheroids are merged into multicellular architectures. Fused multi-spheroids developed cell–cell interactions at the interface of the individual spheroids and evolved into single 3D cell structures over time. The complex spheroids formed with the fusion method can be advantageous for studying cellular interactions and signalling processes between the spheroids.
Although the hanging drop culture method is convenient and straightforward, it suffers from spheroid instability. A slight disruption of the droplet can affect the formation and structure of the spheroids, and maintaining the droplets for an extended period is difficult.
Droplet reactors
Spheroids can also be formed when encapsulated within single droplets prepared by single- and multi-phase flow systems using aqueous hydrogel solutions and oil phases (Fig. 2D). In this method, spheroids are first encapsulated in an aqueous hydrogel solution that flows into the oil phase, contacts calcium ions, and polymerises.26 The precursor hydrogel solution is used as the inner phase to produce spheroid-encapsulated microgels, contributing to structural support for 3D spheroid formation.27Sart et al. developed a droplet-based microfluidic platform in which the 3D spheroid culture, controlled stimulation, and imaging are all available in a single chip.28 Liquid agarose droplets containing cells were loaded into the microfluidic device for spheroid formation and gelation, followed by further testing with external stimuli and imaging. Lee et al. also reported a microfluidic system that used a droplet-based method for spheroid culture.29 To generate the homogenous spheroids, they controlled droplet generation frequency. This device can easily be automated, allowing minimal handling of spheroids. In addition, they could analyse the photothermal therapy of 3D spheroids and drug screening on a single device.
Chen et al. developed an acoustic droplet printing technique to form 3D tissues.30 When two leaky surface acoustic waves converge at the liquid–air interface and interfere, they result in an intense acoustic pressure profile, which overcomes the surface tension of the sample to drop off the droplet containing cells at the end of the jet. This method takes advantage of gentle acoustic force to minimize pressure on the cells. Xia et al. applied the concept to fabricate an acoustic drop-assisted microarray platform for the culture and screening of 3D cell spheroids.31 Initially, the spheroids were formed using hanging drop method, and drugs were applied to the spheroids through the acoustic droplet ejection technology for drug response testing.
As an application of the droplet technique, McMillan et al. developed droplets enveloped in a biocompatible and non-adherent oil–surfactant (polyethylglycol) interface.32 Droplets containing a specific number of cells were formed and dispensed into an oil–surfactant mixture; as the droplets floated on the oil layer, they formed a spherical medium–oil interface. Because the polyethylglycol-surfactant is biocompatible and non-adherent, the medium–oil interface provided a suitable condition for spheroid formation. This single emulsion method of forming spheroids enabled long-term culture of spheroids compared to the double emulsion methods for droplet-based spheroid formation.
Such droplet-based methods are adequate for the spheroid formation and can be easily integrated with microfluidic technologies. However, sustaining the size and stability of the droplets is a major difficulty that must be overcome for the long-term culture of cell spheroids.
Microfluidic devices
3D spheroid culture using microfluidic devices is one of the most widely applied methods and allows replication of the dynamic tumour microenvironment (Fig. 2E). For the fabrication of microfluidic devices, PDMS is the most commonly used material owing to its ease of use, long history, gas permeability, transparency, and relatively low cost.33 However, other materials such as thermoplastics and glass are also used for large-scale production.34Hsiao et al. developed a two-layer microfluidic system to culture 3D multi-cellular spheroids of prostate cancers, osteoblasts, and endothelial cells.35 This microfluidic platform includes a side-chamber microchannel design, enabling the formation of uniformly-sized spheroids that are kept stationary during media exchange. They applied this platform to create cancer niche-like microenvironments for chemotherapeutic development. Frey et al. introduced a fluidically interconnected hanging drop network.36 Multiple spheroids were cultured in each drop, while the fluidic network enabled continuous nutrient supply and intercellular metabolic communication. It connected parallel-formed spheroids on the same chip for complex multi-tissue experiments, a promising technology for research related to body-on-a-chip.
Petreus et al. developed a platform that mimics the pharmacokinetic profile of compounds in tumour spheroids.37 This platform can mimic various physiological scenarios of flow rates by controlling the tilting angle of the device, corresponding to the physiological relevant pharmacokinetic profile. Prince et al. developed a device that is able to form tumour spheroids uniformly under close-to-physiological flow conditions within a microfluidic device.38 The use of flow in this device provides longer culture durations of spheroid culture, model of dynamic drug conditions, and sequential testing of drugs, mimicking clinical pharmacokinetics.
A microfluidic platform with U-shaped barriers was developed by Barisam et al. to study the influence of the environmental conditions on in vitro cultures.39 The U-shaped structures trap the cells and form spheroids, while the microfluidic channel surrounds the spheroids. Using this platform, the research team studied shear stress, hypoxia, glucose metabolism, and anoikis through gene expression from the spheroids. Dornhof et al. established a microfluidic device integrated with electrochemical chemo- and biosensor arrays for simultaneous monitoring of oxygen, lactate, and glucose.40 Depending on the sensor locations in the microfluidic chip, they provided values of different parameters, yielding valuable information for applying the platform in patient-specific chemotherapy for personalized medicine. Soft lithography using PDMS is the most frequently used methods for fabricating microfluidic devices for spheroids formation. However, Li et al. fabricated an open microfluidic device with polymethyl methacrylate and laser cutting.41 They were able to fabricate different liquid guiding rails using laser cutting, and the open microfluidic design of the device enabled direct medium and spheroid access. Therefore, this microfluidic model could provide versatile designs, and proposes propose further spheroid-related studies.
Gao et al. developed a microfluidic device with acoustic bubbles to isolate, collect, and culture spheroids with circulating tumour cells.42 In this device, flow rates were optimized to capture the cells and form spheroids inside the bubbles. Moreover, the formed spheroids were released into the sub-channels in the devices for further culture and observation.
Microfluidic-based 3D spheroids provide significant advantages when mimicking in vivo microenvironments and elucidating cell–cell interactions. However, this method still needs a reduction in manual handling and lower costs. Further improvements are also required to meet the standards for commercial and clinical use.
3D bioprinting
Recently, 3D printing technology has been widely applied in various fields, including the formation of spheroids.10 The 3D printing technique refers to the construction of customised 3D structures under computational control where materials are printed.43 For 3D bioprinting, cells are printed directly and assembled into an organised structure, followed by in vitro culture and detection of biological functions (Fig. 2F).44 Recently, researchers have also used microfluidic channels and chambers as receiving plates for 3D printing on a chip.Duarte Campos et al. used 3D bioprinting to capture the 3D heterogeneity of tissues.45 Elastin-like protein engineered hydrogels were used as bioinks for constructing tissue models, which can be directly dispensed onto endothelialised chip platforms, establishing functional tissue models. Skylar-Scott et al. utilised stem-cell-derived spheroids as organ building blocks, assembling these blocks into living matrices where perfusable vascular channels were introduced by 3D bioprinting.46 Spheroids as matrices provide the requisite cell density, function, and microarchitecture of native tissues. With this technology, perfusable organ-specific tissues can be created using embedded vascular channels for therapeutic applications.
The 3D bioprinting technique receives significant attention because it can generate geometric constructs containing viable cells with high reproducibility. However, despite these advantages, the main limitations are the requirement of an expensive 3D bioprinting machine and the possible damage to the cells during the printing process.
3D multicellular interaction in tumour microenvironment
Among the several existing spheroid formation and culturing methods, cell aggregation in microfluidic systems is the most widely used method for studying multicellular spheroids and their interactions. Microfluidic technology enables precise control of small fluid volumes and can be applied to numerous applications in diagnostics and chemotherapeutic studies.1 Also, as microfluidic platforms can mimic the in vivo microenvironment, many studies have been conducted to study the cell–cell and cell–matrix interactions among spheroids and vascular endothelial cells, fibroblasts, and hepatic stellate cells.Spheroids and endothelial cells
Kwak et al. developed a microfluidic system to study tumour angiogenesis via tumour spheroid interactions with perfused blood vessels (Fig. 3A).47 Within the microfluidic platform, they studied vascular sprouting and vessel lumen formation in engineered blood vessels in a variety of ECM, tumour spheroids, and tumour stromal cells to demonstrate the crosstalk between tumour spheroids and perfused blood vasculatures in vitro. Among the different hydrogels, fibrin ECM was used to embed endothelial cells, which exhibited active patterns related to tumour angiogenesis and migration. Additionally, spheroids containing fibroblasts within the microfluidic device optimally mirrored the tumour–vascular interface in terms of physiological environments and biochemical interactions. This microfluidic model can mimic the tumour microenvironment in vitro, facilitating detailed research on the tumour microenvironment. | ||
| Fig. 3 Interaction of tumour spheroids with other cell types within the tumour microenvironment in microfluidic platforms. (A) A microfluidic platform was developed to study the cellular interaction and angiogenesis between tumour spheroids and perfused blood vessels induced by endothelial cells.42 (B) Co-culture microfluidic platform for culturing tumour spheroids, fibroblasts, and macrophages. Behaviour of fibroblasts and macrophages co-cultured with tumour spheroids is studied within the platform.51 (C) Microfluidic system recapitulating the tumour-immune microenvironment, investigating the effects of neutrophils on invasion of tumour spheroids.54 NETs induce spheroid distortion, correlating with the density of NETs in the stromal collagen region. (D) Microchannel plate-based system for co-culture of tumour spheroids and hepatic stellate cells.57 Activation of the hepatic stellate cells was observed with the changes in morphology (F-actin) and α-SMA expression levels, a marker of activated stellate cells. Panel A is adapted with permission from ref. 42. Copyright 2020 Springer Nature. Panel B is adapted with permission from ref. 51. Copyright 2021 MDPI. Panel C is adapted with permission from ref. 54. Copyright 2021 Institute of Physics. Panel D is adapted with permission from ref. 57. Copyright 2019 Royal Society of Chemistry. | ||
Similarly, Nashimoto et al. focused on vessel perfusion and the corresponding effects on tumour spheroids.48 They developed a tumour-on-a-chip platform that enables the evaluation of tumour activities in a microfluidic tumour vascular network. The spheroids were co-cultured with fibroblasts to induce angiogenic sprouts, establishing a perfusable vascular network in a tumour spheroid that replicated the tumour microenvironment. The perfusable endothelial network enabled long-term perfusion culture of tumour spheroids, provided oxygen and nutrition, and enabled drug administration under perfusion conditions. They compared static and perfusion conditions during drug administration, demonstrating the importance of perfusion as the effects of the drug were overcome by the media supply; additionally, dose-dependent effects were minimal compared with static conditions.
Ko et al. also applied the microfluidic system to model angiogenesis sprouting patterns and their cell–cell interactions.49 They used polystyrene to fabricate the microfluidic device, and the design was simple, incorporating a tapered hole in the centre to pattern a spheroid with various cell types and ECM. Fibroblast-laden hydrogels were located at the centre rail channel, whereas human endothelial cells were located at the side of the centre rail channel for studying angiogenic sprouting. As the platform mediates open microfluidics, it allows the implementation of spontaneous fluid patterning with high repeatability and the modelling of spontaneous capillary flow. Using this platform, the research team established a variety of tumour-endothelial models, including a 3D perfusable blood vessel network, a spheroid-based invasion assay, and a spheroid-mediated angiogenesis model.
To study the biology of cell–cell interaction of angiogenesis with tumour spheroids, Lee et al. mimicked the in vivo tumour microenvironment on a microfluidic chip with tumour spheroids located in the microwells and the endothelial cell layer in the lumen of the channels.50 A pump was used to induce the flow of cell media consistently to the cells for long-term stability. As this flow generates oxygen and nutrient gradients, it can control the metabolic microenvironment to study the corresponding effect on spheroid structure and metastasis. They also studied the effects of matrix metalloproteinase (MMP-1) on the collagen matrix, which exerts proangiogenic functions, and observed shrinkage of the collagen matrix, which contributes to the formation of the self-organising 3D matrix surrounding the spheroids.
Shin et al. developed a platform to model the 3D vascular networks of the brain-on-a-chip.51 Induced neural stem cells (iNSC) were formed into spheroids and cultured with a vascular network. This model of iNSC spheroids and perfused blood vessels was formed in a shape where the vascular network was wrapped around the iNSC spheroid and performed perfusion, and the brain neural stem cells were in direct contact with the blood vessels in vivo. Overall, the co-culture model enhanced differentiation and reduced apoptosis compared with monolayer culture. This platform represents the neurovascular system in vitro, which can increase the understanding of brain homeostasis and pathological conditions.
Spheroids and fibroblasts
Jeong et al. developed a microfluidic model to replicate the tumour microenvironment with spheroids and cancer-associated fibroblasts (CAFs) within a hydrogel scaffold.52 Interactions between tumour spheroids and fibroblasts contribute to the initiation, progression, and metastasis of tumours, while paracrine signalling of fibroblasts and tumour cells develops a mutual stimulation of cell proliferation and drug resistance in tumours. Within the microfluidic chip, the research team mimicked a microenvironmental condition that could recapitulate the interactions between tumour spheroids and fibroblasts without direct contact. Soluble factors are transported through the medium channel. Co-culture of tumour spheroids and fibroblasts induced changes in the morphology and secretion factors of fibroblasts, as confirmed by the growth rate, drug uptake, drug sensitivity, and ECM expression, which increased migration and activation of fibroblasts. With this model, they demonstrated that the 3D architecture of tumours and fibroblasts allows simultaneous interactions among the cells and increases the in vivo resemblance.To investigate the effect of CAFs on cancer cell invasion, Liu et al. developed a microfluidic-based co-culture device to model the in vitro tumour microenvironment.53 This platform consisted of six co-culture units, while communication between the salivary gland adenoid cystic carcinoma (ACC) spheroids and fibroblasts was established by the culture medium diffused into the matrix. The ACC spheroids nested in ECM mimicked the in vivo microenvironment in which tumour spheroids were formed, further invading the adjacent matrix when co-cultured with cancer-induced fibroblasts. The research team also demonstrated that an MMP inhibitor suppressed fibroblast-promoted spheroid invasion, confirming the suitability of the microfluidic platform for elucidating the tumour microenvironment.
Bender et al. established a digital microfluidic platform that enables the formation of tumour spheroids, encapsulation of spheroids in collagen, and exposure of spheroids to migration-modulating agents within a single platform, and is capable of automating the liquid handling steps.54 Applying this liquid handling technique, the research team developed a physiologically relevant invasion model with human colorectal adenocarcinoma spheroids and fibroblasts, modelling the tumour–stromal communication system in the in vitro microenvironment. In this microfluidic system, human colon carcinoma cells and fibroblasts are both formed into spheroids, and secretions from carcinoma spheroids are delivered to fibroblast spheroids, mimicking paracrine signalling. Encapsulating the spheroids in an automated platform within an ECM matrix allows the modelling of cell–ECM interactions and signalling with ease and greater accessibility.
Hockemeyer et al. developed a microfluidic device suitable for studying the cellular interactions between stromal cells and tumour spheroids in the 3D microenvironment.55 Through this platform, they co-cultured tumour spheroids, endothelial cells, and fibroblasts in a microenvironment that recapitulated several cell parameters in vivo. Although fibroblasts and endothelial cells were co-cultured, fibroblasts were studied, and the effects of normal tissue-associated fibroblasts (NAFs) were compared with those of CAFs in tumour-spheroid sprouting and secretion of stromal factors. The NAFs exhibited minimal sprouting, while the addition of stromal cell-derived factor-1 (SDF-1) and CAFs upregulated tumour cell sprouting from the spheroids. Thus, this microfluidic device can be used to evaluate the characteristics of stromal cells in tumour spheroids and to assess the chemotherapeutic effects of anticancer drugs on tumour growth, migration, and apoptosis.
To study the pancreatic ductal adenocarcinoma (PDAC) in vitro, Jang et al. established a culture model on a microfluidic platform consisting of PDAC tumour spheroids, cancer associated fibroblasts, and tumour-associated macrophages (Fig. 3B).56 With the microfluidic model, the research team studied the epithelial–mesenchymal transition (EMT)-induced effects, fibronectin remodelling around the tumour spheroids, increased fibronectin alignment, thickness, and degradation of cancer cells in co-culture conditions with fibroblasts and macrophages. Therefore, this microfluidic culture model of PDAC is effective in studying the activity of anticancer agents against cancer and stromal cells.
Spheroids and immune cells
Ayuso et al. developed a microfluidic model to study natural killer (NK) cell responses to solid tumours.57 In this model, breast cancer spheroids were formed in the ECM matrix, lumens with endothelial and NK cells, and antibodies embedded in the matrix or perfused through lateral blood vessels. The research team studied the movements of NK cells through tumour cell–cell junctions and the cytotoxicity of NK cells within solid tumours. Through this platform, they found that NK cells can detect the presence of tumours several hundred microns away. Additionally, the capability of NK cells within the tumour spheroid to kill the cells was studied, which suggested that NK cell exhaustion is a slow process. This microfluidic system provides a platform for co-culturing various cell types to study the complex cancer immunity process.A microfluidic system recapitulating the tumour microenvironment by integrating tumour spheroids and human monocyte-derived macrophages within a hydrogel scaffold in close co-culture with endothelial cells was developed by Bai et al.58 The platform mimicking the microenvironment was utilised to study the individual subtypes of macrophages in human lung cancer spheroids. Among the macrophage subtypes, M1 and M2b macrophages exhibited the strongest ability to induce EMT, regardless of the distance from the tumour spheroids. However, direct contact between the M2a macrophages and tumour spheroid induced dispersion of the cells from the spheroids. This indicates that M2a macrophages respond to some soluble factors from tumour spheroids, such as colony stimulating factor-1 (CSF-1) and chemokine (C-C motif) ligand 2 (CCL2), which recruit monocytes and promote differentiation to macrophages in the tumour microenvironment. In these microfluidic devices, different hematopoietic cells in the tumour microenvironment can also be studied for immunotherapeutic purposes.
Surendran et al. studied the interactions of neutrophils and tumour spheroids through chemotaxis and generation of neutrophil extracellular traps (NETs) on a 3D bioprinted microfluidic system fabricated on a porous membrane with neutrophils embedded in a collagen matrix (Fig. 3C).59 Activated neutrophils release NETs, forming protein web structures with cytotoxic enzymes as a host defence mechanism. NET formation stimulated the tumour cells from the aggregated state to the invasion state, where they invaded the surrounding collagen matrix. When the number of neutrophils increased, the NET structure in the collagen layer also increased as well, resulting in a higher spheroid distortion.
Spheroids and hepatic stellate cells
Hepatic stellate cells (HSCs) contribute to liver damage during inflammation or mechanical stimulation and are associated with disease progression and chemoresistance.60 HSCs are one of the typical co-culture components of hepatocytes as they are natural companions of hepatocytes in vivo and play a significant role in liver function and regeneration.Lee et al. used microfluidics to study cell–cell interactions between hepatocyte spheroids and hepatic stellate cells along with their paracrine effects.61 They developed a liver-on-a-chip in which primary 3D hepatocyte spheroids and HSCs were co-cultured without any cell–cell contact. In the platform, an osmotic pumping system was used for continuous flow of the medium to the cells, requiring minimal handling and no external power source. Using this platform, they studied the paracrine effects of HSCs on hepatocyte spheroids, demonstrating the cell–cell interaction of HSCs and hepatocyte spheroids in the formation of tight cell–cell contacts, thus improving the liver-specific function of the microfluidic chip.
Chen et al. developed a microchannel plate-based culture model consisting of concave microwells for co-culturing tumour spheroids with stellate cells (Fig. 3D).62 In this model, the different cell lines were in direct contact with each other, providing cancer cell–stellate interaction data under drug stimulation. The microchannel plate is able to form concentration gradients of the treated drugs, mimicking the in vivo tumour microenvironmental interactions. The activated stellate cells expressed EMT-related factors and drug resistance in cancer cells. This platform can be further utilized to study the drug resistance induced by microenvironmental factors.
A liver-on-a-chip was developed by Meng et al., mimicking the mass transfer and structure of the hepatic lobule with hepatocellular carcinoma HepG2 cells and human hepatic stellate cell LX-2 to form spheroids with HUVEC cells in the inner channel to mimic the capillary vessel.63 Nutrients for the hepatic spheroids were delivered through the inner channel, as the cells are able to interact and exchange nutrients and metabolites through the permeable hydrogel. The cell culture medium was perfused through the endothelial channel through physiologically relevant interstitial flow rates. Compared with static culture, the hepatic spheroids showed higher performance in liver-specific functions, such as albumin and urea secretion. Hepatic spheroids and shear flow studies on the liver-mimicking chip indicate the importance of mass transfer in replicating cellular functions of the liver in vitro.
External stimuli in tumour microenvironment
As 3D spheroids in microfluidic devices can mimic the human microenvironment, researchers have applied these platforms to study external biophysical interactions, such as interstitial flow, fluid dynamics, and hypoxia, between spheroids and biophysical cues.Spheroids and interstitial flow
A physiologically-realistic microfluidic platform using tumour spheroids to model the interstitial flow present in in vivo tissues was developed by Huang et al. (Fig. 4A).64 Interstitial flow is the movement of fluids within the interstitial space, a critical component for maintaining homeostasis in healthy tissues and removing metabolic waste from tissues. Moreover, interstitial flow in the tumour microenvironment can impact tumour invasion or the direct application of external stress to cells. The research team applied a 3D microfluidic platform with tumour spheroids, which were embedded within 3D collagen matrices, and studied the effect of interstitial flow on cell–cell adhesion within spheroids during tumour invasion. Through this platform, they demonstrated that interstitial flow promoted tumour invasion by downregulating E-cadherin in neighbouring non-tumourigenic cells, highlighting the importance of external biophysical forces. | ||
| Fig. 4 Multicellular tumour spheroids cultured in external-stimuli-controlled tumour microenvironment. (A) Microfluidic device modelling interstitial flow in vivo, generating interstitial-flow-dependent tumour spheroid modifications.59 (B) Microfluidic platform developed to control the concentration of oxygen in the tumour microenvironment, inducing different oxygen concentrations to tumour spheroids and promote different cell behaviour.62 (C) A microfluidic system able to induce different hydrogel stiffness in which spheroids proliferate, demonstrating higher proliferation in spheroids grown in stiff hydrogels.65 (D) Metabolic gradient induced in a microfluidic device, comparing cellular uptake of metabolic gradients in tumour spheroids and monolayer-culture cells.67 Panel A is adapted with permission from ref. 59. Copyright 2020 Springer Nature. Panel B is adapted with permission from ref. 62. Copyright 2019 Springer Nature. Panel C is adapted with permission from ref. 65. Copyright 2020 Elsevier B.V. Panel D is adapted with permission from ref. 67. Copyright 2021 Springer Nature. | ||
The significance of fluid dynamics on the culture of 3D spheroids and tissue maturation by developing a perfusable microfluidic platform with computational fluid dynamics-based simulation was demonstrated by Lopa et al.65 A perfusion system combined with digital systems allowed the generation of a model to predict cell trapping efficacy, flow rate, and seeding time, demonstrating the possibility of controlling the size and shape of spheroids through fluid flow. The effect of shear stress on spheroid formation was also evaluated; lower shear stress to the cells resulted in larger and less rounded spheroids, whereas higher shear stress resulted in smaller and spherical spheroids. This phenomenon was attributed to the fluid dynamics stimulation of cells, which minimised the shear stress on the surface of the cellular aggregates. This platform provides the advantage of obtaining different forms of spheroids without manipulating the geometry of the chip by tuning non-geometrical parameters. This system holds great potential as a microfluidic platform, as it can be used to develop in vitro models for studying physiological tissue maturation mechanisms.
Collins et al. developed a microfluidic device for a better understanding of the relationship between interstitial flow and cancer metastasis.66 In this device, only a single spheroid from different cell lines was exposed to continuous perfusion culture at a flow rate relevant to tumour interstitial fluid flow in vivo. The spheroids were captured in a well inside through an access port on the device, and flow was provided through the microchannel. The impact of flow on metastatic characteristics of the cancer models was investigated, including studies of metastatic factors, vascular endothelial growth factor (VEGF), and interleukin-6 (IL-6) cytokine levels in the cell culture media, stressing the importance of flow in the microenvironment.
Spheroids and hypoxic environment
Grist et al. applied the microfluidic platform to study tumour spheroid response to hypoxic and normoxic conditions (Fig. 4B).67 They developed a platform capable of recreating physiologically relevant cycling oxygen levels that are not attainable in traditional cell culture environments. Using this microfluidic device, they monitored spheroids during hypoxic exposure and observed that spheroids swell and shrink in response to oxygen concentrations over time; such swelling behaviour was attributed to the individual cells forming the spheroid. Additionally, the device was used to monitor tumour spheroids during drug treatment under chronic and cycling hypoxia as well as normoxia. As spheroids form a metabolic gradient from the spheroid surface to the core, controlling oxygen at the surface modulates the dynamics of the oxygen gradients, which can mimic in vivo oxygen levels within a tumour region adjacent to the perfused blood vessel. This provides a relevant tumour microenvironment for the development of drugs or studying tumour behaviour.A microfluidic device that is able to perform five different oxygen levels with a linear gradient in a tumour spheroid co-culture environment was developed by Fridman et al.68 The gas and tumour spheroid cultures were divided with a thin, gas permeable PDMS membrane. The oxygen gradients were established as they are able to reflect the concentrations in hypoxic tumour regions to ambient air. The device was used to monitor the effect of oxygen gradients on reactive oxygen species generation and the cytotoxicity of anti-tumour drugs.
Spheroids and stiffness
A microfluidic system that can control the mechanical properties of the surrounding microenvironment in which spheroids are cultured was developed by Lee and Cha.69 They used bioactive spherical microgels to support 3D cultures. The mechanical properties depend on the concentration of the polymer for gel formation. Breast cancer cells such as MCF7 were used to form spheroids, which were first encapsulated in gelatine droplets, followed by a photocrosslinking step to form microgels containing spheroids in a microfluidic device. Spheroid proliferation was dependent on the mechanical properties of the microgels, as spheroid cells proliferated faster in microgels with higher stiffness. When MCF7 cells were co-cultured with other cells (macrophages and fibroblasts) to form a spheroid, the cells exhibited higher proliferation regardless of gel stiffness. Lee and Cha studied the controlled mechanical properties of the microgels on tumour physiology over time (Fig. 4C).70 MCF7 cells generated spheroids grow in size over time, regardless of the higher rigidity of microgels, while MDA-MB-231 cells developed migratory polyploid giant cancer cells (PGCC) as they proliferated and formed spheroids only in higher rigidity. MCF7 and SK-BR-3 also formed PGCCs, but compared with MDA-MB-231, they tended to show smaller sizes but were quicker to form spheroids. Furthermore, the cytotoxic effects were influenced by the interactions between the cells and the mechanical microenvironment.Taubenberger et al. developed a physiologically relevant 3D microenvironment depending on the mechanical phenotype and stiffened hydrogels to mimic the stiffened stroma in vivo.71 Cancer cells are sensitive to the mechanical properties of the microenvironment, as they proliferate and push the surrounding stromal compartment, causing compressive stress on the cells. In this microfluidic device, the research team developed breast cancer spheroids in polyethylene glycol–heparin-based 3D microenvironments. To study the effect of microenvironment stiffness on the growth and mechanics of tumour spheroids, they tuned the stiffness of hydrogel cells and observed that stiff microenvironments have a negative influence on tumour spheroid growth. This coincided with a Rho-associated kinase (ROCK)-dependent increase, indicating that ROCK is a potential anticancer drug target. Applying this microfluidic platform to cancer environment studies can provide a better understanding of how cancer cells react to compressive stresses.
Spheroids and metabolic gradients
A study was presented by Patra et al., in which the metabolic activity of spheroids in a microfluidic device was further analysed by NMR spectroscopy as well as micro-NMR imaging (Fig. 4D).72 The research team cultured MCF7 spheroids in a microfluidic device that induced changes in the metabolic concentration in the culture medium for the spheroids. The concentrations of D-glucose, L-lactic acid, L-alanine, and L-glutamine were evaluated over time in single spheroids of different sizes compared to monolayer cultured cells. They found that spheroids consumed D-glucose and produced lactic acid at a 2.5 times slower rate than that of monolayer-culture cells. Monolayer cells produced L-alanine and L-glutamine, whereas spheroids exhibited slight consumption. A microfluidic system combined with NMR can provide a platform for studying spheroid metabolism.Conclusion and outlook
In this review, we focused on the advantages of 3D multicellular spheroid cultures, which can be used to overcome the limitations of 2D cell cultures and in vivo animal models. Multicellular spheroids exhibit complex cell signalling, ECMs, nutrient gradients, drug resistance, and metabolic adaptation, making spheroids a clinically relevant tool for cellular studies. The tumour microenvironment can be replicated more accurately in 3D models than in 2D models. Additionally, 3D culture models are simpler than in vivo models for standardisation and high-throughput production.Although spheroids hold great advantages for in vitro studies, there are still several challenges that must be overcome. First, spheroid production methods need to be optimised to reduce labour and the cost required to culture and study spheroids. Although spheroid culture is cheaper than in vivo experiments, it is not as cost-effective and convenient as 2D culture methods and requires much human labour. Thus, to simultaneously form many spheroids under controlled conditions, robust spheroid formation techniques need to be developed and optimised. Second, there is still a lack of technology for the simple and standardised analysis of spheroids during preclinical studies. Compared with 2D culture or animal models, 3D spheroid culture is a relatively new field of study, resulting in a limited amount of information on standardised assays that can be used for the analysis of spheroids applied in drug screening or clinical research. Finally, although 3D cultured spheroids are more physiologically relevant than 2D cultures, they still lack interaction with the immune system, full organ function, and multiple organ interactions. Recently, organoids have been established as a bridge that connects the existing gaps between spheroids and in vivo models. An organoid is a collection of organ-specific cell types developed from stem cells or organ progenitors that self-organise through cell sorting and spatially restricted lineage commitment in a manner similar to when in vivo.73 Spheroid and organoid models use different cell sources, culture methods, and establishment periods. However, organoids also benefit from the development of spheroid formation and experimental platforms as both are similar with 3D structures composed of multiple cells. Therefore, the advantages of the developed spheroid formation platforms could be utilized for the in vitro formation of organoids. Because ECM is required for the formation of organoid in vitro 3D culture with a cocktail of growth factors, the results of previous studies on spheroid growth in ECM can be utilized and developed for the culture of organoids, while maintaining long-term culture and viability of organoids using microfluidic technology.74 Recently, bioprinting has been applied for in vitro organoid construction, forming complex vascular systems, while reducing the size and increasing the yield of the organoids.75 Thus, the technology and knowledge gained from spheroid-based models can further advance the organoid models.
With further development of the spheroid culture platforms to overcome their limitations, we expect that the spheroid-based microfluidic platforms that mimic the complex tumour microenvironment will have broad applications, such as basic cancer research, drug screening, regenerative medicine, and tissue engineering.
Conflicts of interest
There are no conflicts to declare.Acknowledgements
Fig. 1 and 2 were created with Biorender.com. Fig. 1 was adapted from “Intestine Tissue Culture and Engineering Platforms” by BioRender.com (2022). This research was supported by funding from the Institute for Basic Science (No. IBS-R020-D1).Notes and references
- W. Asghar, R. El Assal, H. Shafiee, S. Pitteri, R. Paulmurugan and U. Demirci, Mater. Today, 2015, 18, 539–553 CrossRef CAS PubMed.
- B. W. Huang and J. Q. Gao, J. Controlled Release, 2018, 270, 246–259 CrossRef CAS PubMed.
- M. Kapalczynska, T. Kolenda, W. Przybyla, M. Zajaczkowska, A. Teresiak, V. Filas, M. Ibbs, R. Blizniak, L. Luczewski and K. Lamperska, Arch. Med. Sci., 2018, 14, 910–919 Search PubMed.
- C. Wang, Z. Tang, Y. Zhao, R. Yao, L. Li and W. Sun, Biofabrication, 2014, 6, 022001 CrossRef PubMed.
- G. F. Muschler, C. Nakamoto and L. G. Griffith, J. Bone Jt. Surg., Am. Vol., 2004, 86, 1541–1558 CrossRef PubMed.
- E. Fennema, N. Rivron, J. Rouwkema, C. van Blitterswijk and J. de Boer, Trends Biotechnol., 2013, 31, 108–115 CrossRef CAS PubMed.
- J. Friedrich, C. Seidel, R. Ebner and L. A. Kunz-Schughart, Nat. Protoc., 2009, 4, 309–324 CrossRef CAS PubMed.
- M. E. Katt, A. L. Placone, A. D. Wong, Z. S. Xu and P. C. Searson, Front. Bioeng. Biotechnol., 2016, 4, 12 Search PubMed.
- D. Liu, S. Chen and M. Win Naing, Biotechnol. Bioeng., 2021, 118, 542–554 CrossRef CAS PubMed.
- N. Chaicharoenaudomrung, P. Kunhorm and P. Noisa, World J. Stem Cells, 2019, 11, 1065–1083 CrossRef PubMed.
- D. Massai, G. Isu, D. Madeddu, G. Cerino, A. Falco, C. Frati, D. Gallo, M. A. Deriu, G. Falvo, D. Labate, F. Quaini, A. Audenino and U. Morbiducci, PLoS One, 2016, 11, e0154610 CrossRef PubMed.
- S. Zhang, P. Liu, L. Chen, Y. Wang, Z. Wang and B. Zhang, Biomaterials, 2015, 41, 15–25 CrossRef CAS PubMed.
- P. Fattahi, A. Rahimian, M. Q. Slama, K. Gwon, A. M. Gonzalez-Suarez, J. Wolf, H. Baskaran, C. D. Duffy, G. Stybayeva, Q. P. Peterson and A. Revzin, Sci. Rep., 2021, 11, 7177 CrossRef CAS PubMed.
- S. R. Caliari and J. A. Burdick, Nat. Methods, 2016, 13, 405–414 CrossRef CAS PubMed.
- N. E. Ryu, S. H. Lee and H. Park, Cells, 2019, 8, 1620 CrossRef CAS PubMed.
- Y. Li and E. Kumacheva, Sci. Adv., 2018, 4, eaas8998 CrossRef PubMed.
- K. M. Charoen, B. Fallica, Y. L. Colson, M. H. Zaman and M. W. Grinstaff, Biomaterials, 2014, 35, 2264–2271 CrossRef CAS PubMed.
- C. S. Shin, B. Kwak, B. Han and K. Park, Mol. Pharm., 2013, 10, 2167–2175 CrossRef CAS PubMed.
- G. Morello, A. Quarta, A. Gaballo, L. Moroni, G. Gigli, A. Polini and F. Gervaso, Carbohydr. Polym., 2021, 274, 118633 CrossRef CAS PubMed.
- R. Foty, J. Visualized Exp., 2011, 51, e2720 Search PubMed.
- N. E. Timmins and L. K. Nielsen, Methods Mol. Med., 2007, 140, 141–151 CAS.
- I. J. Michael, S. Kumar, J. M. Oh, D. Kim, J. Kim and Y.-K. Cho, ACS Appl. Mater. Interfaces, 2018, 10, 33839–33846 CrossRef CAS PubMed.
- L. Zhao, J. Xiu, Y. Liu, T. Zhang, W. Pan, X. Zheng and X. Zhang, Sci. Rep., 2019, 9, 19717 CrossRef CAS PubMed.
- X. Liu, H. Lin, J. Song, T. Zhang, X. Wang, X. Huang and C. Zheng, Micromachines, 2021, 12, 681 CrossRef PubMed.
- H. Cui, X. Wang, J. Wesslowski, T. Tronser, J. Rosenbauer, A. Schug, G. Davidson, A. A. Popova and P. A. Levkin, Adv. Mater., 2021, 33, e2006434 CrossRef PubMed.
- R. Vadivelu, H. Kamble, M. Shiddiky and N.-T. Nguyen, Micromachines, 2017, 8, 94 CrossRef.
- H. F. Chan, Y. Zhang, Y. P. Ho, Y. L. Chiu, Y. Jung and K. W. Leong, Sci. Rep., 2013, 3, 3462 CrossRef PubMed.
- S. Sart, R. F. Tomasi, G. Amselem and C. N. Baroud, Nat. Commun., 2017, 8, 469 CrossRef PubMed.
- J. M. Lee, J. W. Choi, C. D. Ahrberg, H. W. Choi, J. H. Ha, S. G. Mun, S. J. Mo and B. G. Chung, Microsyst. Nanoeng., 2020, 6, 1–10 CrossRef PubMed.
- K. Chen, E. Jiang, X. Wei, Y. Xia, Z. Wu, Z. Gong, Z. Shang and S. Guo, Lab Chip, 2021, 21, 1604–1612 RSC.
- Y. Xia, H. Chen, J. Li, H. Hu, Q. Qian, R. X. He, Z. Ding and S. S. Guo, ACS Appl. Mater. Interfaces, 2021, 13, 23489–23501 CrossRef CAS PubMed.
- K. S. McMillan, A. G. McCluskey, A. Sorensen, M. Boyd and M. Zagnoni, Analyst, 2016, 141, 100–110 RSC.
- K. Moshksayan, N. Kashaninejad, M. E. Warkiani, J. G. Lock, H. Moghadas, B. Firoozabadi, M. S. Saidi and N.-T. Nguyen, Sens. Actuators, B, 2018, 263, 151–176 CrossRef CAS.
- A. D. Castiaux, D. M. Spence and R. S. Martin, Anal. Methods, 2019, 11, 4220–4232 RSC.
- A. Y. Hsiao, Y. S. Torisawa, Y. C. Tung, S. Sud, R. S. Taichman, K. J. Pienta and S. Takayama, Biomaterials, 2009, 30, 3020–3027 CrossRef CAS PubMed.
- O. Frey, P. M. Misun, D. A. Fluri, J. G. Hengstler and A. Hierlemann, Nat. Commun., 2014, 5, 4250 CrossRef CAS PubMed.
- T. Petreus, E. Cadogan, G. Hughes, A. Smith, V. Pilla Reddy, A. Lau, M. J. O'Connor, S. Critchlow, M. Ashford and L. Oplustil O'Connor, Commun. Biol., 2021, 4, 1001 CrossRef CAS PubMed.
- E. Prince, S. Kheiri, Y. Wang, F. Xu, J. Cruickshank, V. Topolskaia, H. Tao, E. W. K. Young, A. P. McGuigan, D. W. Cescon and E. Kumacheva, Adv. Healthcare Mater., 2022, 11, e2101085 CrossRef PubMed.
- M. Barisam, F. R. Niavol, M. A. Kinj, M. S. Saidi, H. Ghanbarian and N. Kashaninejad, J. Sci.: Adv. Mater. Devices, 2022, 7, 100439 CAS.
- J. Dornhof, J. Kieninger, H. Muralidharan, J. Maurer, G. A. Urban and A. Weltin, Lab Chip, 2021, 22, 225–239 RSC.
- Q. Li, K. Niu, D. Wang, L. Xuan and X. Wang, Lab Chip, 2022, Advance Article Search PubMed.
- Y. Gao, M. Wu, Q. Luan, I. Papautsky and J. Xu, Lab Chip, 2022, 22, 805–813 RSC.
- J. Ma, Y. Wang and J. Liu, RSC Adv., 2018, 8, 21712–21727 RSC.
- S. Shrestha, V. K. R. Lekkala, P. Acharya, D. Siddhpura and M. Y. Lee, Essays Biochem., 2021, 65, 481–489 CrossRef PubMed.
- D. F. Duarte Campos, C. D. Lindsay, J. G. Roth, B. L. LeSavage, A. J. Seymour, B. A. Krajina, R. Ribeiro, P. F. Costa, A. Blaeser and S. C. Heilshorn, Front. Bioeng. Biotechnol., 2020, 8, 374 CrossRef PubMed.
- M. A. Skylar-Scott, S. G. M. Uzel, L. L. Nam, J. H. Ahrens, R. L. Truby, S. Damaraju and J. A. Lewis, Sci. Adv., 2019, 5, eaaw2459 CrossRef CAS PubMed.
- T. J. Kwak and E. Lee, Sci. Rep., 2020, 10, 20142 CrossRef CAS PubMed.
- Y. Nashimoto, R. Okada, S. Hanada, Y. Arima, K. Nishiyama, T. Miura and R. Yokokawa, Biomaterials, 2020, 229, 119547 CrossRef CAS PubMed.
- J. Ko, J. Ahn, S. Kim, Y. Lee, J. Lee, D. Park and N. L. Jeon, Lab Chip, 2019, 19, 2822–2833 RSC.
- S. W. Lee, S. Hong, B. Jung, S. Y. Jeong, J. H. Byeon, G. S. Jeong, J. Choi and C. Hwang, Biotechnol. Bioeng., 2019, 116, 3041–3052 CrossRef CAS PubMed.
- N. Shin, Y. Kim, J. Ko, S. W. Choi, S. Hyung, S. E. Lee, S. Park, J. Song, N. L. Jeon and K. S. Kang, Biotechnol. Bioeng., 2022, 119, 566–574 CrossRef CAS PubMed.
- S. Y. Jeong, J. H. Lee, Y. Shin, S. Chung and H. J. Kuh, PLoS One, 2016, 11, e0159013 CrossRef PubMed.
- T. Liu, B. Lin and J. Qin, Lab Chip, 2010, 10, 1671–1677 RSC.
- B. F. Bender, A. P. Aijian and R. L. Garrell, Lab Chip, 2016, 16, 1505–1513 RSC.
- K. Hockemeyer, C. Janetopoulos, A. Terekhov, W. Hofmeister, A. Vilgelm, L. Costa, J. P. Wikswo and A. Richmond, Biomicrofluidics, 2014, 8, 044105 CrossRef CAS PubMed.
- S. D. Jang, J. Song, H. A. Kim, C. N. Im, I. A. Khawar, J. K. Park and H. J. Kuh, Cancers, 2021, 13, 5955 CrossRef CAS PubMed.
- J. M. Ayuso, R. Truttschel, M. M. Gong, M. Humayun, M. Virumbrales-Munoz, R. Vitek, M. Felder, S. D. Gillies, P. Sondel, K. B. Wisinski, M. Patankar, D. J. Beebe and M. C. Skala, OncoImmunology, 2019, 8, 1553477 CrossRef PubMed.
- J. Bai, G. Adriani, T. M. Dang, T. Y. Tu, H. X. Penny, S. C. Wong, R. D. Kamm and J. P. Thiery, Oncotarget, 2015, 6, 25295–25307 CrossRef PubMed.
- V. Surendran, D. Rutledge, R. Colmon and A. Chandrasekaran, Biofabrication, 2021, 13, 035029 CrossRef CAS PubMed.
- D. Ezhilarasan, J. Cell Physiol., 2021, 237(1), 436–449 CrossRef PubMed.
- S. A. Lee, Y. No da, E. Kang, J. Ju, D. S. Kim and S. H. Lee, Lab Chip, 2013, 13, 3529–3537 RSC.
- Y. Chen, W. Sun, L. Kang, Y. Wang, M. Zhang, H. Zhang and P. Hu, Analyst, 2019, 144, 4233–4240 RSC.
- Q. Meng, Y. Wang, Y. Li and C. Shen, Biotechnol. Bioeng., 2021, 118, 612–621 CrossRef CAS PubMed.
- Y. L. Huang, Y. Ma, C. Wu, C. Shiau, J. E. Segall and M. Wu, Sci. Rep., 2020, 10, 9648 CrossRef PubMed.
- S. Lopa, F. Piraino, G. Talo, V. L. Mainardi, S. Bersini, M. Pierro, L. Zagra, M. Rasponi and M. Moretti, Front. Bioeng. Biotechnol., 2020, 8, 366 CrossRef PubMed.
- T. Collins, E. Pyne, M. Christensen, A. Iles, N. Pamme and I. M. Pires, Biomicrofluidics, 2021, 15, 044103 CrossRef CAS PubMed.
- S. M. Grist, S. S. Nasseri, L. Laplatine, J. C. Schmok, D. Yao, J. Hua, L. Chrostowski and K. C. Cheung, Sci. Rep., 2019, 9, 17782 CrossRef PubMed.
- I. B. Fridman, G. S. Ugolini, V. VanDelinder, S. Cohen and T. Konry, Biofabrication, 2021, 13, 035037 CrossRef CAS PubMed.
- D. Lee and C. Cha, Pharmaceutics, 2018, 10, 229 CrossRef CAS PubMed.
- D. Lee and C. Cha, Mater. Sci. Eng., C, 2020, 112, 110932 CrossRef CAS PubMed.
- A. V. Taubenberger, S. Girardo, N. Traber, E. Fischer-Friedrich, M. Krater, K. Wagner, T. Kurth, I. Richter, B. Haller, M. Binner, D. Hahn, U. Freudenberg, C. Werner and J. Guck, Adv. Biosyst., 2019, 3, e1900128 CrossRef PubMed.
- B. Patra, M. Sharma, W. Hale and M. Utz, Sci. Rep., 2021, 11, 53 CrossRef CAS PubMed.
- M. A. Lancaster and J. A. Knoblich, Science, 2014, 345, 1247125 CrossRef PubMed.
- S. Gunti, A. T. K. Hoke, K. P. Vu and N. R. London Jr., Cancers, 2021, 13, 874 CrossRef CAS PubMed.
- Y. Ren, X. Yang, Z. Ma, X. Sun, Y. Zhang, W. Li, H. Yang, L. Qiang, Z. Yang, Y. Liu, C. Deng, L. Zhou, T. Wang, J. Lin, T. Li, T. Wu and J. Wang, Int. J. Bioprint., 2021, 7, 364 CrossRef PubMed.
| This journal is © The Royal Society of Chemistry 2022 |