
Photoactivation of the cGAS-STING pathway and pyroptosis by an endoplasmic reticulum-targeting ruthenium(II) complex for cancer immunotherapy†
Bin-Fa
Liang‡
a,
Shan
Jiang‡
ab,
Yun-Shi
Zhi
a,
Zheng-Yin
Pan
 *b,
Xiu-Qing
Su
a,
Qin
Gong
a,
Zhen-Dan
He
b,
Da-Hong
Yao
b,
Liang
He
*c and
Chen-Yang
Li
*a
*b,
Xiu-Qing
Su
a,
Qin
Gong
a,
Zhen-Dan
He
b,
Da-Hong
Yao
b,
Liang
He
*c and
Chen-Yang
Li
*a
aSchool of Pharmacy, Shenzhen University Medical School, Shenzhen University, Shenzhen 518055, China. E-mail: lcy@szu.edu.cn
bCollege of Pharmacy, Shenzhen Technology University, Shenzhen 518118, China. E-mail: panzhengyin@sztu.edu.cn
cKey Laboratory for Biobased Materials and Energy of Ministry of Education, College of Materials and Energy, South China Agricultural University, Guangzhou 510642, China. E-mail: heliang@scau.edu.cn
First published on 31st January 2025
Abstract
Three multifunctional ruthenium(II) complexes (Ru1–Ru3) modified with cholic acid were synthesized, which exhibited excellent singlet oxygen-generating ability and near-infrared (NIR) aggregation-induced emission (AIE) phosphorescence activity. Cellular toxicity assays revealed that Ru1 displayed pronounced phototoxicity against both human breast cancer cells (MDA-MB-231) and murine breast cancer cells (4T1), achieving a maximum phototoxicity index (PI) of 83.3. Mechanistic studies indicated that Ru1 exhibited superior targeting affinity for the endoplasmic reticulum (ER). Upon irradiation at 450 nm, it stimulated the production of reactive oxygen species (ROS) and initiated ER stress. This stress activated the interferon gene stimulator (STING) pathway's signaling cascade within the ER, prompting a Golgi apparatus response. The consequent activation induced pyroptosis and sequentially engaged the downstream proteins p-TBK1 and p-IRF3 within the STING pathway, thus promoting the secretion of antitumor cytokines and the elicitation of tumor immune responses. In vivo experiments conducted on Balb/c mice have demonstrated significant anti-tumor immune effects exhibited by Ru1. In summary, the immune modulation and targeted intervention by metal complexes represent an innovative and promising therapeutic strategy for cancer. This approach is anticipated to yield new perspectives for the development of metal complexes that augment tumor immunotherapy.
Introduction
Conventional cancer drug therapy employs cytotoxicity to suppress cell growth and eliminate tumors.1 Currently, accumulating clinical evidence indicates that the clinical effects of traditional anticancer drugs depend not only on direct cytostatic and cytotoxic effects, but also on the reactivation of tumor immune responses.2 Immunotherapy is a promising therapeutic modality that selectively exploits the host's immune defenses against cancer.3 An effective immune response is generated following treatment and may trigger immunogenic cell death (ICD) of dying tumor cells to eliminate tumors.4 Some treatments (such as photodynamic therapy (PDT),5 high hydrostatic pressure,6 radiotherapy,7etc.) and drugs (such as chemotherapeutic agents,8 oncolytic viruses,9etc.) can provide or increase the immunogenicity. Most of the ICD inducers used in clinical tumor therapy are non-metallic compounds, such as doxorubicin,10 mitoxantrone,11 cyclophosphamide,12etc. Among the traditional metal anticancer drugs used clinically, oxaliplatin can induce ICD.13 Currently, some non-platinum transition metal complexes, such as arene binuclear ruthenium(II),14 palladium(II)15 and hetero-multinuclear gadolinium(III)–copper(II) complexes,16 have also been proven to be excellent inducers of ICD.In addition to killing tumor cells, ICD inducers can also release immunogenic signals through spatiotemporal synergy. Usually the immune system cannot pick up the signal of damage-associated molecular patterns (DAMPs), but DAMPs are released extracellularly or exposed on the cell membrane during cell death.13 DAMPs, as endogenous danger signals, include the exposure of calreticulin (CRT) on the cell surface, the migration of high mobility group box 1 (HMGB1) to the extracellular environment, the exocytosis of a large amount of ATP, etc., which in turn lead to the immunogenicity of tumor cells.17,18 The release or exposure of DAMPs can enhance the immunogenicity of tumor cells and promote the antigen presentation ability of dendritic cells (DCs), thereby triggering a series of T cell-dependent anti-tumor immune responses.19
Dysregulation of protein folding, export, and hydrolysis in cancer cells leads to endoplasmic reticulum (ER) stress. Drugs that further enhance this effect are also used as anticancer agents, including the clinically approved proteasome inhibitors bortezomib and carfilzomib. Studies have shown that cholic acid groups can enhance the targeting of compounds to the ER, such as the conjugates of cholic acid and N-alkylaminoferrocene-based prodrugs, accumulating in the ER of cancer cells and inducing ER stress, which weakens the mitochondrial membrane potential and generates mitochondrial reactive oxygen species (ROS), leading to cell death via necrosis.20 The coexistence of ER stress and excess ROS increases the class of different DAMPs, which play an important induction role in the ICD process.21 Due to the high (light) stability, high ROS yield, and easy modification of metal complexes, more and more laboratories are now committed to the development of metal complex photosensitizers that target the ER to induce ICD.
Pyroptosis is a distinct form of programmed cell death that is pro-inflammatory in nature and is critical for immunity. During pyroptosis, inflammatory caspase cleaves gasdermin D (GSDMD) and releases its N-terminal domain (GSDMD-N) to the cell membrane to bind with membrane phospholipids and drill holes in the cell membrane. Eventually, the osmotic potential of the membrane is disrupted, resulting in cell swelling, rupture of the cell membrane, and rapid release of inflammatory cytokines (e.g., IL-1β and IL-18). Thus, pyroptosis-induced inflammation can trigger strong antitumor immunity and enhance the efficacy of ICD.22 Shao Feng's team found that inducing pyroptosis in less than 15% of tumor cells was sufficient to clear the entire mouse mammary gland xenograft. This provides a new direction for the design of highly immunogenic photosensitizers.23
Interferon gene stimulator (STING) is a key signaling molecule in innate immune responses and plays a crucial role in anti-tumor immune reactions.24 As a very promising anti-cancer strategy, the STING pathway can be activated in various ways, such as the release of DNA,25 activation by inorganic Mn2+,26 and the use of small molecule activators like MSA-2![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 27 and SR717.28
27 and SR717.28
Ruthenium complexes are the most promising alternatives to clinical platinum-based antitumor drugs and show many unique properties that make them particularly useful in drug design, for example, better water solubility, higher cellular uptake, and rich physiochemical and biological properties.29,30 Ruthenium complexes are presently receiving great attention in the fields of biological, pharmaceutical and medicinal chemistry as anticancer agents. At this stage, several ruthenium complexes have entered into clinical trials, including NAMI-A, KP1019, KP1339 and one PDT agent, TLD143362. According to their chemical structures, ruthenium complexes, currently studied as anticancer candidates, are divided into two main groups: Ru(II)–arene complexes31–34 and Ru(II)–polypyridyl complexes.35–38
In this work, three multifunctional ruthenium(II) complexes Ru1–Ru3 with cholic acid as ligands were synthesized (Scheme 1), which had excellent singlet oxygen generation ability and near-infrared (NIR) aggregation-induced emission (AIE) phosphorescence activity. Through cytotoxicity screening experiments, it was found that Ru1 exhibited high phototoxicity to human breast cancer MDA-MB-231 cells and mouse breast cancer 4T1 cells, with a maximum phototoxicity index (PI) value of 83.3, so Ru1 was selected as our follow-up research object. Mechanism studies showed that Ru1 exhibited excellent ER targeting properties. After laser irradiation (450 nm), it generated ROS, induced ER stress, activated the signaling pool of the STING pathway in the ER, and then prompted a Golgi response. Stimulation led to pyroptosis and sequentially activated the downstream proteins p-TBK1 and p-IRF3 of the STING pathway to produce anti-tumor factors and caused tumor ICD. In vivo experiments showed that Ru1 also had obvious in vivo anti-tumor effects in Balb/c mice. It effectively initiated GSDMD-mediated pyroptosis and triggered ICD. What's more, the antitumor immune-therapeutic effect of Ru1 was verified in a mouse tumor model.
 | ||
| Scheme 1 Chemical structures of Ru1–Ru3. | ||
Results and discussion
Synthesis and characterization
The synthesis route of complexes Ru1–Ru3 is shown in Scheme S1.† Ligand Phen-CA was synthesized as previously reported.39 Complexes Ru1–Ru3 were synthesized according to the conventional methods. Complexes Ru1–Ru3 were characterized by ESI-MS, 1H NMR spectroscopy, 13C NMR spectroscopy and HPLC purity analysis (Fig. S1–S12†).Photophysical and AIE properties
The UV-Vis absorption spectra of Ru1–Ru3 exhibited both strong intra-ligand transition bands at 250–300 nm and a weaker broad band at 350–500 nm (Fig. 1A and S13A, B†), which were attributed to metal–ligand charge transfer (1MLCT). Due to the conjugation of carbon–nitrogen ligands, complexes Ru1–Ru3 have a strong absorption at 450 nm. The emission spectra of Ru1–Ru3 in CH3CN, CH2Cl2 and PBS were measured at 298 K (Fig. 1B and S13C, D†). Complexes Ru1–Ru3 exhibit the largest emission in the near-infrared region (about 550–700 nm), which is attributed to the 3MLCT excited state of the complexes. | ||
| Fig. 1 (A) UV/Vis and (B) emission spectra of Ru1–Ru3 (2 × 10−5 M) recorded in PBS at 298 K. (C) Changes in the absorption spectra of 1,3-diphenylisobenzofuran (DPBF) at 410 nm upon irradiation at 450 nm in the presence of Ru1–Ru3 in aerated DMSO. [Ru(bpy)3]Cl2 was used as a standard. (D) The ESR spectra of 1O2 trapped by 2,2,6,6-tetramethylpiperidine (TEMP) in different groups. | ||
Using 1,3-diphenylisobenzofuran (DPBF) as a singlet oxygen probe, the ability of Ru1–Ru3 to generate singlet oxygen under light conditions was evaluated. A decrease in the DPBF absorption maximum indicates the generation of singlet oxygen. As shown in Fig. 1C, in the control experiment, DPBF hardly undergoes photooxidation under light. However, in the presence of Ru1–Ru3, under the irradiation of 450 nm light, the absorption maximum of DPBF at 410 nm gradually decreased, indicating the generation of 1O2. Using [Ru(bpy)3]Cl2 as a reference (ΦΔ = 0.66 in DMSO),40 the singlet oxygen quantum yields (ΦΔ) of Ru1–Ru3 in aerated DMSO were calculated to be 0.79, 0.81, and 0.89, respectively. Electron spin resonance (ESR) spectroscopy was further employed to verify the generation of ROS (Fig. 1D and S14†). The results demonstrated that the reactive oxygen species generated by the three complexes upon light irradiation are all singlet oxygen, with no superoxide radicals or hydroxyl radicals present. Ru1–Ru3 can effectively photosensitively generate 1O2, indicating their potential application value in PDT. The AIE properties of Ru1–Ru3 were investigated in a mixed solution of water and CH3CN with the water content ranging from 0 to 90% (v/v) (Fig. S15†). Initially, Ru1–Ru3 had weak emission in pure CH3CN, but it gradually increased with the increase of water content, exhibiting typical AIE characteristics. Specifically, they emitted gradually enhanced near-infrared phosphorescence. Furthermore, the photostability of Ru1 was investigated by UV-Vis absorption spectroscopy. The absorption spectra of complexes Ru1–Ru3 dissolved in phosphate buffered saline (PBS) showed no change over time (Fig. S16†), and the absorption spectrum of complex Ru1 also showed little change under light (Fig. S17†), indicating that Ru1 had high photostability and could be further used in experiments under light conditions.
PDT activity
The photodynamic activity of Ru1–Ru3 on MDA-MB-231, 4T1 and HLF cells was detected using the tetramethylazolium salt (MTT) method, with cisplatin as a reference. As shown in Table 1, after Ru1–Ru3 were incubated in the dark for 48 h, the IC50 values of Ru1 and Ru2 were greater than 100 μM, indicating that Ru1 and Ru2 had lower cytotoxicity to MDA-MB-231 and 4T1 cells in the dark, and the dark toxicity of Ru3 was higher than those of Ru1 and Ru2. After irradiating with a 450 nm (15 mW cm−2) laser for 15 min and subsequently incubating for 36 h, complexes Ru1–Ru3 showed obvious phototoxicity to MDA-MB-231 and 4T1 cells with an IC50 value of less than 8.5 μM, while the IC50 values of Ru3 under light were all less than 0.2 μM, and the PI values were all greater than 10, among which the PI value of Ru1 reached 83.3. Light alone had no effect on cell viability (Fig. S18†). The results indicate that the PDT activity of these Ru(II) complexes towards normal cells is lower than that towards cancer cells, especially for Ru1, suggesting that Ru1 has better selectivity towards cancer cells. These results indicated that Ru1–Ru3 had good PDT activity in vitro, in particular, Ru1 had low dark toxicity and superior phototoxicity.| Cell condition | MDA-MB-231 | PIc | 4T1 | PIc | HLF | PIc | |||
|---|---|---|---|---|---|---|---|---|---|
| Darka | Lightb | Darka | Lightb | Darka | Lightb | ||||
| a Cells were incubated with the indicated compounds in the dark for 48 h. b Cells were incubated with the indicated compounds for 12 h in the dark and then irradiated with light at 450 nm. c PI = Phototoxicity index, PI is the ratio of the IC50 values in the dark to those obtained upon light irradiation. | |||||||||
| Ru1 | >100 | 1.2 ± 0.4 | >83.3 | >100 | 1.7 ± 0.5 | >58.8 | >100 | 3.4 ± 0.6 | >29.4 |
| Ru2 | >100 | 8.5 ± 0.9 | >11.8 | >100 | 2.4 ± 0.7 | >41.6 | >100 | 10.9 ± 0.9 | >9.17 |
| Ru3 | 10.5 ± 1.2 | 0.1 ± 0.06 | 105 | 7.9 ± 0.7 | 0.2 ± 0.08 | 39.5 | 13.3 ± 0.8 | 0.22 ± 0.05 | 60.5 |
| Cisplatin | 28.5 ± 2.3 | 27.5 ± 1.8 | 15.8 ± 1.4 | ||||||
ROS generation
It has been widely reported in the literature that PDT activity depends on the level of ROS generated by photoactivation of photosensitizers. Therefore, we used dichlorofluorescein diacetate (DCFH-DA) as a capture probe for ROS to detect the ability of Ru1 to induce ROS production in MDA-MB-231 cells under light irradiation for different durations. Cell fluorescence imaging results showed that in the dark group, the intracellular fluorescence was weaker. However, the fluorescence intensity of the cells under light for 5, 10 and 15 minutes increased gradually (Fig. 2A). Flow cytometry was used to further quantitatively detect the ability of Ru1 to generate ROS in cells under light irradiation for different periods (Fig. 2B). These results indicated that Ru1 could effectively induce cells to produce ROS under light irradiation, and as the light irradiation time increased, more ROS was produced. | ||
| Fig. 2 (A) Confocal microscopy and (B) flow cytometry detection of the intracellular ROS generation ability of complex Ru1 (1.2 μM) mediated by PDT. Scale bar: 20 μm. | ||
Cellular localization and ER stress
In cells, the ER is responsible for the biosynthesis, folding, maturation, stabilization and transport of transmembrane and secreted proteins, and is an interconnected subcellular structure. Studies have shown that ROS can destroy the folding ability of the ER and then lead to the accumulation of unfolded or misfolded proteins in the ER, that is, ER stress. Severe and persistent ER stress is closely related to tumor development, angiogenesis, invasiveness, and immunosuppression.41 Therefore, in order to explore the relationship between Ru1 and the ER in anti-tumor mechanisms, confocal laser microscopy was used to determine the localization of Ru1 in cells. As shown in Fig. 3A, the green fluorescence emitted by the complex itself and the red fluorescence of the ER commercial probe (ER-Tracker Red) were highly overlapped, and the Pearson's Correlation Coefficient (PCC) value reached 0.91, indicating that Ru1 preferentially accumulated in the ER, which was an important target of Ru1-mediated cell death. Some lipophilic ruthenium complexes have also been reported to target the ER.42–44 Andriy Mokhir's team has reported that N-alkylaminoferrocene prodrugs modified with cholic acid effectively target the ER of cells.45 Consequently, we hypothesize that the ER targeting capabilities of Ru1 may be linked to both the lipophilicity of the complex and the presence of the cholic acid group. | ||
| Fig. 3 ER localization of complex Ru1 and induction of ER stress. (A) Co-localization experiment of Ru1 (10 μM, 4 h; Ex = 405 nm and Em = 550–650 nm) in MDA-MB-231 with ER-Tracker Red (1 μM, 30 min; Ex = 633 nm and Em = 660–700 nm). Scale bar: 20 μm. (B) The protein expression levels of CHOP, γ-H2AX and p-STAT3 in cells under light irradiation for different times after adding complex Ru1. | ||
At the same time, western blotting showed that Ru1 increased the expression of ER stress-related protein (CHOP) and decreased the expression of phosphorylated signal transducer and activator of transcription 3 (p-STAT3) with increasing light exposure time (Fig. 3B and S19†). p-STAT3 inhibition can promote the expression of CHOP.46 That is, Ru1 produced a large amount of ROS in the ER after light irradiation and then mediated the ER stress response through p-STAT3. In addition, Ru1 promoted the expression of DNA damage marker γ-H2AX after light irradiation, indicating that Ru1 was a multi-target synergistic complex in cells.
Pyroptosis induction
The Ru1-induced cell death pattern under PDT treatment was studied through a variety of experiments. During the PDT process of Ru1, under a light microscope, most of the dead cells did not float like apoptotic cells, but were tightly attached to the culture plate, and there were a large number of vesicles around the cells (Fig. 4A). These morphological features were similar to pyroptosis. Cell transmission electron microscopy showed that Ru1 did not show apoptotic features such as cell shrinkage, nuclear fragmentation, or apoptotic body formation during PDT. In contrast, dead cells bulged large bubbles from the plasma membrane, and the whole cell underwent typical swelling, while the nucleus remained intact (Fig. 4B), suggesting that PDT of Ru1 could induce pyroptosis in MDA-MB-231 cells. According to literature reports, pyroptosis is the programmed necrosis of cells mediated by the gasdermin family, through the inflammasome to effectively activate some proteins of the caspase family, thereby cleaving the gasdermin family proteins into GSDMD-N and GSDMD-C, and activated GSDMD. The three-dimensional structure of GSDMD-N will change, transfer and insert into the cell membrane to form a hole, and this hole is inserted into the cell membrane by more than 20 GSDMD-N small molecules hand in hand, thereby promoting cell swelling and cytoplasm outflow, leading to rupture of the cell membrane, and ultimately cell pyroptosis. To further identify this mode of death, we detected the expression of GSDMD-N by western blotting. As shown in Fig. 4C and S20,† the expression of GSDMD-N in Ru1-treated cells was significantly increased after 450 nm laser irradiation, while the protein expression of GSDMD was reduced, which jointly verified that Ru1-mediated PDT induced cells to cleave GSDMD (GSDMD-N), perforating cell membranes and triggering pyroptosis to kill cancer cells. | ||
| Fig. 4 Ru1 induces pyroptosis. (A) The morphological changes of MDA-MB-231 cells treated with Ru1 + light under a light microscope, scale bar: 20 μm. (B) TEM pictures of MDA-MB-231 cells treated with Ru1 + light, scale bar: 2.5 μm. The red box indicates the enlarged area, scale bar: 0.5 μm. (C) The protein expression levels of GSDMD and GSDMD-N after Ru1 + light treatment of MDA-MB-231 cells. | ||
Immunogenic death induction
ICD in cancer therapy refers to an immunostimulatory pattern of cell death that may result from tumor vaccination, radiation therapy, and certain types of chemotherapy. Studies have shown that exposure of cell surface calreticulin (CRT), release of high mobility group box 1 (HMGB1), and exocytosis of intracellular ATP are the most fundamental features of ICD. Therefore, we used cell immunofluorescence imaging technology to study the immunogenicity characteristics of Ru1 (Fig. 5A–C). After 450 nm laser light irradiation, the green fluorescence of CRT cells under dark conditions was negligible, but the green fluorescence gradually increased. The enhancement indicated that Ru1 + light induced high expression of CRT. Under the same experimental conditions, the red fluorescence of HMGB1 decreased continuously with the increase of the illumination time, and the red fluorescence was negligible after 15 min of 450 nm laser irradiation, indicating that HMGB1 was released upon treating the cells with Ru1 + light. Large leakage of HMGB1 leads to a gradual decrease of intracellular HMGB1. In addition, the concentration of ATP in the supernatant of cells treated with Ru1 + light increased, indicating that the concentration of ATP secreted to the extracellular space increased. These results indicated that Ru1 + light produced a large amount of ROS in the ER to induce pyroptosis, the integrity of the plasma membrane was destroyed, and a series of DAMPs were released, which greatly enhanced the immunogenicity of cancer cells.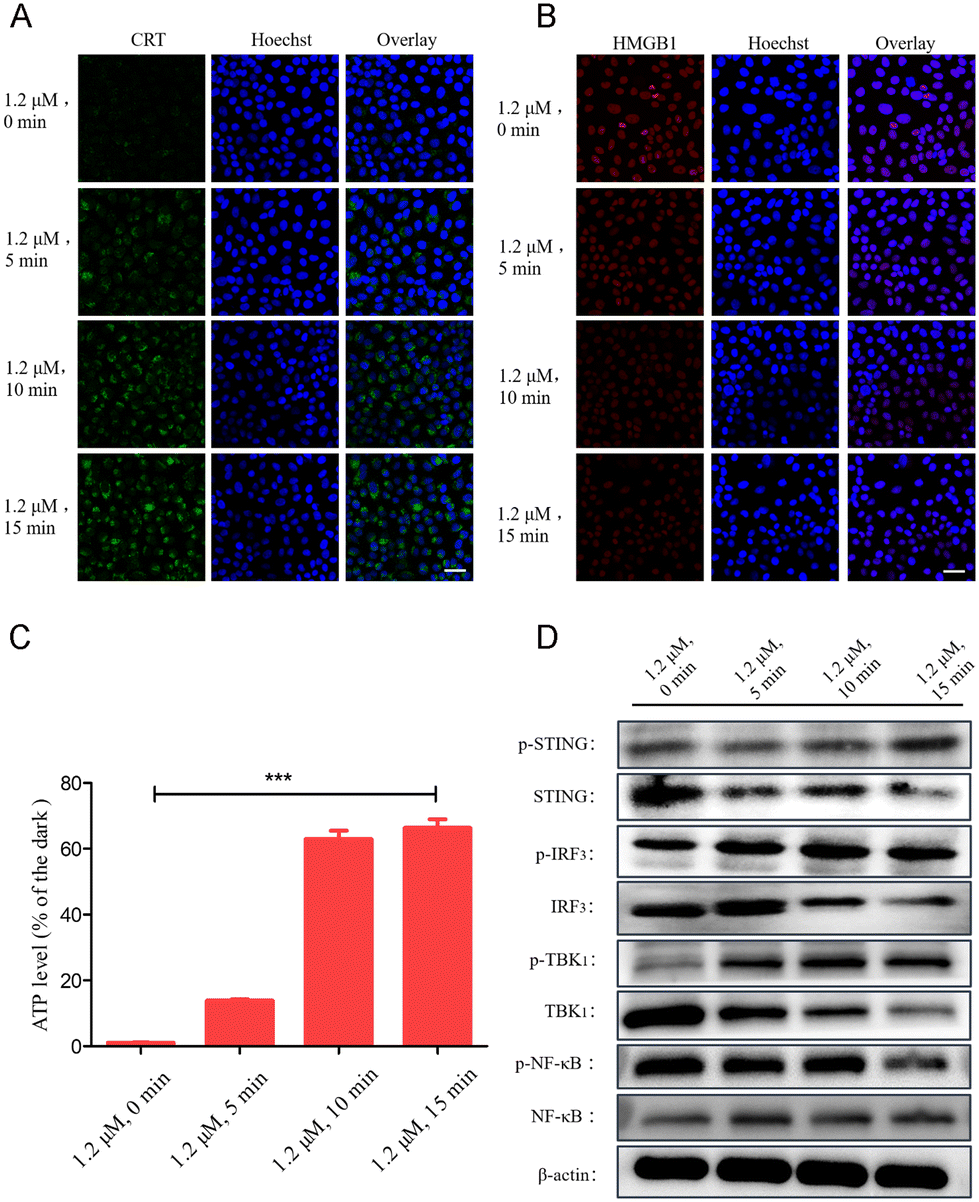 | ||
| Fig. 5 Cell immunofluorescence images of ICD markers CRT and HMGB1. The protein expression levels of (A) CRT and (B) HMGB1 after the cells were treated with Ru1 (1.2 μM) for 12 h under light irradiation for different durations. Scale bar: 20 μm. (C) Determination of the cellular immunogenicity marker ATP (extracellular). (D) Protein expression levels of p-STING, STING, p-IRF3, IRF3, p-TBK1, TBK1, p-NF-κB, and NF-κB in the STING pathway after Ru1 + light treatment of MDA-MB-231 cells. | ||
The cGAS-STING pathway has a critical role in antitumor immune responses, motivating the use of this natural defense pathway to enhance antitumor immunity in next-generation cancer immunotherapy.47 Studies have shown that stimuli other than cGAMP (cyclic GMP-AMP), including cyclic dinucleotides (CDNs) and ER stress, can also activate STING.48,49 Upon activation, STING is translocated from the ER to the Golgi apparatus, where it recruits kinases such as TBK1 and IκB kinase (IKK), which phosphorylate interferon regulatory factor 3 (IRF3) and the NF-κB inhibitor IκBα, respectively. Phosphorylated IRF3 dimerizes and translocates into cells to activate tumor immunity. However, during the PDT process of Ru1, the protein expression levels of p-STING, p-IRF3, p-TBK1, and NF-κB increased with the increase of light irradiation time (Fig. 5D and S21†). That is, Ru1 quickly accumulates in the ER of the cells after being added to the culture medium, and a large amount of ROS is produced after light irradiation, which damages the ER and causes an increase in the pressure of the ER, thereby activating the STING pathway in the ER signaling pool, leading to cellular immunogenic death.
In vivo anti-tumor evaluation
In order to study the anti-tumor efficacy and systemic immunity of Ru1 + light in vivo, we investigated the anti-tumor activity on the transplanted tumor model of Balb/c mice. 4–7 days after the tumor was implanted in mice, the drug was administered when the tumor volume reached 50–100 mm3, and the drug concentration was 5 mg kg−1. The mice treated with PBS + light/dark and Ru1 dark conditions were used as control mice. The tumor volumes of the mice in each group and the body weights of the mice were measured every 2 days. The mice were photographed and dissected after continuing to observe for 7 days after three doses of drugs, and the tumors were taken out. The results showed that Ru1 had an obvious inhibitory effect on mouse 4T1 xenograft tumors in vivo under light (Fig. 6A and B). Flow cytometric analysis showed that Ru1 promoted the maturation of dendritic cells (DC) in tumor-draining lymph nodes. Compared with the control group, the ratio of CD80+/CD86+ dendritic cells in the Ru1 + PDT group was significantly increased (Fig. 6C and D). Then flow cytometry was used to detect toxic T cells (CD8+) and helper T cells (CD4+) at the tumor site. Compared with the control group, the ratio of CD4+/CD8+ T cells in the Ru1 + light group was significantly increased (Fig. 6E–G). Consistent with the results of flow cytometry, this indicated that Ru1 had the ability to induce cytotoxic T lymphocytes to activate the adaptive immune response.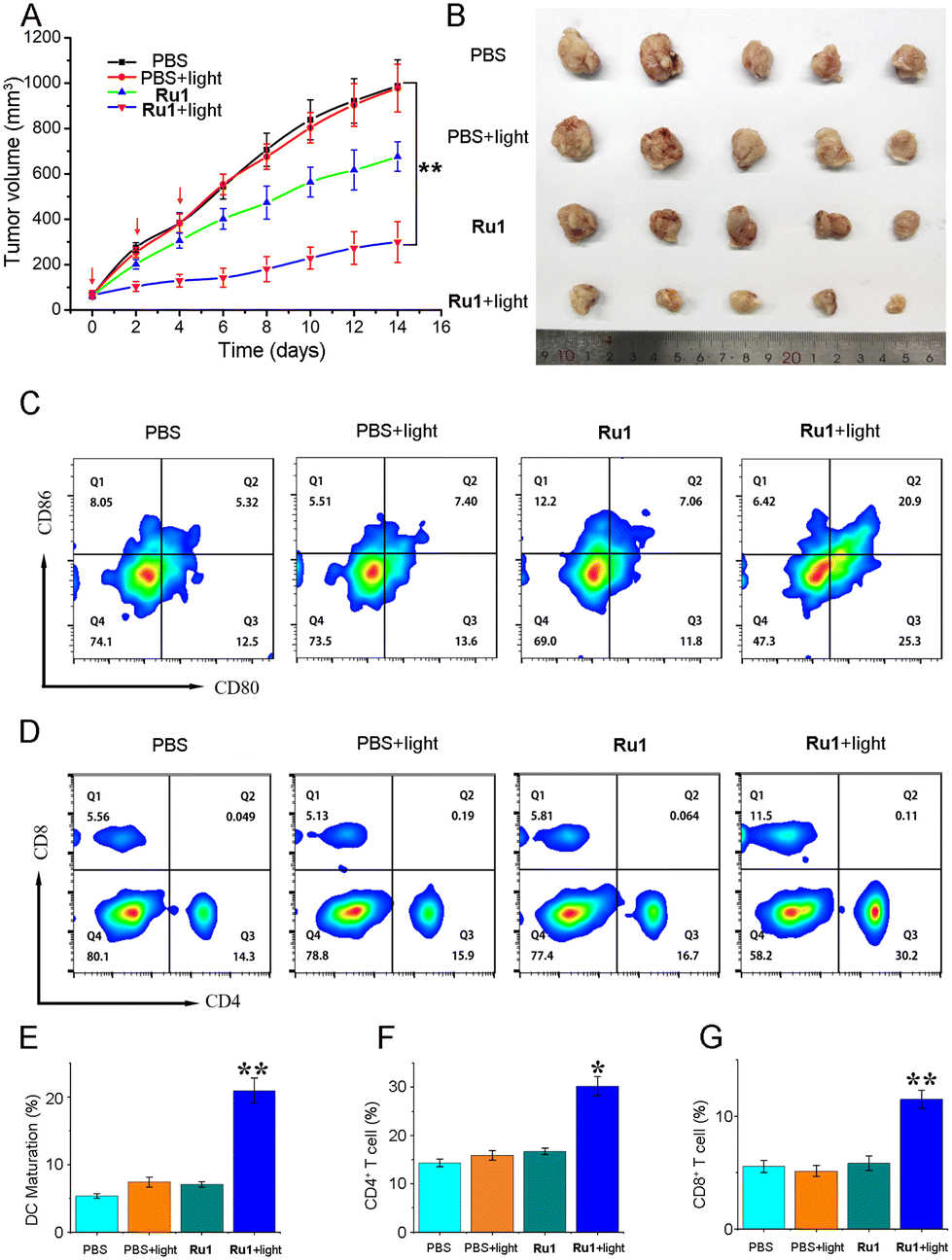 | ||
| Fig. 6 Verification of the anti-tumor activity of Ru1in vivo. (A) Tumor volume changes in mice treated with phosphate buffered saline (PBS) and Ru1 combined with PDT. (B) Tumors isolated from mice after PDT. (C and E) Expression of CD80+ and CD86+ quantitatively detected by flow cytometry in vivo (n = 3). (D, F, and G) Populations of CD4+ and CD8+ T cells in tumors (n = 3). *p < 0.05, **p < 0.01, and ***p < 0.001, using a Student's two-tailed t-test. | ||
In addition, during the process of PDT, the body weight of mice in each group did not change significantly (Fig. S23†), and the results of hematoxylin–eosin (H&E) staining showed no obvious pathological abnormalities in the tissues of the main organs (heart, liver, spleen, lungs, and kidneys) of mice (Fig. S24†), which proved that Ru1 also had good anti-tumor activity in animals and had low toxic side effects.
Conclusions
In summary, we have developed three innovative ruthenium(II) complexes Ru1–Ru3 that exhibit exceptional photophysical properties and AIE characteristics. Notably, Ru1 stands out for its ability to efficiently generate singlet oxygen under visible light irradiation at 450 nm, coupled with its remarkable photostability. Colocalization imaging studies reveal that Ru1 selectively accumulates within the ER. Upon exposure to visible light at 450 nm, Ru1 demonstrates superior phototoxicity against both human breast cancer MDA-MB-231 cells and mouse breast cancer 4T1 cells, achieving the highest PI values. Mechanistic investigations have elucidated that Ru1, upon light irradiation, triggers overproduction of ROS in MDA-MB-231 cells, leading to ER stress. This stress activates the STING pathway's signaling cascade within the ER, subsequently inducing Golgi stress and culminating in pyroptosis. Furthermore, Ru1 sequentially activates the downstream proteins p-TBK1 and p-IRF3 within the STING pathway, thereby stimulating the production of antitumor factors and initiating tumor immunogenic cell death. In addition to its in vitro effects, Ru1 demonstrated significant in vivo antitumor activity in Balb/c mice. In summary, the results demonstrate the potential of Ru1 as a promising photosensitizer for targeted ER imaging and for the photodynamic therapy of tumors.Author contributions
Bin-Fa Liang: investigation, formal analysis, and writing – original draft. Shan Jiang: investigation, formal analysis, and writing – original draft. Yun-Shi Zhi: investigation. Zheng-Yin Pan: conceptualization, funding acquisition, writing – review & editing, and supervision. Xiu-Qing Su: investigation. Qin Gong: investigation. Zhen-Dan He: funding acquisition. Da-Hong Yao: funding acquisition. Liang He: resources and writing – review & editing. Chen-Yang Li: resources and funding acquisition.Data availability
The data supporting this article have been included as part of the ESI.†Conflicts of interest
There are no conflicts to declare.Acknowledgements
This work was supported by the National Natural Science Foundation of China (22477081 and 22107074), the Guangdong Basic and Applied Basic Research Foundation (2024A1515030157, 2022A1515011356, and 2021A1515110211), the Innovation Team Project of the Guangdong Provincial Department of Education (2024KCXTD012), the Shenzhen Natural Science Fund (the Stable Support Plan Program 20220811110339002), the Shenzhen Science and Technology Innovation Program (JCYJ20230808105913028), the Shenzhen Longgang District Science and Technology Innovation Bureau (LGKCYLWS2021000001), and the Natural Science Foundation of Top Talent of SZTU (GDRC202416).References
- C. Martínez-Campa and C. Alonso-González, Editorial for the Special Issue “New Strategies in Cancer Pharmacotherapy: Development of Hormonal Antineoplastic Drugs, Cytotoxic Drugs and Targeted Therapies”, Int. J. Mol. Sci., 2020, 21, 4081 CrossRef PubMed.
- W. Jiang, Y. Wang, J. A. Wargo, F. F. Lang and B. Y. S. Kim, Considerations for designing preclinical cancer immune nanomedicine studies, Nat. Nanotechnol., 2020, 16, 6–15 CrossRef PubMed.
- X. Li, The inducers of immunogenic cell death for tumor immunotherapy, Tumori J., 2018, 104, 1–8 CrossRef CAS PubMed.
- X. Zhao, Q. Liu, C. Zheng, Y. Wang, Y. Zhao, Z. Zhang, J. Hao, Y. Zheng, C. Wang and Y. Liu, Tumor cell surface modification with immuno-amplified nanoparticles to enhance cancer immunotherapy, Mater. Today Chem., 2023, 27, 101303 CrossRef CAS.
- Q. Xiang, C. Yang, Y. Luo, F. Liu, J. Zheng, W. Liu, H. Ran, Y. Sun, J. Ren and Z. Wang, Near-Infrared II Nanoadjuvant-Mediated Chemodynamic, Photodynamic, and Photothermal Therapy Combines Immunogenic Cell Death with PD-L1 Block ade to Enhance Antitumor Immunity, Small, 2022, 18, 2107809 CrossRef CAS PubMed.
- I. Moserova, I. Truxova, A. D. Garg, J. Tomala, P. Agostinis, P. F. Cartron, S. Vosahlikova, M. Kovar, R. Spisek and J. Fucikova, Caspase-2 and oxidative stress underlie the immunogenic potential of high hydrostatic pressure-induced cancer cell death, OncoImmunology, 2016, 6, e1258505 CrossRef PubMed.
- H. Zhou, C. Tu, P. Yang, J. Li, O. Kepp, H. Li, L. Zhang, L. Zhang, Y. Zhao, T. Zhang, C. Sheng and J. Wang, Carbon ion radiotherapy triggers immunogenic cell death and sensitizes melanoma to anti-PD-1 therapy in mice, OncoImmunology, 2022, 11, 2057892 CrossRef PubMed.
- C. Groer, T. Zhang, R. Lu, S. Cai, D. Mull, A. Huang, M. Forrest, C. Berkland, D. Aires and M. L. Forrest, Intratumoral Cancer Chemotherapy with a Carrier-Based Immunogenic Cell-Death Eliciting Platinum(IV) Agent, Mol. Pharmaceutics, 2020, 17, 4334–4345 CrossRef CAS PubMed.
- L. Palanivelu, C.-H. Liu and L.-T. Lin, Immunogenic cell death: The cornerstone of oncolytic viro-immunotherapy, Front. Immunol., 2023, 13, 1038226 CrossRef PubMed.
- H. Zhu, W. Zhou, Y. Wan, K. Ge, J. Lu and C. Jia, Nanomedicine-mediated induction of immunogenic cell death and prevention of PD-L1 overexpression for enhanced hepatocellular carcinoma therapy, Cancer Nanotechnol., 2020, 11, 15 Search PubMed.
- C. Li, H. Sun, W. Wei, Q. Liu, Y. Wang, Y. Zhang, F. Lian, F. Liu, C. Li, K. Ying, H. Huo, Z. Qi and B. Li, Mitoxantrone triggers immunogenic prostate cancer cell death via p53-dependent PERK expression, Cell. Oncol., 2020, 43, 1099–1116 CrossRef CAS PubMed.
- G. Schiavoni, A. Sistigu, M. Valentini, F. Mattei, P. Sestili, F. Spadaro, M. Sanchez, S. Lorenzi, M. T. D'Urso, F. Belardelli, L. Gabriele, E. Proietti and L. Bracci, Cyclophosphamide Synergizes with Type I Interferons through Systemic Dendritic Cell Reactivation and Induction of Immunogenic Tumor Apoptosis, Cancer Res., 2011, 71, 768–778 CrossRef CAS PubMed.
- D. Groza, S. Gehrig, P. Kudela, M. Holcmann, C. Pirker, C. Dinhof, H. H. Schueffl, M. Sramko, J. Hoebart, F. Alioglu, M. Grusch, M. Ogris, W. Lubitz, B. K. Keppler, I. Pashkunova-Martic, C. R. Kowol, M. Sibilia, W. Berger and P. Heffeter, Bacterial ghosts as adjuvant to oxaliplatin chemotherapy in colorectal carcinomatosis, OncoImmunology, 2018, 7, e1424676 Search PubMed.
- G. Xu, Q. Liang, L. Gao, S. Xu, W. Luo, Q. Wu, J. Li, Z. Zhang, H. Liang and F. Yang, Developing an Arene Binuclear Ruthenium(II) Complex to Induce Ferroptosis and Activate the cGAS-STING Pathway: Targeted Inhibiting Growth and Metastasis of Triple Negative Breast Cancer, J. Med. Chem., 2024, 67, 19573–19585 CrossRef CAS PubMed.
- W. Li, S. Li, Z. Zhang, G. Xu, X. Man, F. Yang and H. Liang, Developing a Multitargeted Anticancer Palladium(II) Agent Based on the His-242 Residue in the IIA Subdomain of Human Serum Albumin, J. Med. Chem., 2023, 66, 8564–8579 CrossRef CAS PubMed.
- X. Man, W. Li, M. Zhu, S. Li, G. Xu, Z. Zhang, H. Liang and F. Yang, Rational Design of a Hetero-multinuclear Gadolinium(III)-Copper(II) Complex: Integrating Magnetic Resonance Imaging, Photoacoustic Imaging, Mild Photothermal Therapy, Chemotherapy and Immunotherapy of Cancer, J. Med. Chem., 2024, 67, 15606–15619 Search PubMed.
- S. von Hofsten, M. H. Paulsen, S. N. Magnussen, D. Ausbacher, M. Kranz, A. Bayer, M. B. Strøm and G. Berge, The marine natural product mimic MPM-1 is cytolytic and induces DAMP release from human cancer cell lines, Sci. Rep., 2022, 12, 15586 Search PubMed.
- R. Fan, A. Deng, R. Lin, S. Zhang, C. Cheng, J. Zhuang, Y. Hai, M. Zhao, L. Yang and G. Wei, A platinum(IV)-artesunate complex triggers ferroptosis by boosting cytoplasmic and mitochondrial lipid peroxidation to enhance tumor immunotherapy, Med. Commun., 2024, 5, e570 CAS.
- J. Deng, W. Xu, S. Lei, W. Li, Q. Li, K. Li, J. Lyu, J. Wang and Z. Wang, Activated Natural Killer Cells-Dependent Dendritic Cells Recruitment and Maturation by Responsive Nanogels for Targeting Pancreatic Cancer Immunotherapy, Small, 2022, 18, 2203114 CrossRef CAS PubMed.
- H. G. Xu, M. Schikora, M. Sisa, S. Daum, I. Klemt, C. Janko, C. Alexiou, G. Bila, R. Bilyy, W. Gong, M. Schmitt, L. Sellner and A. Mokhir, An Endoplasmic Reticulum Specific Pro-amplifier of Reactive Oxygen Species in Cancer Cells, Angew. Chem., Int. Ed., 2021, 60, 11158–11162 CrossRef CAS PubMed.
- E.-K. Kim, Y. Kim, J. Y. Yang and H. H. Jang, Prx1 Regulates Thapsigargin-Mediated UPR Activation and Apoptosis, Genes, 2022, 13, 2033 CrossRef CAS PubMed.
- M. Wang, M. Wu, X. Liu, S. Shao, J. Huang, B. Liu and T. Liang, Pyroptosis Remodeling Tumor Microenvironment to Enhance Pancreatic Cancer Immunotherapy Driven by Membrane Anchoring Photosensitizer, Adv. Sci., 2022, 9, 2202914 CrossRef PubMed.
- Q. Wang, Y. Wang, J. Ding, C. Wang, X. Zhou, W. Gao, H. Huang, F. Shao and Z. Liu, A bioorthogonal system reveals antitumour immune function of pyroptosis, Nature, 2020, 579, 421–426 CrossRef CAS PubMed.
- R. Fan, R. Lin, S. Zhang, A. Deng, Y. Hai, J. Zhuang, Y. Liu, M. Cheng and G. Wei, Novel Pt(IV) complex OAP2 induces STING activation and pyroptosis via mitochondrial membrane remodeling for synergistic chemo-immunotherapy, Acta Pharm. Sin. B, 2024, 14, 1742–1758 CrossRef CAS PubMed.
- Y.-Y. Ling, Z.-Y. Li, X. Mu, Y.-J. Kong, L. Hao, W.-J. Wang, Q.-H. Shen, Y.-B. Zhang and C.-P. Tan, Self-assembly of a ruthenium-based cGAS-STING photoactivator for carrier-free cancer immunotherapy, Eur. J. Med. Chem., 2024, 275, 116638 CrossRef CAS PubMed.
- C. Qu, X. Shao, R. Jia, G. Song, D. Shi, H. Wang, J. Wang and H. An, Hypoxia Reversion and STING Pathway Activation through Large Mesoporous Nanozyme for Near-Infrared-II Light Amplified Tumor Polymetallic-Immunotherapy, ACS Nano, 2024, 18, 22153–22171 CrossRef CAS PubMed.
- S. Zhang, D. Song, W. Yu, J. Li, X. Wang, Y. Li, Z. Zhao, Q. Xue, J. Zhao, J. P. Li and Z. Guo, Combining cisplatin and a STING agonist into one molecule for metalloimmunotherapy of cancer, Natl. Sci. Rev., 2024, 11, nwae020 CrossRef CAS PubMed.
- Q. Zhou, D. Dutta, Y. Cao and Z. Ge, Oxidation-Responsive PolyMOF Nanoparticles for Combination Photodynamic-Immunotherapy with Enhanced STING Activation, ACS Nano, 2023, 17, 9374–9387 Search PubMed.
- S. Thota, D. A. Rodrigues, D. C. Crans and E. J. Barreiro, Ru(II) Compounds: Next-Generation Anticancer Metallotherapeutics, J. Med. Chem., 2018, 61, 5805–5821 CrossRef CAS PubMed.
- L. Zeng, P. Gupta, Y. Chen, E. Wang, L. Ji, H. Chao and Z.-S. Chen, The development of anticancer ruthenium(II) complexes: from single molecule compounds to nanomaterials, Chem. Soc. Rev., 2017, 46, 5771–5804 RSC.
- G. Xu, C. Li, C. Chi, L. Wu, Y. Sun, J. Zhao, X.-H. Xia and S. Gou, A supramolecular photosensitizer derived from an Arene-Ru(II) complex self-assembly for NIR activated photodynamic and photothermal therapy, Nat. Commun., 2022, 13, 3064 Search PubMed.
- Y. Yang, Y. Gao, J. Zhao and S. Gou, An electron-accepting half-sandwich iridium(III) complex for the treatment of hypoxic tumors via synergetic chemo- and phototherapy, Inorg. Chem. Front., 2024, 11, 436–450 RSC.
- Y. Huang, M. Lv, B. Guo, G. Hu, Y. Qian, Z. Su, X. Xue and H.-K. Liu, Selectively attacking tumor cells of Ru/Ir–arene complexes based on meclofenamic acid via cyclooxygenase-2 inhibition, Dalton Trans., 2023, 52, 6922–6933 RSC.
- M. Lv, X. Qian, S. Li, J. Gong, Q. Wang, Y. Qian, Z. Su, X. Xue and H.-K. Liu, Unlocking the potential of iridium and ruthenium arene complexes as anti-tumor and anti-metastasis chemotherapeutic agents, J. Inorg. Biochem., 2023, 238, 112057 CrossRef CAS PubMed.
- K. Xiong, C. Qian, Y. Yuan, L. Wei, X. Liao, L. He, T. W. Rees, Y. Chen, J. Wan, L. Ji and H. Chao, Necroptosis Induced by Ruthenium(II) Complexes as Dual Catalytic Inhibitors of Topoisomerase I/II, Angew. Chem., Int. Ed., 2020, 59, 16631–16637 CrossRef CAS PubMed.
- L. Hao, J. Wang, Z.-Y. Pan, Z.-W. Mao and C.-P. Tan, Photodegradation of carbonic anhydrase IX via a binding-enhanced ruthenium-based photosensitizer, Chem. Commun., 2022, 58, 8069–8072 RSC.
- W.-J. Wang, X. Mu, C.-P. Tan, Y.-J. Wang, Y. Zhang, G. Li and Z.-W. Mao, Induction and Monitoring of DNA Phase Separation in Living Cells by a Light-Switching Ruthenium Complex, J. Am. Chem. Soc., 2021, 143, 11370–11381 CrossRef CAS PubMed.
- C. Liang, J. Xie, S. Luo, C. Huang, Q. Zhang, H. Huang and P. Zhang, A highly potent ruthenium(II)-sonosensitizer and sonocatalyst for in vivo sonotherapy, Nat. Commun., 2021, 12, 5001 CrossRef CAS PubMed.
- X. Xu, M. Chen, S. Jiang, Z. Pan and C. Zhao, Endoplasmic Reticulum-Targeting Iridium(III) Nanosonosensitizer Amplifies Immunogenic Cell Death for Boosted Tumor Sono-Immunotherapy, Adv. Funct. Mater., 2024, 26, 2314780 CrossRef.
- L. V. Lutkus, S. S. Rickenbach and T. M. McCormick, Singlet oxygen quantum yields determined by oxygen consumption, J. Photochem. Photobiol., A, 2019, 378, 131–135 CrossRef CAS.
- J. Nam, Proteostasis In The Endoplasmic Reticulum: Road to Cure, Cancers, 2019, 11, 1793 CrossRef PubMed.
- L. Xu, P.-P. Zhang, X.-Q. Fang, Y. Liu, J.-Q. Wang, H.-Z. Zhou, S.-T. Chen and H. Chao, A ruthenium(II) complex containing a p-cresol group induces apoptosis in human cervical carcinoma cells through endoplasmic reticulum stress and reactive oxygen species production, J. Inorg. Biochem., 2019, 191, 126–134 Search PubMed.
- M. R. Gill, D. Cecchin, M. G. Walker, R. S. Mulla, G. Battaglia, C. Smythe and J. A. Thomas, Targeting the endoplasmic reticulum with a membrane-interactive luminescent ruthenium(II) polypyridyl complex, Chem. Sci., 2013, 4, 4512–4519 RSC.
- B. Purushothaman, P. Arumugam, H. Ju, G. Kulsi, A. A. S. Samson and J. M. Song, Novel ruthenium(II) triazine complex [Ru(bdpta)(tpy)]2+ co-targeting drug resistant GRP78 and subcellular organelles in cancer stem cells, Eur. J. Med. Chem., 2018, 156, 747–759 CrossRef CAS PubMed.
- H. G. Xu, M. Schikora, M. Sisa, S. Daum, I. Klemt, C. Janko, C. Alexiou, G. Bila, R. Bilyy, W. Gong, M. Schmitt, L. Sellner and A. Mokhir, An Endoplasmic Reticulum Specific Pro-amplifier of Reactive Oxygen Species in Cancer Cells, Angew. Chem., Int. Ed., 2021, 60, 11158–11162 Search PubMed.
- M. Song, C. Wang, H. Yang, Y. Chen, X. Feng, B. Li and H. Fan, P-STAT, 3 Inhibition Activates Endoplasmic Reticulum Stress-Induced Splenocyte Apoptosis in Chronic Stress, Front. Physiol., 2020, 11, 00680 CrossRef PubMed.
- R. Huang, Q. Ning, J. Zhao, X. Zhao, L. Zeng, Y. Yi and S. Tang, Targeting STING for cancer immunotherapy: From mechanisms to translation, Int. Immunopharmacol., 2022, 113, 109304 CrossRef CAS PubMed.
- A. P. McFarland, S. Luo, F. Ahmed-Qadri, M. Zuck, E. F. Thayer, Y. A. Goo, K. Hybiske, L. Tong and J. J. Woodward, Sensing of Bacterial Cyclic Dinucleotides by the Oxidoreductase RECON Promotes NF-κB Activation and Shapes a Proinflammatory Antibacterial State, Immunity, 2017, 46, 433–445 CrossRef CAS PubMed.
- J. A. Smith, STING, the Endoplasmic Reticulum, and Mitochondria: Is Three a Crowd or a Conversation?, Front. Immunol., 2021, 11, 611347 CrossRef PubMed.
Footnotes |
| † Electronic supplementary information (ESI) available. See DOI: https://doi.org/10.1039/d4qi03021d |
| ‡ These authors contributed equally to this work. |
| This journal is © the Partner Organisations 2025 |