
 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a Creative Commons Attribution 3.0 Unported Licence
Success stories of natural product-derived compounds from plants as multidrug resistance modulators in microorganisms
Xiaohan Zhai†
a,
Guoyu Wu†a,
Xufeng Taoa,
Shilei Yanga,
Linlin Lva,
Yanna Zhua,
Deshi Dong *a and
Hong Xiang*b
*a and
Hong Xiang*b
aDepartment of Pharmacy, First Affiliated Hospital of Dalian Medical University, Dalian, China. E-mail: dongdeshi@dmu.edu.cn; xianghong@dmu.edu.cn
bLaboratory of Integrative Medicine, First Affiliated Hospital of Dalian Medical University, Dalian, China
First published on 8th March 2023
Abstract
Microorganisms evolve resistance to antibiotics as a function of evolution. Antibiotics have accelerated bacterial resistance through mutations and acquired resistance through a combination of factors. In some cases, multiple antibiotic-resistant determinants are encoded in these genes, immediately making the recipient organism a “superbug”. Current antimicrobials are no longer effective against infections caused by pathogens that have developed antimicrobial resistance (AMR), and the problem has become a crisis. Microorganisms that acquire resistance to chemotherapy (multidrug resistance) are a major obstacle for successful treatments. Pharmaceutical industries should be highly interested in natural product-derived compounds, as they offer new sources of chemical entities for the development of new drugs. Phytochemical research and recent experimental advances are discussed in this review in relation to the antimicrobial efficacy of selected natural product-derived compounds as well as details of synergistic mechanisms and structures. The present review recognizesand amplifies the importance of compounds with natural origins, which can be used to create safer and more effective antimicrobial drugs by combating microorganisms that are resistant to multiple types of drugs.
1. Introduction
Multidrug-resistant microorganisms (MDRMOs) are progressively being regarded as a major global health challenge.1,2 As a result of conventional antibiotic pressure, bacterial efflux pumps were increased, reducing drug concentrations; enzymes were induced to modify and inactivate antibiotic compounds; or the drug target site was elevated, reducing antibiotic potency (Fig. 1).3,4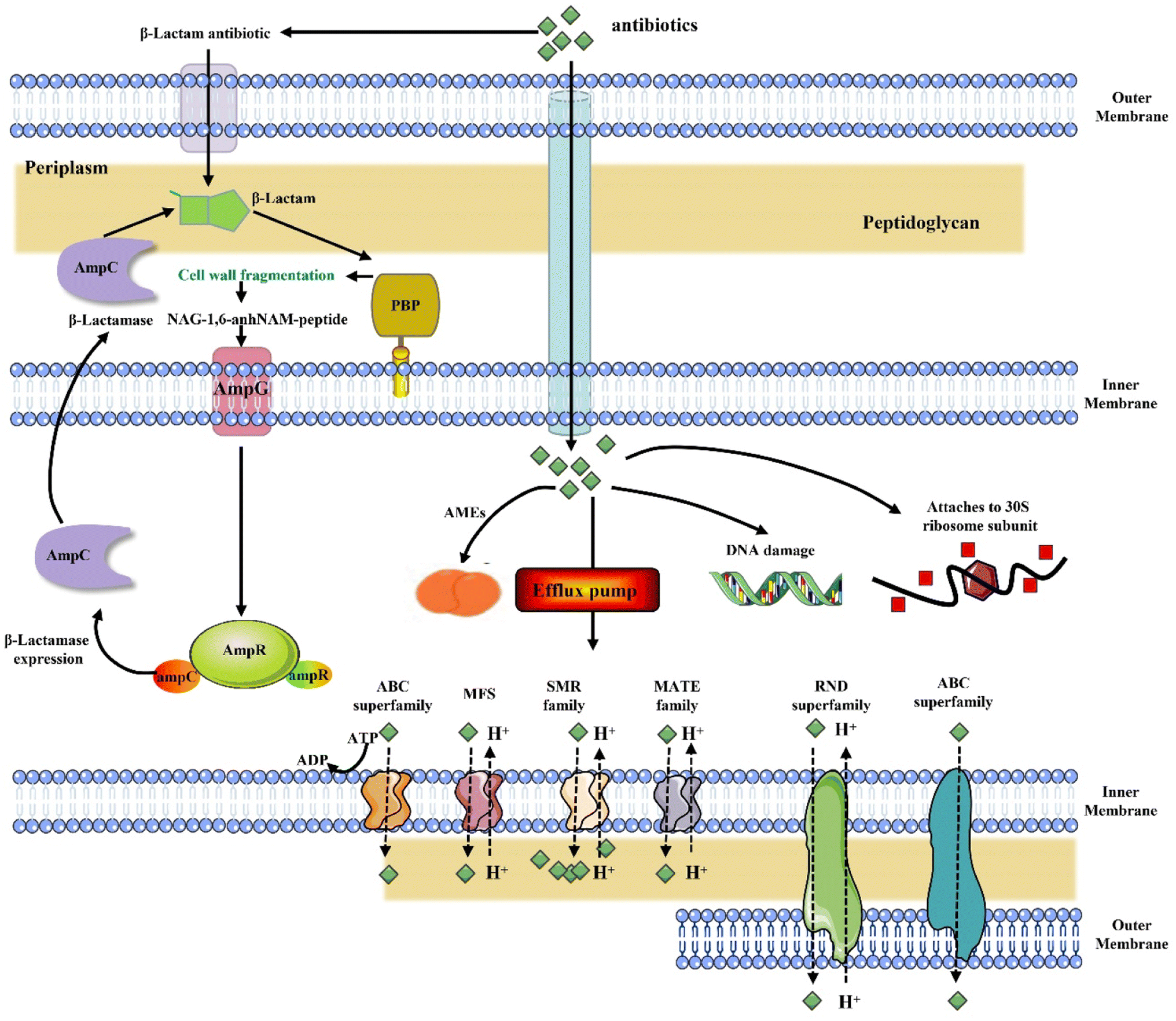 | ||
| Fig. 1 Mechanism of development of multidrug resistance. AMEs: modifying aminoglycoside modifying enzymes; ABC: ATP-binding cassette; MFS: major facilitator superfamily; MATE: multidrug and toxin extrusion; SMR: small multidrug resistance; RND: resistance-nodulation-cell division; PBP: penicillin-binding protein. | ||
Infections by pathogenic bacteria, especially those that are multidrug resistant (MDR), are leading to a growing global health crisis. As a result, MDR bacteria can spread rapidly in hospitals and in the community, posing a staggering burden on health care systems both economically and epidemiologically.5 In addition, the inability of current clinical technologies to accurately diagnose infections in a timely manner further aggravates the resistance situation.6 By 2050, the number of antimicrobial resistance-related deaths could amount to 10 million, which is far more than the number of cancer deaths.2 As we approach the postantibiotic era, normal infections and minor injuries will no longer be treatable. However, in contrast to the rapidly spreading threat of antibiotic resistance, recent decades have seen a dramatic decline in new antibiotic development, with many pharmaceutical companies abandoning antibiotic discovery programs. Due to the subsiding development of antibiotics, alternative therapeutics are needed to treat MDR bacteria, and antimicrobial stewardship necessitates urgent action to stop the evolution of resistance and prevent the emergence of new resistance.
Drug development is aided by natural compounds, especially in regard to the discovery of antimicrobial drugs.7–9 Microorganisms have become increasingly resistant and the use of antibiotics is causing numerous side effects, making it essential to develop new and improved antibacterial agents using plant-based derivatives.10,11 The purpose of this review is to discuss compounds of natural products and derivatives derived from plants that have been investigated as modulators of MDR by inhibiting or otherwise stopping their activity.
2. Molecular antibiotic resistance mechanisms
By overusing antibiotics, microbes have evolved resistance to many conventional antimicrobials. There are several mechanisms that can lead to drug resistance in microbes, including inactivation or modification of antimicrobial drugs, alteration of drug target sites and expulsion of antimicrobial drugs across the cell membrane reduces drug accumulation. The combination of all of these factors contributes to the development of resistance to current antimicrobial therapy.2.1. Alterers: inactivated antimicrobial drugs
Current therapeutic β-lactamase inhibitors are only effective against class A β-lactamases and do not protect against other β-lactamases. A further challenge for β-lactams and β-lactamase inhibitors is that some strains produce multiple enzymes. Despite reports that new β-lactamase inhibitors are appearing increasingly frequently in the medical literature, some of these inhibitors demonstrate good inhibition properties, as well as inhibiting a variety of β-lactamases simultaneously; other than AVE1330 and NXL104, none of these inhibitors have even reached phase I trials.16,17 Molecular modeling and resolution of new lactamase structures may help to overcome one of the most serious threats to human health.
2.2. Blockers: modify the targets of antimicrobial agents
As a dynamic matrix, the cell wall is constantly being constructed and recycled at the same time as enzymes are involved in the process. With each generation, approximately half of a G (−) bacteria's cell wall is remodeled.25–27 A fragment of the cell wall can be biologically recycled for cell wall synthesis, and bacteria as well as eukaryotes use them as messengers for communication. PBP4, a penicillin-binding protein with DD-carboxypeptidase and 4,3-endopeptidase activity, has emerged as a key player in the detection and response to β-lactam antibiotics.28 Muropeptides are also very relevant as signaling molecules that are capable of provoking the multiresistance protein AmpC. When P. aeruginosa is resistant to a β-lactam antibiotic, signaling molecules have been identified in only two peptides over 20.29 Antibiotic resistance can be caused by many mechanisms, including the expression of AmpC, but new mechanisms linking cell-wall recycling to resistance have been described recently.30 There is a metabolic pathway in P. putida that links cell wall recycling to an intrinsic resistance to phosphomycin. As peptidoglycan plays a crucial role in bacterial survival and is remodeled by a large number of proteins, an emerging pathway could link recycling and antibiotic resistance in the near future.
2.3. Expellers: pump out antimicrobial agents
The association between efflux and drug resistance was first described over 40 years ago. For drug resistance to be effective, there must be an equal or greater amount of efflux than influx. In turn, drug entry is determined by the composition of the cell envelope and the presence of porins or other entry channels. There is no doubt that porin loss can contribute to resistance, and it has been observed to occur during treatment.42,43 Additionally, several cases of efflux-mediated resistance have been reported among clinical isolates, some of which involve the overexpression of pumps.
It is essential to have a clear understanding of efflux regulation because the overexpression of efflux proteins is caused by alterations in the regulatory system and transcriptional regulator mutations, including local and global mutations.44,45 There can be differences in efflux regulation between species, as well as within species, based on the physiological state of the cell. There are many pumps that are interconnected and regulated by complex circuits. Inhibiting or deleting one pump can lead to the expression of others.46–48 This complicates the interpretation of resistant phenotypes caused by efflux pump networks.
2.4. Decreased intracellular concentration
3. MDR modulators of natural product-derived compounds from plants
Even though bioactive molecules with plant origins are utilized for a variety of purposes, the use of these molecules for fighting MDR bacteria and restoring the effectiveness of antibiotics in clinics has largely been ignored. According to medicinal chemists, natural products are miscellaneous molecules that have evolved to interact with a wide variety of protein targets to accomplish specific goals.54–57 Hence, natural sources are considered to be an advantageous source for discovering new antimicrobial molecules. Over half of the drugs approved by the American Food and Drug Administration (FDA) are based on or inspired by natural products.The necessary starting materials for drug discovery that are used by pharmaceutical companies have always been provided by nature, since these materials have always solved many complex clinical problems. Natural products have been studied extensively as modulators of MDRs. The MDR modulators of natural product-derived compounds from plants are summarized in Table 1 as three major categories: flavonoids, alkaloids and terpenoids.
| Classification | Compound | Source | Antibacterial activity | Mechanism | Reference |
|---|---|---|---|---|---|
| a MRSA, methicillin-resistant Staphylococcus aureus; VER, vancomycin-resistant Enterococcus; MIC90, lowest drug concentration inhibiting 90% of bacterial or fungi growth; MIC, minimum inhibitory concentration; MDR, multidrug-resistant; UPEC, Uropathogenic Escherichia coli; CRKP, carbapenem-resistant Klebsiella pneumoniae; CRSM, carbapenem-resistant Serratia marcescens. | |||||
| Flavonoids | (1) Isobavachalcone (IBC) | Psoralen | MRSA and VRE (MIC50: 0.5 μg mL; MIC90: 4–8 μg mL−1) | Membrane homeostasis | 66 |
| (2) α-Mangostin (AMG) | |||||
| Quercetin | — | P. aeruginosa and A. baumannii (MIC: 16–256 μg mL−1) | Enzyme inhibition and efflux pump inhibition | 67 | |
| ATCC 25922 (MIC: 1 μg mL−1) | |||||
| E. coli clinical isolates (MIC: 64 μg mL−1) | |||||
| (1) 3,7-Diacylquercetin | Quercetin | Multidrug-resistant gram-positive (MIC: 0.13 to 128 μg mL−1) | Enzymatic inhibition (DNA gyrase and topo IV) | 69 | |
| (2) Quercetin 6′′-acylgalactoside | |||||
| (3) Quercetin 2′′,6′′-diacylgalactoside analogues | |||||
| (1) Procyanidin B3-3-O-gallate | W. uniflora | MRSA | Abnormal cell formation | 71 | |
| (2) Rhamnetin 3-O-(6′′-galloyl)-β-D-glucopyranoside | |||||
| (3) Rhamnetin 3-O-α-L-rhamnopyranoside | |||||
| (4) Quercetin 3-O-(6′′-galloyl)-β-D-glucopyranoside | |||||
| (1) Isobavachalcone; | — | E. coli AG100A and E. aerogenes EA298 isobavachalcone (MIC: 8 μg mL−1) | Efflux pump components (AcrAB, TolC) | 72 | |
| (2) Diospyrone | Diospyrone (MIC: 4 μg mL−1) | ||||
| Lupinifolin | Albizia myriophylla | MDR enterococcal clinical isolates (MIC: 0.5 and 2.0 μg mL−1) | Membrane permeability and salt tolerance | 73 | |
| Phloretin | Apple | E. coli ZJ478 or Salmonella sp. Stain (MIC: 2 μg mL−1) | Cell membranes | 74–76 | |
| Type A procyanidin (TAP | Cinnamomum zeylanicum | UPEC | Biofilm formation | 77 | |
| Artonin I | Morus mesozygia Stapf | MDR bacterial (MIC: 4–8 mg L−1) | Efflux pump and cell membrane | 78 | |
| AFPO | Chalcones | S. aureus 10 strain (MIC: 1024 μg mL−1) | Internal resistance mechanism | 79 | |
| E. coli 06 strain (MIC: 256 μg mL−1) | |||||
| 3′,4′,7-trihydroxyflavone | Kernel | P. stuartii ATCC299645 (MIC: 4 μg mL−1) | Efflux pumps | 80 | |
| Epicatechin gallate (ECg) | Green tea | S. aureus BB568, EMRSA-15, EMRSA-16 (MIC: 128 mg L−1) | Cell membrane | 89 | |
| Curcumin | — | A. baumannii (MIC >256 μg mL−1 | — | 65 | |
| Epigallocatechin gallate | Green tea | A. baumannii (MIC >128–1024 μg mL−1) | — | 65 | |
| Eriodictyol | Citrus fruits | S. aureus USA 300 (MIC >512 μg mL−1) | Inhibitor of SrtA | 92 | |
| Rutin | — | K. pneumoniae ATCC700603 (MIC: 1024 μg mL−1) | Downregulated luxS gene and wabG gene | 63 | |
| E. coli ATCC25922 (MIC: 512 μg mL−1) | |||||
| Baicalein | — | MRSA | Cell wall and Tet K-mediated tetracycline efflux | 70 | |
| Ursolic acid (UA) | Apple pomace | CRKP (MIC: 0.8 mg mL−1) | Cell membrane integrity | 93 | |
| 5-Hydroxy-3,7,4′-trimethoxyflflavone | V. gardneriana | MDR bacterial strains S. aureus 358 and E. coli 27 (MIC≤512 μg mL−1) | — | 94 | |
| Alkaloids | Compound 1f (with a 7-phenyl group) | 7-Substituted cyclophosphamide (CBBR) | MRSA and VRE (MIC: 1-8 μg mL−1) | DNA Topo IV ParE subunit | 102 |
| 1,4-Naphthoquinones | — | MRSA (MIC: 0.0078 to 0.125 mg mL−1) | — | 103 | |
| Chelerythrine (CHE) | — | CRSM (MIC: 125 mg mL−1) | Integrity of the cell membrane | 105 | |
| Berberine hydrochloride (BBH) | Berberine | MDR A. baumannii (MIC: 256 mg L−1) | AdeB and AdeB transporters | 106 | |
| Piperine | White pepper | MDR strains of P. aeruginosa, E. coli (in the presence of 125 mg mL−1) | — | 107 | |
| Squalamine | Dogfish shark squalus acanthias | P. aeruginosa ATCC 27853 (MIC: 8 mg L−1) | Cell membranes | 108 | |
| E. coli ATCC 25922 (MIC: 4 mg L−1) | |||||
| S. aureus ATCC 25923 (MIC: 2 mg L−1) | |||||
| N-tetradecyl derivative (BnI-14) | Quaternary ammonium compounds (QACs) | S. aureus ATCC 29213 (MIC: 0.03 μg mL−1) | Membrane disruption and DNA | 110 | |
| S. aureus ATCC 29213 clinical/MRSA (MIC: 0.12 μg mL−1) | |||||
| E. coli clinical (MIC: 0.98 μg mL−1) | |||||
| Arborinine | R. angustifolia | Candida albicans arborinine (MIC: 250 μg mL−1) | ICL1 gene | 111 | |
| Arborinine (MIC: 500 μg mL−1) | |||||
| Catharanthine | Catharanthus roseous | P. aeruginosa (MIC: 400 mg L−1) | Efflux pump proteins MexA, MexB and OprM | 113 | |
| Chanoclavine | I. muricata | E. coli | Efflux pumps and the cell envelope | 115 | |
| Terpenoids | Limonene | — | E. coli (MIC: 16 μg mL−1) | Cell membrane and DNA transcription and translation | 118 and 119 |
| Carvacrol | — | S. aureus (MIC: 256 μg mL−1) | Efflux pump inhibition | 120 and 122 | |
| Thymol | |||||
| Oleanolic acid | — | E. faecalis (MIC: 6.25 mg L−1) | Efflux pump | 121 | |
| MRSA COLOXA (MIC: 1600 mg L−1) | |||||
| Clerodane diterpene 16α-hydroxycleroda-3,13(14)-Z-dien-15,16-olide (CD) | Polyalthia longifolia | MRSA-ST1745/ST2071/P4620 (MIC: 31.25 μg mL−1) | EtBr efflux | 123 | |
| MRSA-P4627/P4423/ST3151 (MIC: 15.62 μg mL−1) | |||||
| Geraniol (Ger) | Geranium oil | C. albicans (MIC: 225 μg mL−1) | Cell wall integrity and cell adhesion | 124 | |
| Oleanane triterpenoid aegicerin | ClaVija procera | MTB (MIC: 1.6-3.12 μg mL−1) | — | 125 | |
| Artesunate (ART) | Artemisia annua L | MDR E. coli isolates | mRNA expression of qnrB and qnrS | 126 | |
| Glycyrrhizin (GLY) | Glycyrrhiza glabra | MDR9 (MIC: 40 mg mL−1) | — | 127 | |
| B1045 (MIC: 15 mg mL−1) | |||||
| Conjugates | — | S. aureus (MIC: 20 μg mL−1) | — | 129 | |
| E. coli (MIC: 20 μg mL−1) | |||||
| 1,8-Cineole | Rosemary volatile oil | E. coli | Biofilm biomass disruption | 135 | |
3.1. Flavonoids
Among the most important classes of phenolic compounds are flavonoids, which are plant secondary metabolites in nonglycosylated (aglycones) or glycosidic states. In addition to the 2-phenylchromane ring system, flavonoids also have different subclasses depending on the substitution at ring B and oxidation status of ring C.58,59 There has been considerable evidence that flavonoids possess a variety of pharmacological properties, including anticancer, anti-inflammatory, and antiviral properties.60–62 Additionally, flavonoids have been found to exhibit antibacterial properties against many pathogenic microorganisms, such as Staphylococcus aureus (S. aureus), Vibrio harveyi, Pseudomonas aeruginosa (P. aeruginosa), and Enterococcus faecalis (E. faecalis).63–65 There are over 8000 known flavonoids in plants, making them a potentially valuable resource for new antibiotic discoveries. We reviewed the potential antimicrobial activity of flavonoids against fungal and bacterial infections in Sections 3.1.1–3.1.13, and their structure was drown in Fig. 2.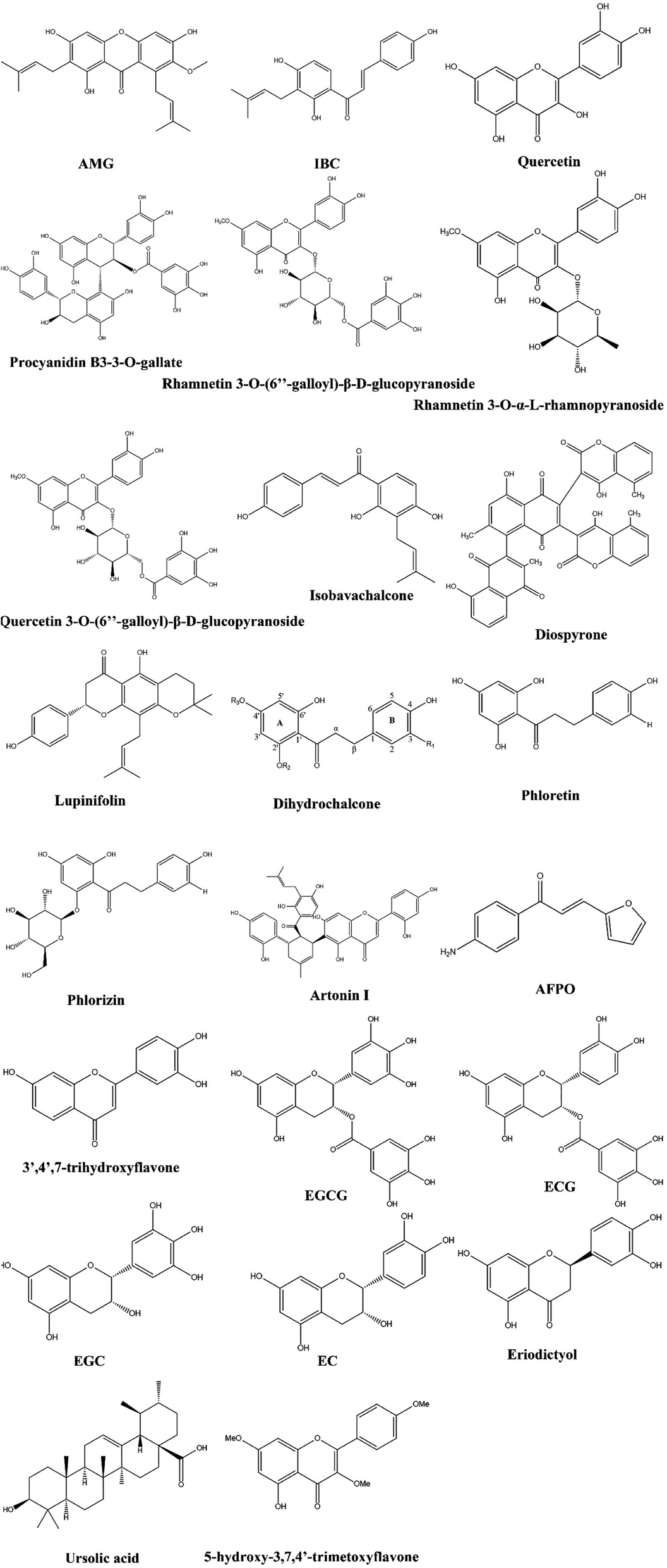 | ||
| Fig. 2 Structure of flavonoids having MDR modulatory activity. | ||
A high percentage of therapeutic antimicrobials are no longer effective against MDR strains, and EGCG should be able to improve their treatment. Another study found that EGCG was effective in modifying resistance in Campylobacter spp. by inhibiting efflux pump activity in the wild-type strain NCTC 11168 as well as in the cmeB, cmeF, and cmeR mutants.91 EGCG prevents conjugative drug resistance by reducing the transfer of drug resistance plasmids in a dose-dependent manner. Tetracycline resistance is reversed by EGCG when it inhibits the Tet(K) efflux pump (EP) in staphylococci, resulting in antibiotic sensitivity in infected staphylococci.92
Curcumin (CCM) and EGCG were investigated and evaluated against multidrug-resistant strains of A. baumannii via checkerboard and time-kill assays. CCM had an MIC of >256 μg mL−1 against all strains of A. baumannii, whereas EGCG had an MIC of 128–1024 μg mL−1. Five of nine isolates showed synergy, and four isolates showed additive effects, according to checkerboard studies. CCM's MIC was reduced by 3- to 7-fold when EGCG was added, and a CCM MIC of 4 μg mL−1 was the strongest interaction. In comparison with the most effective polyphenol alone, a combination of CCM and EGCG (1![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :8 and 1:4) demonstrated 4- to 5-log reductions in viable counts after 24 hours. Although CCM alone exhibits little antibacterial activity, EGCG significantly enhances its activity against MDR A. baumannii. Combinations of these two substances may be useful for medicinally treating or preventing infections caused by A. baumannii.65
:8 and 1:4) demonstrated 4- to 5-log reductions in viable counts after 24 hours. Although CCM alone exhibits little antibacterial activity, EGCG significantly enhances its activity against MDR A. baumannii. Combinations of these two substances may be useful for medicinally treating or preventing infections caused by A. baumannii.65
Through its direct binding to the peptidoglycan in the cell wall, baicalein can work synergistically with tetracycline to compromise the integrity of the cell wall. In addition to inhibiting Tet K-mediated tetracycline efflux, baicalein inhibits Tet M and other pumps that are responsible for methicillin-resistant S. aureus (MRSA) growth.71
An antimicrobial agent found in apple pomace, ursolic acid (UA), exhibits antimicrobial properties against some microorganisms. UA was found to be effective against Klebsiella pneumoniae (CRKP) at an MIC of 0.8 mg mL−1. The drug disrupted CRKP's cell membrane integrity, inhibited biofilm formation, and inactivated cells that were encased in biofilms.94
The compound 5-hydroxy-3,7,4′-trimetoxyflavone, which has a chemical formula of C18H16O6, was isolated from the leaves of V. gardneriana. In combination with norfloxacin and gentamicin, this flavone exhibits antimicrobial activity against the MDR bacteria S. aureus 358 and E. coli 27. Hence, this natural compound contributes to the control of resistant bacteria by increasing antibiotic activity.95
3.2. Alkaloids
Compared to synthetic antibacterial agents, alkaloids have fewer side effects, a higher efficacy and superior antibacterial properties against both resistance acquired pathogens and nonresistance acquired pathogens.96,97 In addition, alkaloids combined with antibiotics increased their efficacy, reduced side effects, and increased their concentration.98–100 In addition to inhibiting protein synthesis and efflux pumps, alkaloids inhibit the formation of biofilms, inhibit drug deactivating enzymes, and destroy cell membranes and cell walls.101,102 Using this natural gift to its full potential requires advanced studies. A variety of advanced methods, such as virtual screening, molecular modeling, and biological studies, can be used to alter the properties of these substances. This new era of antimicrobial agents may arise through the development of these alkaloids (Fig. 3).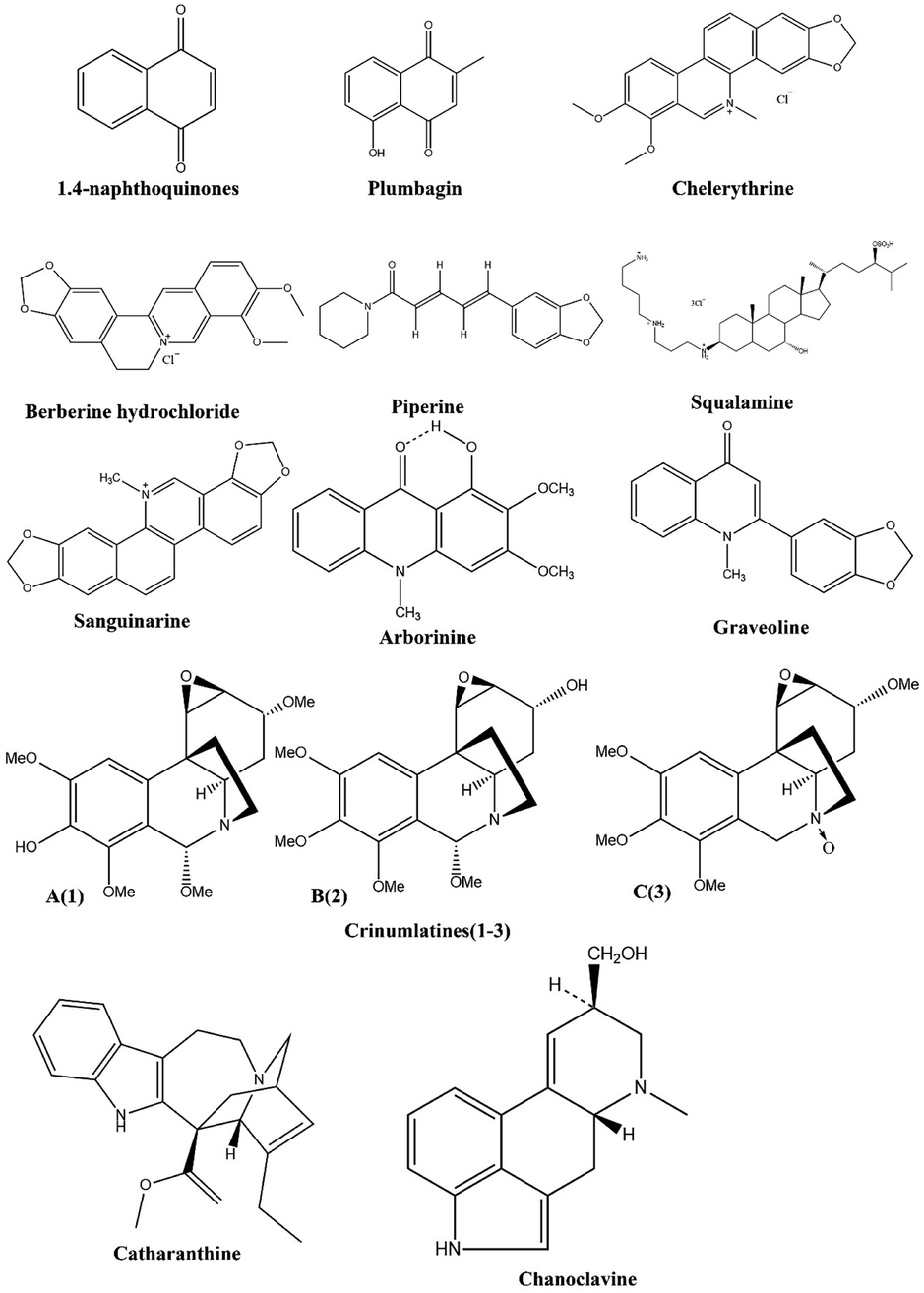 | ||
| Fig. 3 Structure of alkaloids having MDR modulatory activity. | ||
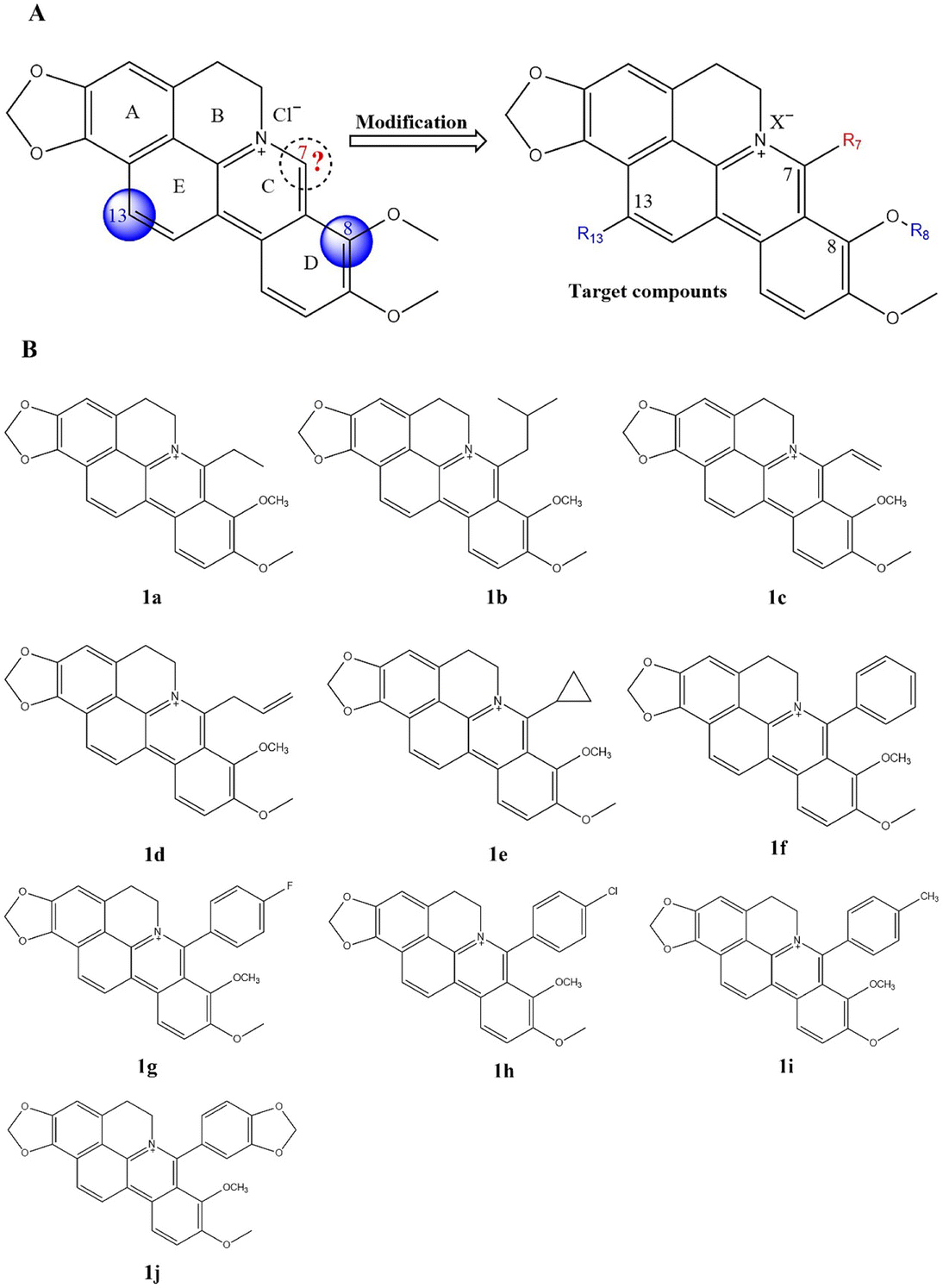 | ||
| Fig. 4 (A) The chemical structure of CBBR and the structure modification strategy. (B.) the structure of the target compounds 1a–j. | ||
Arborinine and graveoline exhibit inhibitory activity against C. albicans at 250 and 500 lg/ml, respectively. MIC/MFC indicated that these alkaloids played a bactericidal role. Analysis of gene and protein expression revealed effective effects for both compounds on ICL1 gene andprotein expression. Hence, arborinine and graveoline appear to be promising inhibitors of ICL1 in C. albicans glyoxylate cycles.112 A phytochemical investigation of the 90% ethanol aqueous extract of the bulbs of Crinum latifolium led to the isolation of three new crinane-type alkaloids, designated crinumlatines A-C (1–3). Compounds 1–3 exhibited some antimicrobial activity against the tested Gram-negative bacteria with minimum inhibitory concentration values less than 50 lg mL−1.113
Catharanthine was deduced first through in silico inhibition of efflux pump proteins MexA, MexB and OprM. Catharanthine-induced efflux pump inhibition may be useful for lowering antibiotic doses, reducing drug resistance, and increasing the efficacy of old antibiotics against multidrug-resistant bacteria.114
Plants selected for anti-mastitis protection contain higher concentrations of phytochemicals, with alkaloids of A. sativum and B. persicum showing significant antibacterial activity compared to that of the other selections. Additionally, ATCC strains show greater inhibition of phytochemicals of tested plant species compared to that of MDR bacterial strains.115
We can see that chanoclavine may make it easier to repurpose antibiotics that have become nonfunctional because of acquired bacterial resistance. By inhibiting efflux pumps, weakening the cell envelope, and increasing cell growth, the molecule synergizes with conventional antibiotics to target bacterial cell membranes or cell walls.116
3.3. Terpenoids
Terpenoids, which are abundant in nature and produced in response to microbial attack, have huge potential as antimicrobial agents through a variety of mechanisms, such as disruption of membranes, anti-quorum sensing, inhibition of protein synthesis and ATP.92,117,118 Combinations of terpenoids and antimicrobials, such as combination therapy, have increased the effectiveness of treatment against multidrug resistant microorganisms by showing synergy (Fig. 5).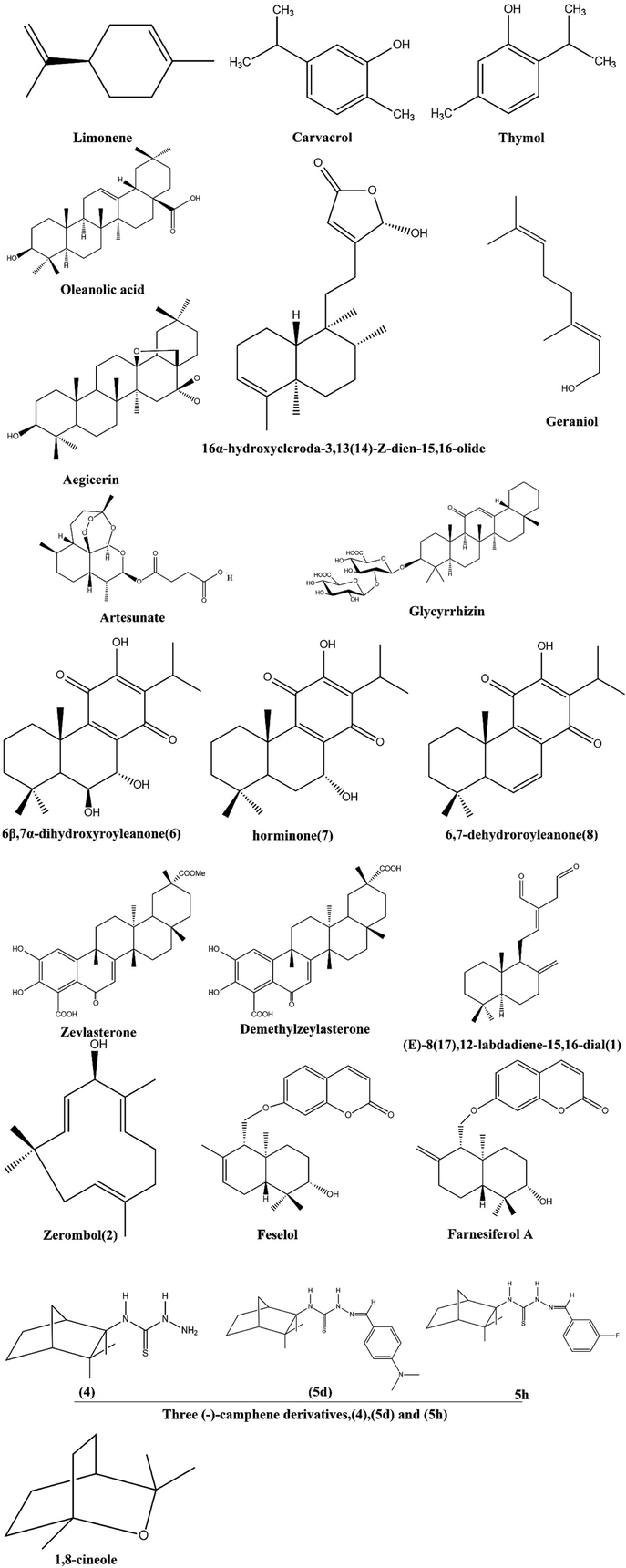 | ||
| Fig. 5 Structure of terpenoids having MDR modulatory activity. | ||
Feselol and farnesiferol A are sesquiterpene coumarins that are obtained by extracting aerial parts of Ferula vesceritensis (Apiaceae) in dichloromethane, which is associated with a model of the recombinant nucleotide binding site of Cryptosporidium parvum's enteropathogenic efflux pump.135
For the first time, the antibiofilm activity of 1,8-cineole against E. coli producing MDR ESBLs was demonstrated.136 There is no doubt that the compound is capable of causing substantial bacterial death in biofilm-attached as well as biofilm-released cells. The (−)-camphene derivatives tested in the present study, particularly (4) (5d) and (5 h), were shown to be promising anti-TB molecule scaffolds due to the low MIC values in acidic pH against the reference strain H37Rv and MDR M. tuberculosis clinical isolates, their low cytotoxicity and the synergism observed with PZA.137
All bacteria tested were inhibited by the highest polar fraction of Cistus ladaniferus essential oil, which contained mono- and sesquiterpene alcohols with MIC values ranging from 0.05 to 0.8 mg mL−1.138 There was high activity of this fraction against the MDR strsain Enterobacter aerogenes EA289, and its mechanism involves a distortion of the cell wall with detachment of the outer cytoplasmic membrane.
In vitro antibacterial activity of the essential oil of E. camaldulensis against multidrug resistant A. baumannii wound isolates, demonstrating the plant's traditional use for wounds. As constituents of the E. camaldulensis oil, polar terpene compounds and spathulenol may contribute to the observed antibacterial activity.139 Additionally, synergistic interactions of the antibiotics ciprofloxacin, gentamicin and polymyxin B with E. camaldulsensis essential oils were detected in this study for the first time against MDR A. baumannii isolates.
4. Conclusion and perspectives
In contemporary society, antibiotics have become an important part of everyday life; the wonders of antibiotics reveal how useful nature can be. Multidrug resistance (MDR) as well as cross-resistance to other drugs have become global concerns due to inappropriate use and overprescription of medications. In response to this emerging problem, we are entering an era of responsibility. Novel solutions are urgently needed to address this significant issue. Natural products are currently being investigated for their antimicrobial properties. Based on the current literature, it appears that there is an urgent need to coordinate efforts for meaningful research and for finding novel alternatives that utilize natural products.The present study presents a wealth of information regarding the phytocomplexes and mode of action of purified antimicrobials that are isolated and purified from potential medicinal plants. Additionally, these components can be used to design new nutraceuticals or other drugs that are effective. New therapeutics will, however, be discovered by exploring the additional bioactivity of the corresponding species and genus.
Conflicts of interest
There are no conflicts to declare.References
- N. S. Isles, A. Mu, J. C. Kwong, B. P. Howden and T. P. Stinear, Gut microbiome signatures and host colonization with multidrug-resistant bacteria, Trends Microbiol., 2022, 30, 853–865 CrossRef CAS PubMed.
- X. Pang, D. Li, J. Zhu, J. Cheng and G. Liu, Beyond Antibiotics: Photo/Sonodynamic Approaches for Bacterial Theranostics, Nanomicro Lett., 2020, 12, 144 CAS.
- F. Baquero, J. L. Martinez, V. F. Lanza, J. Rodriguez-Beltran, J. C. Galan, A. San Millan, R. Canton and T. M. Coque, Evolutionary Pathways and Trajectories in Antibiotic Resistance, Clin. Microbiol. Rev., 2021, 34, e0005019 CrossRef CAS PubMed.
- M. F. Chellat, L. Raguz and R. Riedl, Targeting Antibiotic Resistance, Angew. Chem., Int. Ed. Engl., 2016, 55, 6600–6626 CrossRef CAS PubMed.
- S. H. Jung, C. M. Ryu and J. S. Kim, Bacterial persistence: Fundamentals and clinical importance, J. Microbiol., 2019, 57, 829–835 CrossRef PubMed.
- A. J. Trotter, A. Aydin, M. J. Strinden and J. O'Grady, Recent and emerging technologies for the rapid diagnosis of infection and antimicrobial resistance, Curr. Opin. Microbiol., 2019, 51, 39–45 CrossRef CAS PubMed.
- M. S. Butler, Natural products to drugs: natural product-derived compounds in clinical trials, Nat. Prod. Rep., 2008, 25, 475–516 RSC.
- E. K. Davison and M. A. Brimble, Natural product derived privileged scaffolds in drug discovery, Curr. Opin. Chem. Biol., 2019, 52, 1–8 CrossRef CAS PubMed.
- T. Rodrigues, D. Reker, P. Schneider and G. Schneider, Counting on natural products for drug design, Nat. Chem., 2016, 8, 531–541 CrossRef CAS PubMed.
- K. H. Kyung, Antimicrobial properties of allium species, Curr. Opin. Biotechnol., 2012, 23, 142–147 CrossRef CAS PubMed.
- M. Daglia, Polyphenols as antimicrobial agents, Curr. Opin. Biotechnol., 2012, 23, 174–181 CrossRef CAS PubMed.
- K. Bush and P. A. Bradford, beta-Lactams and beta-Lactamase Inhibitors: An Overview, Cold Spring Harbor Perspect. Med., 2016, 6, a025247 CrossRef PubMed.
- K. M. Papp-Wallace, A. Endimiani, M. A. Taracila and B. R. A. Carbapenems, past, present, and future, Antimicrob. Agents Chemother., 2011, 55, 4943–4960 CrossRef CAS PubMed.
- V. Miguel-Ruano and J. A. Hermoso, Teaching an old dog new tricks: repurposing beta-lactams, Trends Pharmacol. Sci., 2021, 42, 617–619 CrossRef CAS PubMed.
- R. B. Hamed, J. R. Gomez-Castellanos, L. Henry, C. Ducho, M. A. McDonough and C. J. Schofield, The enzymes of beta-lactam biosynthesis, Nat. Prod. Rep., 2013, 30, 21–107 RSC.
- K. Bush and P. A. Bradford, Interplay between beta-lactamases and new beta-lactamase inhibitors, Nat. Rev. Microbiol., 2019, 17, 295–306 CrossRef CAS PubMed.
- D. Yahav, C. G. Giske, A. Gramatniece, H. Abodakpi, V. H. Tam and L. Leibovici, New beta-Lactam-beta-Lactamase Inhibitor Combinations, Clin. Microbiol. Rev., 2020, 34, e00115–e00120 CrossRef PubMed.
- M. S. Ramirez and M. E. Tolmasky, Aminoglycoside modifying enzymes, Drug Resistance Updates, 2010, 13, 151–171 CrossRef CAS PubMed.
- B. O. Costa, M. H. Cardoso and O. L. Franco, Development of Peptides that Inhibit Aminoglycoside-Modifying Enzymes and beta-Lactamases for Control of Resistant Bacteria, Curr. Protein Pept. Sci., 2020, 21, 1011–1026 CrossRef CAS PubMed.
- G. C. Sati, V. A. Sarpe, T. Furukawa, S. Mondal, M. Mantovani, S. N. Hobbie, A. Vasella, E. C. Bottger and D. Crich, Modification at the 2'-Position of the 4,5-Series of 2-Deoxystreptamine Aminoglycoside Antibiotics To Resist Aminoglycoside Modifying Enzymes and Increase Ribosomal Target Selectivity, ACS Infect. Dis., 2019, 5, 1718–1730 CrossRef CAS PubMed.
- P. Macheboeuf, C. Contreras-Martel, V. Job, O. Dideberg and A. Dessen, Penicillin binding proteins: key players in bacterial cell cycle and drug resistance processes, FEMS Microbiol. Rev., 2006, 30, 673–691 CrossRef CAS PubMed.
- M. W. Shalaby, E. M. E. Dokla, R. A. T. Serya and K. A. M. Abouzid, Penicillin binding protein 2a: An overview and a medicinal chemistry perspective, Eur. J. Med. Chem., 2020, 199, 112312 CrossRef CAS PubMed.
- M. Goel, R. Kalra, P. Ponnan, J. Jayaweera and W. W. Kumbukgolla, Inhibition of penicillin-binding protein 2a (PBP2a) in methicillin resistant Staphylococcus aureus (MRSA) by combination of oxacillin and a bioactive compound from Ramalinaroesleri, Microb. Pathog., 2021, 150, 104676 CrossRef CAS PubMed.
- T. Dominguez-Gil, R. Molina, M. Alcorlo and J. A. Hermoso, Renew or die: The molecular mechanisms of peptidoglycan recycling and antibiotic resistance in Gram-negative pathogens, Drug Resistance Updates, 2016, 28, 91–104 CrossRef PubMed.
- M. A. Boudreau, J. F. Fisher and S. Mobashery, Messenger functions of the bacterial cell wall-derived muropeptides, Biochemistry, 2012, 51, 2974–2990 CrossRef CAS PubMed.
- S. Cho, Q. Wang, C. P. Swaminathan, D. Hesek, M. Lee, G. J. Boons, S. M. Mobashery, S. Mobashery and R. A. Mariuzza, Structural insights into the bactericidal mechanism of human peptidoglycan recognition proteins, Proc. Natl. Acad. Sci. U. S. A., 2007, 104, 8761–8766 CrossRef CAS PubMed.
- K. L. Woodhams, J. M. Chan, J. D. Lenz, K. T. Hackett and J. P. Dillard, Peptidoglycan fragment release from Neisseria meningitidis, Infect. Immun., 2013, 81, 3490–3498 CrossRef CAS PubMed.
- M. Lee, D. Hesek, B. Blazquez, E. Lastochkin, B. Boggess, J. F. Fisher and S. Mobashery, Catalytic spectrum of the penicillin-binding protein 4 of Pseudomonas aeruginosa, a nexus for the induction of beta-lactam antibiotic resistance, J. Am. Chem. Soc., 2015, 137, 190–200 CrossRef CAS PubMed.
- M. Lee, S. Dhar, S. De Benedetti, D. Hesek, B. Boggess, B. Blazquez, K. Mathee and S. Mobashery, Muropeptides in Pseudomonas aeruginosa and their Role as Elicitors of beta-Lactam-Antibiotic Resistance, Angew. Chem., Int. Ed. Engl., 2016, 55, 6882–6886 CrossRef CAS PubMed.
- J. Gisin, A. Schneider, B. Nagele, M. Borisova and C. Mayer, A cell wall recycling shortcut that bypasses peptidoglycan de novo biosynthesis, Nat. Chem. Biol., 2013, 9, 491–493 CrossRef CAS PubMed.
- L. Yi and X. Lu, New Strategy on Antimicrobial-resistance: Inhibitors of DNA Replication Enzymes, Curr. Med. Chem., 2019, 26, 1761–1787 CrossRef CAS PubMed.
- A. Seddek, T. Annamalai and Y. C. Tse-Dinh, Type IA Topoisomerases as Targets for Infectious Disease Treatments, Microorganisms, 2021, 9, 86 CrossRef CAS PubMed.
- D. C. Hooper, Mechanisms of action of antimicrobials: focus on fluoroquinolones, Clin. Infect. Dis., 2001, 15(Suppl 1), S9–S15 CrossRef PubMed.
- S. Rebuffat, Ribosomally synthesized peptides, foreground players in microbial interactions: recent developments and unanswered questions, Nat. Prod. Rep., 2022, 39, 273–310 RSC.
- D. N. Wilson, Ribosome-targeting antibiotics and mechanisms of bacterial resistance, Nat. Rev. Microbiol., 2014, 12, 35–48 CrossRef CAS PubMed.
- J. L. M. Liljeruhm, L. Bao, T. Truu, M. Calvo-Noriega, N. S. Freyer, A. Liiv, J. Wang, R. C. Blanco, R. Ero, J. Remme and A. C. Forster, Plasticity and conditional essentiality of modification enzymes for domain V of Escherichia coli 23S ribosomal RNA, RNA, 2022, 28, 796–807 CrossRef CAS PubMed.
- V. Chandrasekaran, N. Desai, N. O. Burton, H. Yang, J. Price, E. A. Miska and V. Ramakrishnan, Visualizing formation of the active site in the mitochondrial ribosome, Elife, 2021, 10, e68806 CrossRef CAS PubMed.
- F. Prosdocimi, G. S. Zamudio, M. Palacios-Perez and S. M. V. J. Torres de Farias, The Ancient History of Peptidyl Transferase Center Formation as Told by Conservation and Information Analyses, Life, 2020, 10, 134 CrossRef CAS PubMed.
- Z. D. Aron, A. Mehrani, E. D. Hoffer, K. L. Connolly, P. Srinivas, M. C. Torhan, J. N. Alumasa, M. Cabrera, D. Hosangadi, J. S. Barbor, S. C. Cardinale, S. M. Kwasny, L. R. Morin, M. M. Butler, T. J. Opperman, T. L. Bowlin, A. Jerse, S. M. Stagg, C. M. Dunham and K. C. Keiler, trans-Translation inhibitors bind to a novel site on the ribosome and clear Neisseria gonorrhoeae in vivo, Nat. Commun., 2021, 12, 1799 CrossRef CAS PubMed.
- E. Breiner-Goldstein, Z. Eyal, D. Matzov, Y. Halfon, G. Cimicata, M. Baum, A. Rokney, A. V. Ezernitchi, A. N. Lowell, J. J. Schmidt, H. Rozenberg, E. Zimmerman, A. Bashan, L. Valinsky, Y. Anzai, D. H. Sherman and A. Yonath, Ribosome-binding and anti-microbial studies of the mycinamicins, 16-membered macrolide antibiotics from Micromonospora griseorubida, Nucleic Acids Res., 2021, 49, 9560–9573 CrossRef CAS PubMed.
- M. L. Pinel-Marie, R. Brielle, C. Riffaud, N. Germain-Amiot, N. Polacek and B. Felden, RNA antitoxin SprF1 binds ribosomes to attenuate translation and promote persister cell formation in Staphylococcus aureus, Nat. Microbiol., 2021, 6, 209–220 CrossRef CAS PubMed.
- S. Kumar, M. Lekshmi, A. Parvathi, M. Ojha, N. Wenzel and M. F. Varela, Functional and Structural Roles of the Major Facilitator Superfamily Bacterial Multidrug Efflux Pumps, Microorganisms, 2020, 8, 266 CrossRef CAS PubMed.
- L. Vachova, V. Stovicek, O. Hlavacek, O. Chernyavskiy, L. Stepanek, L. Kubinova and Z. Palkova, Flo11p, drug efflux pumps, and the extracellular matrix cooperate to form biofilm yeast colonies, J. Cell Biol., 2011, 194, 679–687 CrossRef CAS PubMed.
- P. Blanco, S. Hernando-Amado, J. A. Reales-Calderon, F. Corona, F. Lira, M. Alcalde-Rico, A. Bernardini, M. B. Sanchez and J. L. Martinez, Bacterial Multidrug Efflux Pumps: Much More Than Antibiotic Resistance Determinants, Microorganisms, 2016, 4, 14 CrossRef PubMed.
- I. T. Paulsen, Multidrug efflux pumps and resistance: regulation and evolution, Curr. Opin. Microbiol., 2003, 6, 446–451 CrossRef CAS PubMed.
- D. Du, X. Wang-Kan, A. Neuberger, H. W. van Veen, K. M. Pos, L. J. V. Piddock and B. F. Luisi, Multidrug efflux pumps: structure, function and regulation, Nat. Rev. Microbiol., 2018, 16, 523–539 CrossRef CAS PubMed.
- J. A. Delmar, C. C. Su and E. W. Yu, Bacterial multidrug efflux transporters, Annu. Rev. Biophys., 2014, 43, 93–117 CrossRef CAS PubMed.
- D. Du, H. W. van Veen and B. F. Luisi, Assembly and operation of bacterial tripartite multidrug efflux pumps, Trends Microbiol., 2015, 23, 311–319 CrossRef CAS PubMed.
- C. R. Lee, J. H. Lee, M. Park, K. S. Park, I. K. Bae, Y. B. Kim, C. J. Cha, B. C. Jeong and S. H. Lee, Biology of Acinetobacter baumannii: Pathogenesis, Antibiotic Resistance Mechanisms, and Prospective Treatment Options, Front. Cell. Infect. Microbiol., 2017, 7, 55 Search PubMed.
- C. O. Vrancianu, I. Gheorghe, I. B. Czobor and M. C. Chifiriuc, Antibiotic Resistance Profiles, Molecular Mechanisms and Innovative Treatment Strategies of Acinetobacter baumannii, Microorganisms, 2020, 8, 935 CrossRef CAS PubMed.
- Y. Smani, A. Fabrega, I. Roca, V. Sanchez-Encinales, J. Vila and J. Pachon, Role of OmpA in the multidrug resistance phenotype of Acinetobacter baumannii, Antimicrob. Agents Chemother., 2014, 58, 1806–1808 CrossRef PubMed.
- P. Espinal, A. Pantel, D. Rolo, S. Marti, R. Lopez-Rojas, Y. Smani, J. Pachon, J. Vila and J. P. Lavigne, Relationship Between Different Resistance Mechanisms and Virulence in Acinetobacter baumannii, Microb. Drug Resist., 2019, 25, 752–760 CrossRef CAS PubMed.
- Y. Sato, Y. Unno, S. Kawakami, T. Ubagai and Y. Ono, Virulence characteristics of Acinetobacter baumannii clinical isolates vary with the expression levels of omps, J. Med. Microbiol., 2017, 66, 203–212 CrossRef PubMed.
- A. L. Harvey, R. Edrada-Ebel and R. J. Quinn, The re-emergence of natural products for drug discovery in the genomics era, Nat. Rev. Drug Discov., 2015, 14, 111–129 CrossRef CAS PubMed.
- W.-H. Jesse and J. C. V. Li, Drug discovery and natural products: end of an era or an endless frontier?, Science, 2009, 325, 161–165 CrossRef PubMed.
- Z. Guo, Y. Tang, W. Tang and Y. Chen, Heptose-containing bacterial natural products: structures, bioactivities, and biosyntheses, Nat. Prod. Rep., 2021, 38, 1887–1909 RSC.
- L. Zhang, J. Song, L. Kong, T. Yuan, W. Li, W. Zhang, B. Hou, Y. Lu and G. Du, The strategies and techniques of drug discovery from natural products, Pharmacol. Ther., 2020, 216, 107686 CrossRef CAS PubMed.
- N. F. Shamsudin, Q. U. Ahmed, S. Mahmood, S. A. Ali Shah, A. Khatib, S. Mukhtar, M. A. Alsharif, H. Parveen and Z. A. Zakaria, Antibacterial Effects of Flavonoids and Their Structure-Activity Relationship Study: A Comparative Interpretation, Molecules, 2022, 27, 1149 CrossRef CAS PubMed.
- S. Lin, J. D. Wade and S. Liu, De Novo Design of Flavonoid-Based Mimetics of Cationic Antimicrobial Peptides: Discovery, Development, and Applications, Acc. Chem. Res., 2020, 54, 104–119 CrossRef PubMed.
- A. T. Rufino, V. M. Costa, F. Carvalho and E. Fernandes, Flavonoids as antiobesity agents: A review, Med. Res. Rev., 2021, 41, 556–585 CrossRef CAS PubMed.
- V. Vukics and A. Guttman, Structural characterization of flavonoid glycosides by multi-stage mass spectrometry, Mass Spectrom. Rev., 2010, 29, 1–16 CrossRef CAS PubMed.
- L. Pourcel, J. Routaboul, V. Cheynier, L. Lepiniec and I. Debeaujon, Flavonoid oxidation in plants: from biochemical properties to physiological functions, Trends Plant Sci., 2007, 12, 29–36 CrossRef CAS PubMed.
- Z. Wang, Z. Ding, Z. Li, Y. Ding, F. Jiang and J. Liu, Antioxidant and antibacterial study of 10 flavonoids revealed rutin as a potential antibiofilm agent in Klebsiella pneumoniae strains isolated from hospitalized patients, Microb. Pathog., 2021, 159, 105121 CrossRef CAS PubMed.
- C. J. Raorane, J.-H. Lee, Y.-G. Kim, S. K. Rajasekharan, R. García-Contreras and J. Lee, Antibiofilm and Antivirulence Efficacies of Flavonoids and Curcumin Against Acinetobacter baumannii, Front. Microbiol., 2019, 10, 990 CrossRef PubMed.
- W. B. Jonathan and D. W. W., In vitro activity of curcumin in combination with epigallocatechin gallate (EGCG) versus multidrug-resistant Acinetobacter baumannii, BMC Microbiol., 2014, 14, 172 CrossRef PubMed.
- M. Song, Y. Liu, T. Li, X. Liu, Z. Hao, S. Ding, P. Panichayupakaranant, K. Zhu and J. Shen, Plant Natural Flavonoids Against Multidrug Resistant Pathogens, Adv. Sci., 2021, 8, e2100749 CrossRef PubMed.
- B. J. Denny, P. A. Lambert and P. W. West, The flavonoid galangin inhibits the L1 metallo-beta-lactamase from Stenotrophomonas maltophilia, FEMS Microbiol. Lett., 2002, 208, 21–24 CAS.
- A. Pal and A. Tripathi, Quercetin inhibits carbapenemase and efflux pump activities among carbapenem-resistant Gram-negative bacteria, APMIS, 2020, 128, 251–259 CrossRef CAS PubMed.
- S. Das, S. Batra, P. P. Gupta, M. Kumar, V. K. Srivastava, A. Jyoti, N. Singh and S. Kaushik, Identification and evaluation of quercetin as a potential inhibitor of naphthoate synthase fromEnterococcus faecalis, J. Mol. Recognit., 2019, 32, e2802 CAS.
- A. M. Hossion, Y. Zamami, R. K. Kandahary, T. Tsuchiya, W. Ogawa, A. Iwado and K. Sasaki, Quercetin diacylglycoside analogues showing dual inhibition of DNA gyrase and topoisomerase IV as novel antibacterial agents, J. Med. Chem., 2011, 54, 3686–3703 CrossRef CAS PubMed.
- S. Qu, C. Dai, Z. Shen, Q. Tang, H. Wang, B. Zhai, L. Zhao and Z. Hao, Mechanism of Synergy Between Tetracycline and Quercetin Against Antibiotic Resistant Escherichia coli, Front. Microbiol., 2019, 10, 2536 CrossRef PubMed.
- J. S. Yu, J. H. Kim, L. Rashan, I. Kim, W. Lee and K. H. Kim, Potential Antimicrobial Activity of Galloyl-Flavonoid Glycosides From Woodfordia uniflora Against Methicillin-Resistant Staphylococcus aureus, Front. Microbiol., 2021, 12, 784504 CrossRef PubMed.
- V. Kuete, B. Ngameni, J. G. Tangmouo, J. M. Bolla, S. Alibert-Franco, B. T. Ngadjui and J. M. Pages, Efflux pumps are involved in the defense of Gram-negative bacteria against the natural products isobavachalcone and diospyrone, Antimicrob. Agents Chemother., 2010, 54, 1749–1752 CrossRef CAS PubMed.
- W. Sianglum, K. Muangngam, N. Joycharat and S. P. Voravuthikunchai, Mechanism of Action and Biofilm Inhibitory Activity of Lupinifolin Against Multidrug-Resistant Enterococcal Clinical Isolates, Microb. Drug Resist., 2019, 25, 1391–1400 CrossRef CAS PubMed.
- J. Wang, R. Yang, Z. Xiao, Q. Xu, P. Li and F. Ma, Dihydrochalcones in Malus inhibit bacterial growth by reducing cell membrane integrity, Food Funct., 2020, 11, 6517–6527 RSC.
- D. Barreca, E. Bellocco, G. Lagana, G. Ginestra and C. Bisignano, Biochemical and antimicrobial activity of phloretin and its glycosilated derivatives present in apple and kumquat, Food Chem., 2014, 160, 292–297 CrossRef CAS PubMed.
- R. Du, Q. Lv, W. Hu, X. Hou, Y. Zhou, X. Deng, L. Sun, L. Li, Y. Deng and J. Wang, Phloretin potentiates polymyxin E activity against gram-negative bacteria, Life Sci., 2021, 287, 120085 CrossRef CAS PubMed.
- S. Vasudevan, G. Thamil Selvan, S. Bhaskaran, N. Hari and A. P. Solomon, Reciprocal Cooperation of Type A Procyanidin and Nitrofurantoin Against Multi-Drug Resistant (MDR) UPEC: A pH-Dependent Study, Front. Cell. Infect. Microbiol., 2020, 10, 421 CrossRef CAS PubMed.
- S. Farooq, A. T. Wahab, C. D. Fozing, A. U. Rahman and M. I. Choudhary, Artonin I inhibits multidrug resistance in Staphylococcus aureus and potentiates the action of inactive antibiotics in vitro, J. Appl. Microbiol., 2014, 117, 996–1011 CrossRef CAS PubMed.
- C. A. N. Ferraz, S. R. Tintino, A. M. R. Teixeira, P. N. Bandeira, H. S. Santos, B. G. Cruz, C. E. S. Nogueira, T. F. Moura, R. L. S. Pereira, D. M. Sena, Jr., T. S. Freitas, J. E. Rocha and H. D. M. Coutinho, Potentiation of antibiotic activity by chalcone (E)-1-(4'-aminophenyl)-3-(furan-2-yl)-prop-2-en-1-one against gram-positive and gram-negative MDR strains, Microb. Pathog., 2020, 148, 104453 CrossRef CAS PubMed.
- J. K. Dzotam, I. K. Simo, G. Bitchagno, I. Celik, L. P. Sandjo, P. Tane and V. Kuete, In vitro antibacterial and antibiotic modifying activity of crude extract, fractions and 3',4',7-trihydroxyflavone from Myristica fragrans Houtt against MDR Gram-negative enteric bacteria, BMC Complementary Altern. Med., 2018, 18, 15 CrossRef PubMed.
- F. Taleghani, G. Rezvani, M. Birjandi and M. Valizadeh, Impact of green tea intake on clinical improvement in chronic periodontitis: A randomized clinical trial, J. Stomatol., Oral Maxillofac. Surg., 2018, 119, 365–368 CrossRef CAS PubMed.
- C. Astill, M. R. Birch, C. Dacombe, P. G. Humphrey and P. T. Martin, Factors affecting the caffeine and polyphenol contents of black and green tea infusions, J. Agric. Food Chem., 2001, 49, 5340–5347 CrossRef CAS PubMed.
- M. Radji, R. A. Agustama, B. Elya and C. R. Tjampakasari, Antimicrobial activity of green tea extract against isolates of methicillin–resistant Staphylococcus aureus and multi–drug resistant Pseudomonas aeruginosa, Asian Pac. J. Trop. Biomed., 2013, 3, 663–667 CrossRef CAS PubMed.
- E. D. Pauli, I. S. Scarminio and R. Tauler, Analytical investigation of secondary metabolites extracted from Camellia sinensis L. leaves using a HPLC-DAD-ESI/MS data fusion strategy and chemometric methods, J. Chemom., 2016, 30, 75–85 CrossRef CAS.
- L. H. Yao, Y. M. Jiang, N. Caffin, B. D’Arcy, N. Datta, X. Liu, R. Singanusong and Y. Xu, Phenolic compounds in tea from Australian supermarkets, Food Chem., 2006, 96, 614–620 CrossRef CAS.
- Y. S. Cho, N. L. Schiller and K. H. Oh, Antibacterial effects of green tea polyphenols on clinical isolates of methicillin-resistant Staphylococcus aureus, Curr. Microbiol., 2008, 57, 542–546 CrossRef CAS PubMed.
- T. S. S. S. Yam and J. M. Hamilton-Miller, Microbiological activity of whole and fractionated crude extracts of tea (Camellia sinensis), and of tea components, FEMS Microbiol. Lett., 1997, 152, 169–174 CrossRef CAS PubMed.
- P. D. Stapleton, S. Shah, J. C. Anderson, Y. Hara, J. M. Hamilton-Miller and P. W. Taylor, Modulation of beta-lactam resistance in Staphylococcus aureus by catechins and gallates, Int. J. Antimicrob. Agents, 2004, 23, 462–467 CrossRef CAS PubMed.
- P. D. Stapleton, S. Shah, J. M. Hamilton-Miller, Y. Hara, Y. Nagaoka, A. Kumagai, S. Uesato and P. W. Taylor, Anti-Staphylococcus aureus activity and oxacillin resistance modulating capacity of 3-O-acyl-catechins, Int. J. Antimicrob. Agents, 2004, 24, 374–380 CrossRef CAS PubMed.
- M. Kurincic, A. Klancnik and S. Smole Mozina, Effects of efflux pump inhibitors on erythromycin, ciprofloxacin, and tetracycline resistance in Campylobacter spp. isolates, Microb. Drug Resist., 2012, 18, 492–501 CrossRef CAS PubMed.
- R. Gaur and G. H. Bao, Chemistry and Pharmacology of Natural Catechins from Camellia sinensis as Anti-MRSA Agents, Curr. Top. Med. Chem., 2021, 21, 1519–1537 CrossRef CAS PubMed.
- L. Wang, Q. Li, J. Li, S. Jing, Y. Jin, L. Yang, H. Yu, D. Wang, T. Wang and L. Wang, Eriodictyol as a Potential Candidate Inhibitor of Sortase A Protects Mice From Methicillin-Resistant Staphylococcus aureus-Induced Pneumonia, Front. Microbiol., 2021, 12, 635710 CrossRef PubMed.
- W. Qian, W. Wang, J. Zhang, T. Wang, M. Liu, M. Yang, Z. Sun, X. Li and Y. Li, Antimicrobial and antibiofilm activities of ursolic acid against carbapenem-resistant Klebsiella pneumoniae, J. Antibiot., 2020, 73, 382–391 CrossRef CAS PubMed.
- I. Macedo, J. H. da Silva, P. T. da Silva, B. G. Cruz, J. P. C. do Vale, H. S. Dos Santos, P. N. Bandeira, E. B. de Souza, M. R. Xavier, H. D. M. Coutinho, R. Braz-Filho and A. M. R. Teixeira, Structural and Microbiological Characterization of 5-Hydroxy-3,7,4'-Trimethoxyflavone: A Flavonoid Isolated from Vitex gardneriana Schauer Leaves, Microb. Drug Resist., 2019, 25, 434–438 CrossRef CAS PubMed.
- Z. Cao, P. Yang and Q. Zhou, Multiple biological functions and pharmacological effects of lycorine, Sci. China: Chem., 2013, 56, 1382–1391 CrossRef CAS PubMed.
- N. T. Son and A. I. Elshamy, Flavonoids and other Non-alkaloidal Constituents of Genus Erythrina: Phytochemical Review, Comb. Chem. High Throughput Screening, 2021, 24, 20–58 CrossRef CAS PubMed.
- S. Lopez-Molina, C. Galiana-Rosello, C. Galiana, A. Gil-Martinez, S. Bandeira and J. Gonzalez-Garcia, Alkaloids as Photosensitisers for the Inactivation of Bacteria, Antibiotics, 2021, 10, 1505 CrossRef CAS PubMed.
- W. T. Beaulieu, D. G. Panaccione, Q. N. Quach, K. L. Smoot and K. Clay, Diversification of ergot alkaloids and heritable fungal symbionts in morning glories, Commun. Biol., 2021, 4, 1362 CrossRef PubMed.
- J. T. Wibowo, P. Ahmadi, S. I. Rahmawati, A. Bayu, M. Y. Putra and A. Kijjoa, Marine-Derived Indole Alkaloids and Their Biological and Pharmacological Activities, Mar. Drugs, 2021, 20, 3 CrossRef PubMed.
- F. Barbosa, E. Pinto, A. Kijjoa, M. Pinto and E. Sousa, Targeting antimicrobial drug resistance with marine natural products, Int. J. Antimicrob. Agents, 2020, 56, 106005 CrossRef CAS PubMed.
- K. I.-N. Karen Kubo, S. Ohnuki, Y. Yashiroda, S. C. Li, H. Kimura, Y. Kawamura, Y. Shimamoto, K. Tominaga, D. Yamanaka, Y. Adachi, S. Takashima, Y. Noda, C. Boone and Y. Ohya, Jerveratrum-Type Steroidal Alkaloids Inhibit b-1,6-Glucan Biosynthesis in Fungal Cell Walls, Microbiol. Spectrum, 2022, 10, e0087321 CrossRef PubMed.
- Y. S. Yang, X. Lu, Q. X. Zeng, J. Pang, T. Y. Fan, X. F. You, S. Tang, Y. X. Wang and D. Q. Song, Synthesis and biological evaluation of 7-substituted cycloberberine derivatives as potent antibacterial agents against MRSA, Eur. J. Med. Chem., 2019, 168, 283–292 CrossRef CAS PubMed.
- J. K. Y. Yap, S. Y. Y. Tan, S. Q. Tang, V. K. Thien and E. W. L. Chan, Synergistic Antibacterial Activity Between 1,4-Naphthoquinone and beta-Lactam Antibiotics Against Methicillin-Resistant Staphylococcus aureus, Microb. Drug Resist., 2021, 27, 234–240 CrossRef CAS PubMed.
- M. Wang, B. Ma, Y. Ni, X. Xue, M. Li, J. Meng, X. Luo, C. Fang and Z. Hou, Restoration of the Antibiotic Susceptibility of Methicillin-Resistant Staphylococcus aureus and Extended-Spectrum beta-Lactamases Escherichia coli Through Combination with Chelerythrine, Microb. Drug Resist., 2021, 27, 337–341 CrossRef CAS PubMed.
- W.-D. Qian, J. Huang, J.-N. Zhang, X.-C. Li, Y. Kong, T. Wang and Y.-D. Li, Antimicrobial and Antibiofilm Activities and Mechanism of Action of Chelerythrine Against Carbapenem-Resistant Serratia marcescens In Vitro, Microb. Drug Resist., 2021, 27, 1105–1116 CrossRef CAS PubMed.
- X. Li, Y. Song, L. Wang, G. Kang, P. Wang, H. Yin and H. Huang, A Potential Combination Therapy of Berberine Hydrochloride With Antibiotics Against Multidrug-Resistant Acinetobacter baumannii, Front. Cell. Infect. Microbiol., 2021, 11, 660431 CrossRef CAS PubMed.
- G. B. Manjunath, S. P. Awasthi, M. S. H. Zahid, N. Hatanaka, A. Hinenoya, E. Iwaoka, S. Aoki, T. Ramamurthy and S. Yamasaki, Piperine, an active ingredient of white pepper, suppresses the growth of multidrug-resistant toxigenic Vibrio cholerae and other pathogenic bacteria, Lett. Appl. Microbiol., 2022, 74, 472–481 CrossRef CAS PubMed.
- K. Alhanout, S. Malesinki, N. Vidal, V. Peyrot, J. M. Rolain and J. M. Brunel, New insights into the antibacterial mechanism of action of squalamine, J. Antimicrob. Chemother., 2010, 65, 1688–1693 CrossRef CAS PubMed.
- K. Tushar, M. K. S. Beuria and P. Dulal, Sanguinarine blocks cytokinesis in bacteria by inhibiting FtsZ assembly and bundling, Biochemistry, 2005, 44, 16584–16593 CrossRef PubMed.
- D. Crncevic, L. Krce, L. Mastelic, A. Maravic, B. Soldo, I. Aviani, I. Primozic, R. Odzak and M. Sprung, The mode of antibacterial action of quaternary N-benzylimidazole salts against emerging opportunistic pathogens, Bioorg. Chem., 2021, 112, 104938 CrossRef CAS PubMed.
- L. Z. M. Kamal, M. A. A. Adam, S. N. M. Shahpudin, A. N. Shuib, R. Sandai, N. M. Hassan, Y. Tabana, D. F. Basri, L. T. L. Than and D. Sandai, Identification of Alkaloid Compounds Arborinine and Graveoline from Ruta angustifolia (L.) Pers for their Antifungal Potential against Isocitrate lyase (ICL1) gene of Candida albicans, Mycopathologia, 2021, 186, 221–236 CrossRef CAS PubMed.
- H. Tian, Q.-J. Liu, J.-T. Wang and L. Zhang, Antimicrobial crinane-type alkaloids from the bulbs of Crinum latifolium, J. Asian Nat. Prod. Res., 2020, 23, 1023–1029 CrossRef PubMed.
- G. R. Dwivedi, R. Tyagi, Sanchita, S. Tripathi, S. Pati, S. K. Srivastava, M. P. Darokar and A. Sharma, Antibiotics potentiating potential of catharanthine against superbug Pseudomonas aeruginosa, J. Biomol. Struct. Dyn., 2018, 36, 4270–4284 CrossRef CAS PubMed.
- R. Amber, M. Adnan, A. Tariq, S. N. Khan, S. Mussarat, A. Hashem, A. A. Al-Huqail, A. F. Al-Arjani and E. F. Abd Allah, Antibacterial activity of selected medicinal plants of northwest Pakistan traditionally used against mastitis in livestock, Saudi J. Biol. Sci., 2018, 25, 154–161 CrossRef PubMed.
- G. R. Dwivedi, A. Maurya, D. K. Yadav, V. Singh, F. Khan, M. K. Gupta, M. Singh, M. P. Darokar and S. K. Srivastava, Synergy of clavine alkaloid 'chanoclavine' with tetracycline against multi-drug-resistant E. coli, J. Biomol. Struct. Dyn., 2019, 37, 1307–1325 CrossRef CAS PubMed.
- M. M. Rahman, M. S. Rahaman, M. R. Islam, M. E. Hossain, F. Mannan Mithi, M. Ahmed, M. Saldias, E. K. Akkol and E. Sobarzo-Sanchez, Multifunctional Therapeutic Potential of Phytocomplexes and Natural Extracts for Antimicrobial Properties, Antibiotics (Basel), 2021, 10, 1076 CrossRef CAS PubMed.
- N. Jubair, M. Rajagopal, S. Chinnappan, N. B. Abdullah, A. Fatima and A. Zarrelli, Review on the Antibacterial Mechanism of Plant-Derived Compounds against Multidrug-Resistant Bacteria (MDR), J. Evidence-Based Complementary Altern. Med., 2021, 2021, 1–30 CrossRef PubMed.
- A. C. Justino de Araujo, P. R. Freitas, C. Rodrigues Dos Santos Barbosa, D. F. Muniz, J. E. Rocha, A. C. Albuquerque da Silva, C. Datiane de Morais Oliveira-Tintino, J. Ribeiro-Filho, L. Everson da Silva, C. Confortin, W. D. Amaral, C. Deschamps, J. M. Barbosa-Filho, N. T. Ramos de Lima, S. R. Tintino and H. D. Melo Coutinho, GC-MS-FID characterization and antibacterial activity of the Mikania cordifolia essential oil and limonene against MDR strains, Food Chem. Toxicol., 2020, 136, 111023 CrossRef CAS PubMed.
- A. Gupta, E. Jeyakumar and R. Lawrence, Strategic approach of multifaceted antibacterial mechanism of limonene traced in Escherichia coli, Sci. Rep., 2021, 11, 13816 CrossRef CAS PubMed.
- C. R. Dos Santos Barbosa, J. R. Scherf, T. S. de Freitas, I. R. A. de Menezes, R. L. S. Pereira, J. F. S. Dos Santos, S. S. P. de Jesus, T. P. Lopes, Z. de Sousa Silveira, C. D. de Morais Oliveira-Tintino, J. P. S. Junior, H. D. M. Coutinho, S. R. Tintino and F. A. B. da Cunha, Effect of Carvacrol and Thymol on NorA efflux pump inhibition in multidrug-resistant (MDR) Staphylococcus aureus strains, J. Bioenerg. Biomembr., 2021, 53, 489–498 CrossRef CAS PubMed.
- A. Martins, A. Vasas, M. Viveiros, J. Molnar, J. Hohmann and L. Amaral, Antibacterial properties of compounds isolated from Carpobrotus edulis, Int. J. Antimicrob. Agents, 2011, 37, 438–444 CrossRef CAS PubMed.
- F. Hosseinkhani, F. Jabalameli, M. Banar, N. Abdellahi, M. Taherikalani, W. B. Leeuwen and M. Emaneini, Monoterpene isolated from the essential oil of Trachyspermum ammi is cytotoxic to multidrug-resistant Pseudomonas aeruginosa and Staphylococcus aureus strains, Rev Soc Bras Med Trop, 2016, 49, 172–176 CrossRef PubMed.
- V. K. Gupta, N. Tiwari, P. Gupta, S. Verma, A. Pal, S. K. Srivastava and M. P. Darokar, A clerodane diterpene from Polyalthia longifolia as a modifying agent of the resistance of methicillin resistant Staphylococcus aureus, Phytomedicine, 2016, 23, 654–661 CrossRef CAS PubMed.
- S. Singh, Z. Fatima and S. Hameed, Insights into the mode of action of anticandidal herbal monoterpenoid geraniol reveal disruption of multiple MDR mechanisms and virulence attributes in Candida albicans, Arch. Microbiol., 2016, 198, 459–472 CrossRef CAS PubMed.
- L. C. Rosario Rojas, C. A. José, J. V. Abraham, H. L. Walter, G. Lamas., C. Sarasara., H. G. Robert and B. H. Gerald, Aegicerin, the first oleanane triterpene with wide-ranging antimycobacterial activity, isolated from Clavija procera, J. Nat. Prod., 2006, 69, 845–846 CrossRef PubMed.
- S. Wei, Y. Yang, W. Tian, M. Liu, S. Yin and J. Li, Synergistic Activity of Fluoroquinolones Combining with Artesunate Against Multidrug-Resistant Escherichia coli, Microb. Drug Resist., 2020, 26, 81–88 CrossRef CAS PubMed.
- L. D. Hazlett, S. A. Ekanayaka, S. A. McClellan and R. Francis, Glycyrrhizin Use for Multi-Drug Resistant Pseudomonas aeruginosa: In Vitro and In Vivo Studies, Investig. Ophthalmol. Vis. Sci., 2019, 60, 2978–2989 CrossRef CAS PubMed.
- M. F. S. Patrícia Rijo, A. P. Francisco., R. Rojas., H. G. Robert, J. V. Abraham, B. Rodríguez. and C. Moiteiro., Antimycobacterial metabolites from Plectranthus: royleanone derivatives against Mycobacterium tuberculosis strains, Chem. Biodiversity, 2010, 7, 922–932 CrossRef PubMed.
- S. S. Swain, S. K. Paidesetty and R. N. Padhy, Development of antibacterial conjugates using sulfamethoxazole with monocyclic terpenes: A systematic medicinal chemistry based computational approach, Comput. Methods Programs Biomed., 2017, 140, 185–194 CrossRef PubMed.
- L. H. Yu, X. Wei, M. Ma, X. J. Chen and S. B. Xu, Possible inhibitory molecular mechanism of farnesol on the development of fluconazole resistance in Candida albicans biofilm, Antimicrob. Agents Chemother., 2012, 56, 770–775 CrossRef CAS PubMed.
- L. Cernakova, S. Dizova, D. Gaskova, I. Jancikova and H. Bujdakova, Impact of Farnesol as a Modulator of Efflux Pumps in a Fluconazole-Resistant Strain of Candida albicans, Microb. Drug Resist., 2019, 25, 805–812 CrossRef CAS PubMed.
- A. Sharma, A. Biharee, A. Kumar and V. Jaitak, Antimicrobial Terpenoids as a Potential Substitute in Overcoming Antimicrobial Resistance, Curr. Drug Targets, 2020, 21, 1476–1494 CrossRef CAS PubMed.
- H. Siddique, B. Pendry and M. M. Rahman, Terpenes from Zingiber montanum and Their Screening against Multi-Drug Resistant and Methicillin Resistant Staphylococcus aureus, Molecules, 2019, 24, 385 CrossRef PubMed.
- K. Oughlissi-Dehak, P. Lawton, S. Michalet, C. Bayet, N. Darbour, M. Hadj-Mahammed, Y. A. Badjah-Hadj-Ahmed, M. G. Dijoux-Franca and D. Guilet, Sesquiterpenes from aerial parts of Ferula vesceritensis, Phytochemistry, 2008, 69, 1933–1938 CrossRef CAS PubMed.
- N. M. Vazquez, F. Mariani, P. S. Torres, S. Moreno and E. M. Galvan, Cell death and biomass reduction in biofilms of multidrug resistant extended spectrum beta-lactamase-producing uropathogenic Escherichia coli isolates by 1,8-cineole, PLoS One, 2020, 15, e0241978 CrossRef CAS PubMed.
- H. C. de Carvalho, A. L. Ieque, T. L. Valverde, V. P. Baldin, J. E. Meneguello, P. A. Z. Campanerut-Sa, F. Vandresen, L. D. Ghiraldi Lopes, M. R. Passos Souza, N. C. S. Santos, V. L. Dias Siqueira, K. R. Caleffi-Ferracioli, R. B. Lima Scodro and R. F. Cardoso, Activity of (-)-Camphene Derivatives Against Mycobacterium tuberculosis in Acidic pH, Med. Chem., 2021, 17, 485–492 CrossRef CAS PubMed.
- V. L. Elodie Guinoiseau, L. Anne, T. Félix, C. Joseph and L. Berti., Susceptibility of the multi-drug resistant strain of Enterobacter aerogenes EA289 to the terpene alcohols from Cistus ladaniferus essential oil, Nat. Prod. Commun., 2011, 6, 1159–1162 Search PubMed.
- P. Knezevic, V. Aleksic, N. Simin, E. Svircev, A. Petrovic and N. Mimica-Dukic, Antimicrobial activity of Eucalyptus camaldulensis essential oils and their interactions with conventional antimicrobial agents against multi-drug resistant Acinetobacter baumannii, J. Ethnopharmacol., 2016, 178, 125–136 CrossRef CAS PubMed.
Footnote |
| † These authors contributed equally to this work. |
| This journal is © The Royal Society of Chemistry 2023 |