
 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a Creative Commons Attribution 3.0 Unported Licence
Linking iron-deficiency with allergy: role of molecular allergens and the microbiome
Franziska
Roth-Walter
 *a,
Luis F.
Pacios
b,
Rodolfo
Bianchini
a and
Erika
Jensen-Jarolim
ac
*a,
Luis F.
Pacios
b,
Rodolfo
Bianchini
a and
Erika
Jensen-Jarolim
ac
aDepartment of Comparative Medicine, at the Interuniversity Messerli Research Institute, University of Veterinary Medicine Vienna, Medical University of Vienna and University of Vienna, Vienna, Austria. E-mail: Franziska.roth-walter@meduniwien.ac.at
bCentro de Biotecnología y Genómica de Plantas (CBGP, UPM-INIA), Universidad Politécnica de Madrid (UPM) – Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria (INIA), Campus de Montegancedo-UPM, 28223 Pozuelo de Alarcón, Madrid, Spain
cInstitute of Pathophysiology and Allergy Research, Center of Pathophysiology, Infectiology and Immunology, Medical University of Vienna, Vienna, Austria
First published on 9th November 2017
Abstract
Atopic individuals tend to develop a Th2 dominant immune response, resulting in hyperresponsiveness to harmless antigens, termed allergens. In the last decade, epidemiological studies have emerged that connected allergy with a deficient iron-status. Immune activation under iron-deficient conditions results in the expansion of Th2-, but not Th1 cells, can induce class-switching in B-cells and hampers the proper activation of M2, but not M1 macrophages. Moreover, many allergens, in particular with the lipocalin and lipocalin-like folds, seem to be capable of binding iron indirectly via siderophores harboring catechol moieties. The resulting locally restricted iron-deficiency may then lead during immune activation to the generation of Th2-cells and thus prepare for allergic sensitization. Moreover, iron-chelators seem to also influence clinical reactivity: mast cells accumulate iron before degranulation and seem to respond differently depending on the type of the encountered siderophore. Whereas deferoxamine triggers degranulation of connective tissue-type mast cells, catechol-based siderophores reduce activation and degranulation and improve clinical symptoms. Considering the complex interplay of iron, siderophores and immune molecules, it remains to be determined whether iron-deficiencies are the cause or the result of allergy.
 Franziska Roth-Walter | Franziska Roth-Walter studied Chemistry and completed her PhD at the University of Vienna. She did her post-doctoral training in Immunology at the Medical University of Vienna, Austria, and the Mount Sinai School of Medicine, New York, USA. Since 2011 she has been working in the Interuniversity Messerli Research Institute in Vienna. She habilitated for Immunology in 2014 at the Medical University of Vienna. Her research is focused on understanding the mechanisms in mucosal immunity and allergy. |
 Luis F. Pacios | Luis F. Pacios is a physical chemist, obtained his PhD in Quantum Chemistry at the University Complutense of Madrid and completed his training in Clarkson University, New York, USA. Since 1987 he has been Associate Professor in the Technic University of Madrid where he currently teaches Protein Structure and Computational Structural Biology. His research focuses on the computational modeling of molecular structures and interactions in protein–ligand systems. |
 Rodolfo Bianchini | Rodolfo Bianchini is a senior scientist at the Interuniversity Messerli Research Institute in the Department of Comparative Medicine, Vienna, Austria. He obtained his PhD in Clinical and Experimental Pharmacology at the University of Perugia, Italy, and completed his post-doctoral training at the University of Perugia and the University of Salzburg in Austria. His research is primarily focused on modulating macrophage responses in allergy and cancer. |
 Erika Jensen-Jarolim | Erika Jensen-Jarolim, MD, immunologist, holds a professorship at the Medical University of Vienna, affiliated to the Interuniversity Messerli Research Institute where she directs the Department for Comparative Medicine. She is active on multiple boards and councils. She sees allergic patients and applies molecular allergy diagnosis. Her research focuses on immune mechanisms of molecular allergens and the immune regulation in allergen immunotherapy and cancer, in line with the concept of AllergoOncology. |
Introduction
Iron is an essential nutrient utilized in almost every aspect of normal cell function. All cells require iron to proliferate, iron being essential for DNA biosynthesis, protein function and cell cycle progression. In humans, iron is critical for a wide variety of biological processes as it allows transportation of oxygen, aids in the energy household and is essential for a healthy immune system.Iron deficiency is probably the most common cause for anaemia, which by clinical definition is an insufficient mass of circulating red blood cells, and in public health terms is defined as a haemoglobin concentration below the thresholds given by the WHO, UNICEF and UNU.1,2 As such, iron deficiency can exist in the absence of anaemia, if it is mild or if the deficiency lasts not long enough.2
Iron deficiency can be absolute or functional, though combinations also exist. In absolute iron deficiency, the body has to cope with increased iron demand (e.g. during growth, blood donations, bleedings, and infections) that cannot sufficiently be compensated by dietary iron absorption and the release of recycled iron from senescent erythrocytes by macrophages.3 In contrast, during functional iron deficiency, the body has enough iron stores in the form of iron-laden ferritin in the liver, spleen and bone marrow, but iron-trafficking is on hold and/or reversed.
The different settings of iron metabolism are illustrated in Fig. 1. Under normal steady-state conditions, recycled iron is continuously released into the circulation by macrophages and dietary iron by enterocytes via the iron-exporter ferroportin in the form of Fe(II) (Fig. 1A). Subsequently, ceruloplasmin or membrane-bound hephaestin oxidizes iron to Fe(III) for transport via transferrin.4 Functional iron deficiency (Fig. 1B) is the consequence of an activated immune system that is triggered by danger signals derived from pathogens or damaged tissue and which results in a stop of the continuous iron release from enterocytes and macrophages and the accumulation of circulating iron into macrophages.
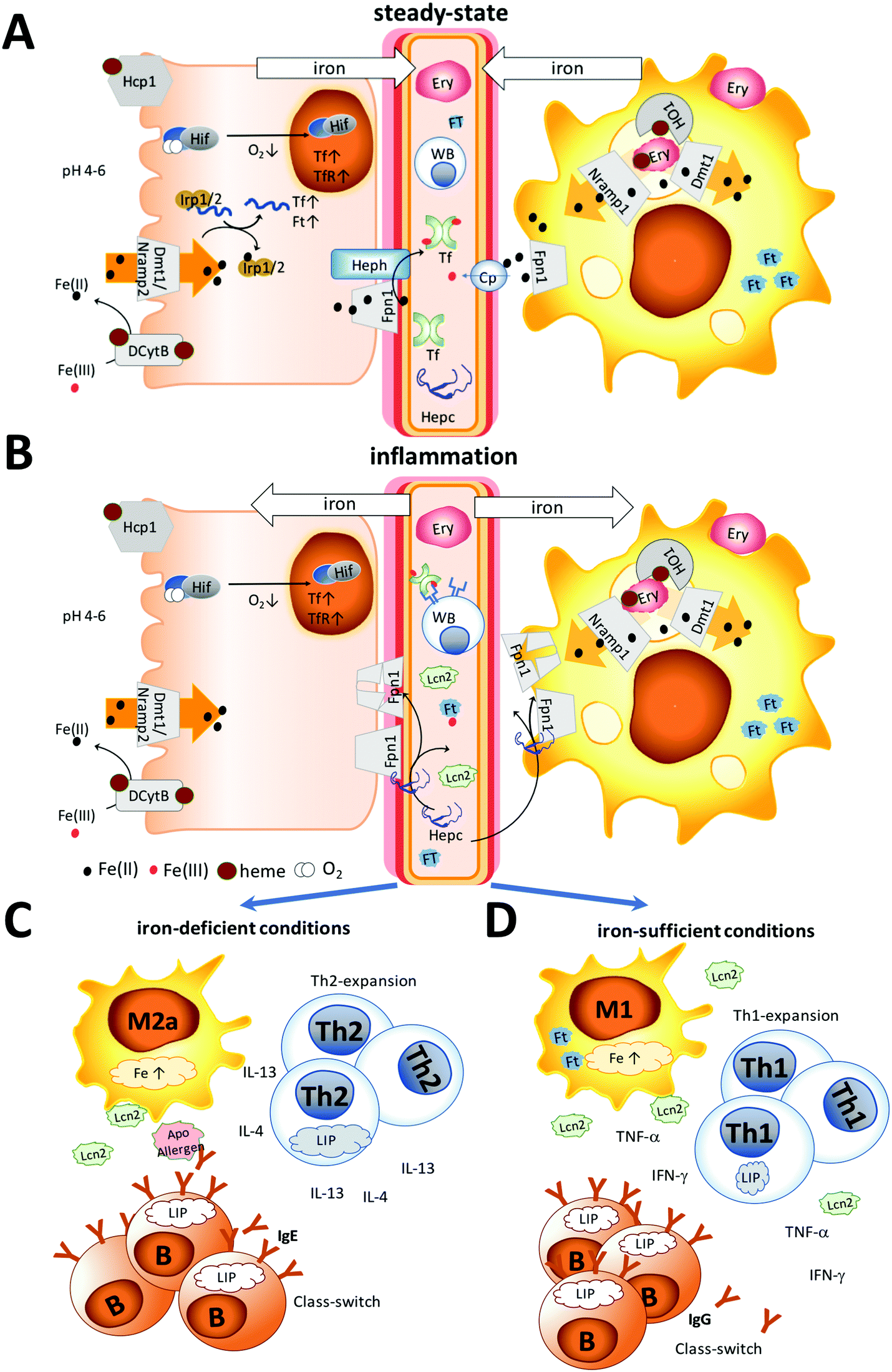 | ||
| Fig. 1 Iron homeostasis under steady-state and inflammation. (A) Under steady-state conditions, dietary iron absorption occurs via (1) the divalent metal-ion transporter 1 (Dmt1) upon reduction of ferric iron in its ferrous form by ascorbic acid or duodenal cytochrome b (Dcytb) and (2) the heme transporter (Hcp1) in the form of heme. Iron is released into the circulation by (3) macrophages recycling senescent erythrocytes with the help of heme oxygenase 1 (HO1) and dietary iron by enterocytes via the iron-exporter ferroportin (Fpn1) in the form of Fe(II). Subsequently, ceruloplasmin (Cp) or membrane-bound hephaestin (Heph) oxidizes iron to Fe(III) for transport via transferrin (Tf). Intracellularly iron concentrations are regulated by the iron regulatory proteins (Irp)1/2. Binding of these proteins to iron results in the transcription of transferrin and ferritin (Ft). Also, a decrease of oxygen can activate hypoxia-inducible factor (Hif), leading to transcription of Tf and the transferrin receptor (TfR). (B) Under inflammation, not only hepcidin (Hepc), but also innate proteins like lipocalin 2 (Lcn2) and ferritin are released into the circulation, Hepc binding to ferroportin (FpN1), thereby initiating its degradation and leading to cytoplasmic accumulation of iron. The immune response differs depending on the human's iron status. (C) Under limited iron supply Th2-, but not Th1-cells, will expand due to their larger intracellular iron pool and secrete IL-4 and IL-13. M2 macrophages will differentiate into an allergic subtype, further providing a Th2 environment. B cells expand via transferrin receptor-independent mechanisms and induce class-switching in the presence of IL-4 and IL-13 towards IgE. Apo-allergens may cause local iron-deprivation or interfere with the regulatory functions of Lcn2 and further modulate the immune-activation. (D) In contrast, under sufficient iron-supply, macrophages may release Lcn2, Th1 cells will expand, secreting IFN-γ and TNF-α, and B cells will generate IgG antibodies. | ||
Discrimination between the two forms is complicated by the existence of various degrees and levels of absolute and functional iron deficiency and their smooth transition into anemia.3 Moreover, many proteins contributing to iron homeostasis are also innate proteins, e.g. ferritin, lactoferrin, and lipocalin 2, which are released into the circulation upon immune activation to impede iron-sequestration by pathogens. This all complicates the assessment of the true iron status.
An interesting aspect of iron deficiency it that a decrease in red blood cells is often accompanied by a relative increase of the white blood cell population representing the immune cells per se.
With respect to allergy, several epidemiological studies have correlated a greater degree of iron deficiency in allergic subjects than in non-allergic individuals.5,6 Whether this is the result of an overboard in the immune response, in which a functional iron-deficiency becomes absolute, or whether an absolute iron-deficiency lays the ground for the generation of allergy remains to be determined.
This review aims to address several important environmental, immunological and physiological aspects that may influence an individual's iron homeostatic status and may contribute to the establishment of allergy.
Humans and iron
Redox chemistry of iron
Under physiological conditions, iron is largely found in the ferrous Fe(II) or ferric Fe(III) form and the metal's redox cycling properties make it highly suited to act as a biocatalyst in proteins or as an electron carrier. In general, Fe(III) prefers oxygen ligands, while Fe(II) favors nitrogen and sulfur ligands.7 In the human body typical examples of Fe(III) binding proteins are lactotransferrin, transferrin and ferritin oxidizing Fe(II) to Fe(III) upon binding, and low molecular weight compounds in the blood also bind Fe(III), with citric acid being the major representative.8 Also, amino acids, ATP/AMP, inositol phosphates and 2,5-dihydroxybenzoic acid have been described to chelate Fe(III) but not Fe(II).9 A different picture emerges in the cytosolic compartment of cells, in which about 1 μM of Fe(II) predominates the labile iron pool, with glutathione in cellular concentrations ranging from 0.5 to 10 mM10,11 acting as a buffer9 and thus serving as a means for the subsequent incorporation of Fe(II) into a wide range of iron-dependent enzymes and electron transfer proteins.9Importantly, free redox-active iron can be very toxic under aerobic conditions due to the Haber–Weiss cycle.12 In this cycle Fe(III) is reduced to Fe(II) by superoxide or other reducing agents and the oxidation of Fe(II) produces Fe(III) and hydroxyl radicals. As such, iron serves as a catalyst and minute amounts of free iron are sufficient to produce significant levels of reactive oxygen species (ROS).
Human iron homeostasis
An 80 kg human body contains approximately 4 g of iron. Hemoglobin iron accounts for approximately 60% of total iron, the vast majority of which is found within circulating erythrocytes.13 Most of the remaining iron is stored in the liver within ferritin (≈1 g). From an immunological point of view, it is interesting to note that the next largest iron-stores are the macrophages (≈0.6 g) in the spleen, liver and bone marrow.14 Around 0.3 g of iron is stored as myoglobin of muscles. All other cellular iron-containing proteins and enzymes are estimated to bind a total of ≈8 mg of iron. Iron is delivered to most tissues via circulating transferrin, which carries ≈2 mg of this metal in the steady state. Iron mass in the total extracellular fluid volume is about 10 mg, implicating that the transferrin iron pool turns over several times a day.15 In healthy men plasma iron turnover ranges from 25 to 35 mg16 per day, of which only 5 to 10% is provided by absorption of dietary iron in the gut, and the rest is predominantly iron recycled from monocytes and macrophages of the liver, adipose tissue, bone marrow, spleen and lymph nodes.14Nutritional iron uptake
Dietary iron requirements, as well as bioavailability, are mainly determined by an individual's iron homeostatic status, affected by physiological conditions and reflected to a large extent in serum hepcidin levels.17 The chief area of iron absorption is the duodenum and the proximal jejunum.18 The duodenum has some unique characteristics as its pH is more acidic, with a pH ranging from 4 to 5 compared to the rest of the small intestine that has a pH-range between 7 and 9, and is the site where pancreatic juices and bile enter the small intestine. Depending on its form, iron will also be transported,(a) as heme (from meat) into the enterocytes via the high-affinity folate transporter, which is also the intestinal heme transporter PCP/HCP1 (SLC46A1).19–21 Interestingly, the duodenal cytochrome b, Dcytb, is also able to bind on the lumen and on the cytoplasmic side to heme molecules,22–25 though the implications of these binding sites have so far been investigated only for the cytoplasmic heme binding site.26
(b) as non-heme iron, typically through low molecular weight chelates of ferric iron, which can derive from meat or plants. After reduction by ascorbic acid and/or duodenal cytochrome b, Dcytb,24,25 iron enters the cells (enterocytes, macrophages, T cells) via the divalent metal-ion transporter 1, DMT1, pathway.26
(c) via other uptake-pathways that seem to exist – e.g. bile itself in “premicellar” concentrations has been shown to interact with iron(II)27 and contribute to iron absorption.28,29 Iron-carrying proteins like ferritin from food are efficiently absorbed without depending on reduction or the heme transporter via receptor-mediated, clathrin-dependent endocytosis.30 Absorption and increased iron accumulation were also found in the liver, when Fe(II) was ingested with glycine and asparagine, but not with other amino acids.31
Once in the cell, iron is exported by ferroportin 1, also known as IREG1, MTP1, SLC40A1, FPN1, and HFE4, into the circulation.32 In general, iron excretion is suppressed by iron deficiency and anaemia and enhanced during erythropoiesis and hypoxia.26
Known inhibitors of bioavailability for non-heme iron are phytates, which are inositol polyphosphates found predominantly in nuts, seeds and grains that form insoluble precipitates with iron,33 and polyphenolic compounds. Many of these fruit- and plant-derived polyphenols bind with high affinity to iron34 and can greatly affect iron homeostasis,35,36 as upon consumption, the plasma concentration of polyphenols can commonly reach values about 1–10 μM.37 In most studies, the consumption of large quantities of purified polyphenols has been reported to decrease the volunteers’ iron status. However, under more natural circumstances, it is reasonable to assume that polyphenols will be present in the food matrix already complexed with iron. As such, one study demonstrated that, while purified polyphenols decreased the iron parameters in the subjects, ingestion of polyphenols in context with iron significantly improved the iron and redox status in vivo.38
Regulation of iron homeostasis
Hepcidin, a 25 amino acid-long peptide, is the major regulator of iron homeostasis. It is mainly expressed in the liver, but can also be produced by parietal cells of the stomach39 and by macrophages.As schematically presented in Fig. 1A, under steady-state conditions only a low concentration of hepcidin is present in circulation, with a median concentration of 7.8 nM found in men and 4.1 nM vs. 8.5 nM, in pre- and post-menopausal women, respectively.40 Dietary and recycled iron is continuously released into the circulation to meet the daily iron requirements of the human body. Excretion of ferrous iron into the circulation is mediated by ferroportin, and subsequently, ceruloplasmin or membrane-bound hephaestin oxidizes Fe(II) to Fe(III) for further transport via transferrin.
During inflammation (Fig. 1B), iron is removed from the circulation, mainly by upregulation of hepcidin, and many innate proteins like Lcn2 and ferritin are also secreted into the circulation to sequester iron. Importantly, also hepcidin-independent processes have been described in humans with iron-deficiency.40 Hepcidin binds to the iron-exporter ferroportin, leading to its degradation, while iron is retained within the cells.40,41 Thus, increasing body iron levels cause increased hepcidin expression, resulting in increased macrophage and liver cell iron sequestration, and decreased dietary iron absorption; the result is a reduction in serum iron.42 In contrast, decreasing body iron levels cause decreased hepcidin expression, resulting in an increased release of macrophage iron and accelerated dietary iron absorption.
Under absolute iron deficiency, a decreased hepcidin concentration will result in the release of the remaining iron stores and in an increase in the dietary iron absorption. Intracellular iron is also regulated by iron regulatory proteins (IRPs) 1 and 2 with iron-responsive elements, in which upregulation of these proteins reflects low body iron stores and an increase of dietary iron absorption.43 Moreover, oxygen can regulate iron homeostasis. During hypoxia, the hypoxia-inducible factors (HIF) can target genes encoding transferrin and the transferrin receptor, leading to increased expression of transferrin and thus increased transport of ferric iron into cells.44
Non-transferrin bound iron and the labile iron pool
Excreted iron is usually bound to transferrin, ferritin or heme. In the circulation, also non-transferrin-bound iron, NTBI, is present, which represents a heterogeneous population that comprises organic anions with low affinity to iron (e.g. citrate, phosphates and carboxylates), polyfunctional ligands (chelates, siderophores and polypeptides), albumin45 and surface components of membranes (e.g. glycans and sulfonates) able to bind Fe(II) and Fe(III). The NTBI is typically present in concentrations up to 10 μM and its existence has been correlated with high levels of transferrin saturation. Similarly, intracellularly, this heterogeneous population of redox-active Fe complexes is called the labile iron pool, LIP. In cells, the iron concentration usually ranges from 20 to 100 μM and is largely associated with proteins, whereas only a minor fraction of the cellular iron is present as the LIP (>1%, up to 5 μM46). In the cytosol, Fe(II) is prevalent, with glutathione probably acting as a major buffer. The LIP is primarily found in erythroid and myeloid cells, as well as in neuronal cells.Importantly, though the NTBI or LIP represents only a fraction of the total intra- and extracellular iron, fine-tuning the LIP-levels has physiologically and immunologically wide-reaching consequences. Both iron forms are the immediate therapeutic targets of diseases associated with a misbalance in the iron homeostasis, e.g. hereditary hemochromatosis, myelodysplastic syndromes and sickle-cell disease.47
The immune system
The immune system and iron
Iron is essential for many peroxide- and nitrous oxide-generating enzymes,48 and regulates cell growth, cell differentiation, and cytokine production. Iron can activate protein kinase C, which leads to phosphorylation of compounds regulating cell proliferation. In addition, iron is necessary for myeloperoxidase activity to form hydroxyl radicals, enabling neutrophils to efficiently eliminate bacteria.49 Thus, any misbalance in the iron homeostasis towards either deficiency or overload has wide-reaching immunological consequences.Macrophages are immune cells known to store iron. The reason for the high presence of iron in macrophages is their pivotal role in systemic iron recycling, where senescent erythrocytes are cleared by predominantly splenic macrophages.50 In addition to their important role in erythrocyte clearance and maintenance, their anti-inflammatory or inflammatory state seems to be dependent on the iron content. Anti-inflammatory macrophages have a lower iron-content and an increased expression of proteins associated with iron efflux and can be discriminated from inflammatory macrophages, which harbour high iron-levels.51 Macrophage phagocytosis is generally unaffected by iron deficiency, even though macrophage bactericidal activity is affected.48 Neutrophils under iron-limited conditions show impaired or lower killing activity due to the reduced activity of myeloperoxidase and reduced mobility to inflamed sites. Likewise, NK-cells exhibit lower activity due to reduced differentiation and proliferation under iron-restricted conditions. Importantly, T lymphocytes can also actively modulate the NTBI pool by uptake and export, with T cell deficiency associated with an accumulation of iron in the liver and pancreas.52 Impaired iron-uptake via transferrin receptor 1 (TfR1) caused by mutations can result in severe B- and T-cell deficiencies due to the lack of activation, which can partly be compensated through the internalization of iron via NTBI-pathways.53
Allergy and iron
Allergy is an immune-mediated disease, caused by an aberrant immune response towards exogenous, normally harmless antigens derived from pollen, house dust mites, animal dander, insect venom and food components. In particular, in the westernized world the prevalence of allergy seems to increase, with almost 20% of the adults in Germany having at least one self-reported doctor diagnosed allergy.54 Also in the United States, the prevalence of respiratory allergies is approximately 20%, whereas the prevalence of food allergies has increased from 3 to 5% and the prevalence of skin allergies from 7 to 12%.55 Allergy defined as type I hypersensitivity causes an immediate reaction usually within minutes upon re-exposure to the antigen in an individual with preformed antigen-specific IgE antibodies. Allergic symptoms may affect the eyes, nose, skin, the lungs or the gastrointestinal tract, leading to red eyes, an itchy rash, runny nose, shortness of breath, swelling or diarrhea. In severe cases, a systemic reaction can occur, resulting in an anaphylactic shock.Mechanistically, it is important to note that the presence of specific IgE antibodies, referred to as allergic sensitization, is a precondition of allergic symptoms. They are a product from plasma cells when stimulated by T-helper 2 cell (Th2-) cytokines IL-4 and IL-13. A Th2 bias in the immune response is typically associated with allergies. Re-exposure to the allergen results in binding and cross-linking of IgE antibodies bound via high-affinity receptors on effector cells. Mast cells then discharge their granules containing histamine, leukotrienes and other active agents by exocytosis, causing allergic reactions.56 However, many people may just be sensitized, thus already having IgE-antibodies, but not yet an allergic reaction upon exposure to the respective allergen.54
As such, the events leading to the initiation of IgE formation are poorly understood, though it is generally accepted that the shift in the immune balance towards Th2 directly correlates with the overproduction of allergen-specific IgE.
Epidemiological and experimental evidence
Along with a lower iron status increasing the risk of atopy, high iron concentration in umbilical cord samples was associated with a decreased risk of wheezing and eczema in the population-based Avon Longitudinal Study of Parents and Children.60 Similarly, in a British study decreased serum ferritin levels were found in children with atopic eczema.61 The lower iron-status is a consistent and a reproducible finding in multiple US cohorts, which clearly associates atopy with anemia.5 Also in a case-controlled, population-based Korean study, low beta-carotene, iron, folic acid, and dietary vitamin E were associated with atopic dermatitis in young children.62 Another study showed a greatly reduced incidence of wheezing and asthma in infants of mothers who were supplemented during pregnancy with vitamin C, a known contributor to increasing iron bioavailability.63 The change in the iron status impacted allergy also in vivo in a murine model of allergic asthma, in which oral iron supplementation, as well as systemic iron administration, suppressed airway manifestations.64
Misregulated iron-metabolism can affect atopy or the generation of allergies. Patients with a much too high iron load due to frequent blood transfusions have a decreased CD4/CD8 ratio,65,66 and their increased serum ferritin levels significantly correlate with the number of transfusions.
More female allergy sufferers: association with iron?
Finally, as iron homeostasis differs between the genders67 and before and after adolescence, it may contribute to the differences seen in the prevalence of allergy in the various groups. During childhood, boys are more affected by allergy than girls, but this changes in adulthood and women are more likely to be affected than men. In a German evaluation, 24% of men, but 35% of women suffer from at least one allergy and 2.9% of men, but 6.4% of women suffer from food allergies.68 Over 20% of Portuguese women were found iron-deficient and other studies have also confirmed that females present more often iron deficiency.69,70All in all, there is consistent evidence that the iron-status of allergic subjects is reduced and may be linked with the disease.
Therefore, iron levels affect in opposite ways the immune regulation in allergy and cancer.
Immune effects of iron
Thus, it is very likely that under conditions of lymphocyte expansion the limited iron supply in allergic subjects will favour the development of a Th2-environment, thereby preparing for later allergic sensitisation.
Corna et al. demonstrated that under iron deprivation by deferoxamine both M1 and M2 macrophages enhanced IRP1 activity, whereas IRP2 was more strongly enhanced in M2 macrophages.82 Deferoxamine treatment also does not lead to the suppression of ferritin heavy chain expression in M1 macrophages, indicating high enough iron stores, whereas in M2 macrophages TfR1 was upregulated for continuous iron supply.51 As such, M1 macrophages are not as affected by iron-deficient conditions as M2 macrophages. Importantly, M2 cells are not as efficient in expressing molecules involved in antigen presentation, such as MHC class II (I-Ab), or the costimulatory molecule CD86 after T-cell stimulation under iron-deficient conditions, whereas M1 macrophages seem unaffected by iron chelators.
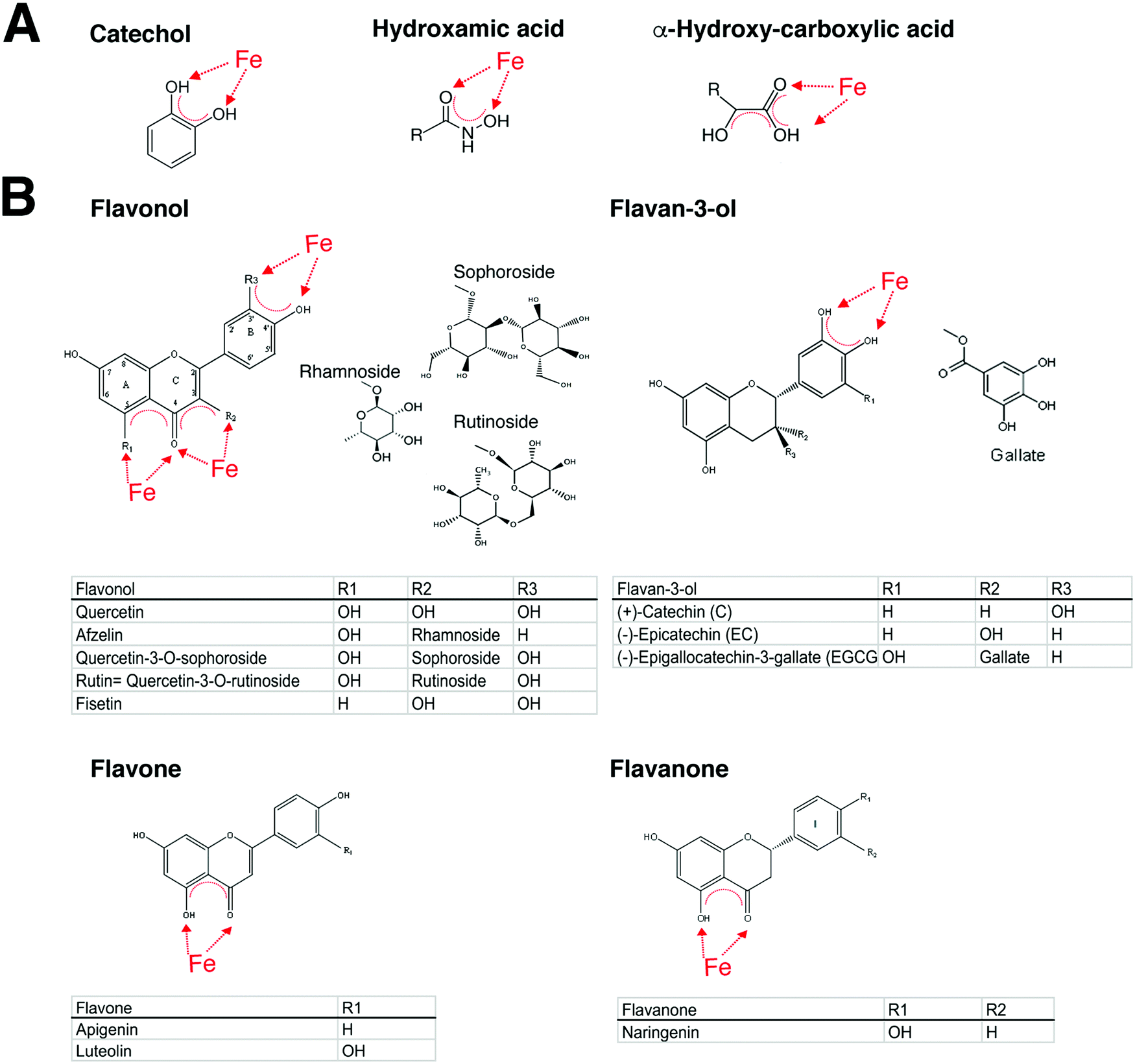 | ||
| Fig. 2 Structures with affinity to iron. (A) Siderophores can be classified by their ligands interacting with iron as catecholates, hydroxamates, and hydroxyl-carboxylates. (B) The four different subclasses of flavonoids namely flavonols, flavan-3-ols, flavones and flavanones, with described ligand-binding activity to allergens or described as having anti-allergic properties. | ||
Allergens
Nearly all major allergens from mammals belong to the lipocalin family,102 while plant allergens usually originate from the prolamin (2S albumin, lipid binding proteins (LTPs)) and cupin (7S, 11S) superfamilies or from the pathogenesis-related (PR)-10 family.103 All members of these families share certain characteristics like their great structural stability and their ability to serve as carriers for a variety of compounds with lipidic segments.104,105
Allergens deriving from mammalian sources usually belong to the lipocalin family. Lipocalins show unusually low levels of overall sequence conservation with pairwise sequence identities often below 20%. Nevertheless, as illustrated in Fig. 3 the lipocalin fold is highly conserved.106 This β-barrel structure shapes a calyx-like site which gives the name to the protein family and is the main feature regarding the binding abilities of the lipocalin fold.107 While the wider end of the barrel is open to the solvent and rich in polar and charged amino acids, the narrower end is an inner, buried region rich in hydrophobic amino acids. Loops flanking the calyx display a great sequence variability that endows lipocalins with the ability to bind a large variety of ligands having polar and non-polar moieties. This property has been exploited in protein design that uses lipocalins as scaffolds to engineer novel binding proteins (“anticalins”).108
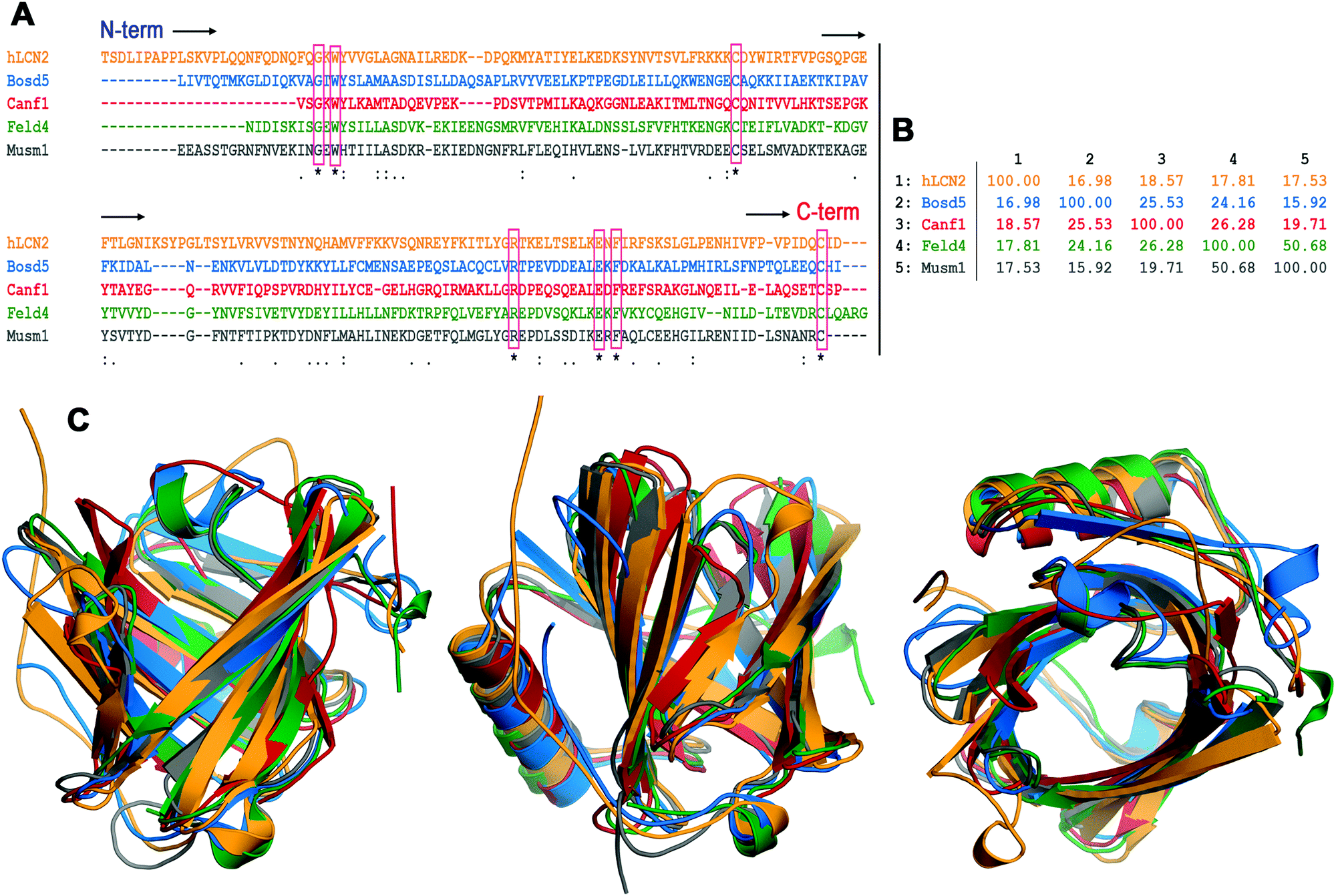 | ||
| Fig. 3 Sequence and structural alignment of lipocalins. (A) Clustal multiple sequence alignment of the following lipocalins. hLcn2: human Lcn2, Bos d 5: bovine β-lactoglobulin, Can f 1: dog allergen, Fel d 4: cat allergen, and Mus m 1: pheromone binding rodent major urinary protein from a mouse. The seven residues strictly conserved are boxed. (B) Percent identity matrix for the multiple sequence alignment in A. (C) Three views of the structural superposition of the hLcn2 (orange) crystal structure PDB id 1l6 m, Bos d 5 (blue) crystal structure PDB id 3NPO, Can f 1 (red) and Fel d 4 (green) homology model structures (ref. 100), and Mus m 1 (gray) crystal structure PDB id 1MUP. The percentages of allergen residues and RMSD values for backbone atoms in the structural superposition with hLcn2 are the following – Bos d 5: 67%, 1.362 Å; Can f 1: 66%, 1.574 Å; Fel d 4: 68%, 1.235 Å; Mus m 1: 73%, 1.158 Å. | ||
Lipocalins are usually secreted and can be found in the dander, urine, fur, and saliva of animals.109 They have been described as powerful bacteriostatic agents against various microorganisms, by impeding their iron sequestration and binding to siderophore–iron complexes,110,111 but in accordance with their ligand binding plasticity, lipocalins also seem to act as carriers for lipids and hormones.
The human homologue lipocalin 2 (Lcn2, NGAL) has immune-regulatory function, but is also a growth factor112 and a stress protein that is released under various inflammatory conditions and in cancer.113 The binding of Lcn2 to bacterial siderophores, which are low molecular weight compounds that are amongst the strongest soluble Fe(III) binding agents known, is considered a key feature against bacterial infections. Secreted Lcn2 seems to have specialized towards siderophores with catechol-moieties that facilitate their binding in the calyx site (see Fig. 4).114 Moreover, Lcn2 has been described to bind to endogenous siderophores like 2,5-dihydroxybenzoic acid (2,5-DHBA),115 thereby probably ensuring that excess free iron does not accumulate in the cytoplasm. Mammalian cells lacking this endogenous siderophore accumulate abnormally high levels of intracellular iron, leading to high levels of reactive oxygen species.115
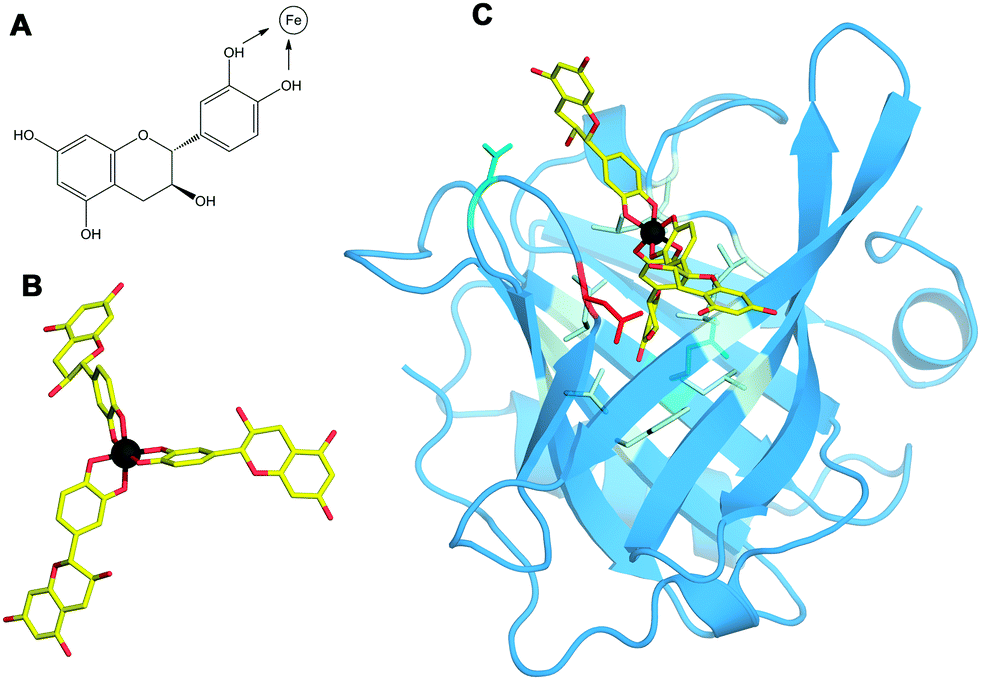 | ||
| Fig. 4 Structure of the Bos d 5–Fe(catechin)3 complex. (A) The structural formula of catechin. Catechol hydroxyls participating in the chelation of iron are marked with arrows. (B) 3D structure of the Fe(catechin)3 ligand showing the spatial arrangement of catechols to set the hexadentate octahedral coordination of iron (carbons in yellow, oxygen in red, and iron in dark gray). (C) 3D structure of the Bos d 5–Fe(catechin)3 complex showing the location of the ligand at the calyx of the lipocalin. Residues within a 3.5 Å distance from the ligand are displayed as sticks with the following color code: nine non-polar in pale cyan, two polar in cyan and one acidic in red. | ||
Importantly, also many fruits and plants feature the presence of polyphenols/flavonoids known to behave as high-affinity iron chelators, a fact that is commonly overlooked,34 but that may contribute to their mast cell stabilizing ability. In addition, the ability of those compounds to chelate iron has been proposed to be a key element in their anti-oxidant properties.118
With regard to allergy, there seems to exist a particular role for catechol-type siderophores, which may also be related to the fact that catechol groups chelate iron119,120 at physiologically relevant pHs.
In this respect, it is relevant that lipocalin 2 can bind to only catechol-containing siderophores and not to others. This is an important characteristic that also seems to be extendable to many major allergens. Many lipocalin allergens such as the major milk allergen Bos d 5, or the major birch allergen Bet v 1, a prototype for the PR-10 protein family with a lipocalin-like architecture, are capable of transporting iron via catechol-type siderophores.121,122 Importantly, their loading state, apo- (empty) or holo- (filled), seemed to be decisive for the subsequent immune response. Apo-allergens can mount a Th2-response in vitro, whereas the holo forms are rather immune-suppressive, indicating that the iron-carrying form impedes allergic sensitization.121,122
As such, the natural ligand of the major birch pollen allergen Bet v 1 has been identified to be the flavonoid quercetin-3-O-sophoroside;123 the major peanut allergens Ara h 2 and Ara h 6, belonging to the 2S family, bind the flavonoid epigallocatechin-3-gallate;124 the major peanut allergen Ara h 1 from the 7S family forms large complexes by binding proanthocyanidins, which are oligomers consisting of catechin and epicatechin and their gallic acid esters.125 Accordingly, the pathogenesis-related PR-10 proteins and major allergens in strawberries, Fra a 1 and Fra a 3, have been crystallized with catechin ligands.126
In summary, there is solid evidence that allergens are capable of binding in particular catechol-type structures like quercetin and rutin with high iron affinities that surpass iron-affinity constants of deferoxamine.127,128
Importantly, a great number of these polyphenols have been described in the literature to exhibit anti-allergic properties. Taking lipocalins as an example, allergens are not only structurally at the border between self and non-self, but may also functionally interfere with the human immune-regulatory processes.
How normally harmless antigens become allergens with their characteristic Th2 skewing capacity is not known. There are several possible scenarios: (i) iron deficiency turns holo-lipocalins into apo-allergens, or (ii) exogenous lipocalins enter the body already devoid of a ligand in their calyx. In this case, they – due to their high affinity – will immediately sequester endogenous siderophores or comparable ligands directly at the mucosal sites, thereby contributing locally to an iron-deficient state. Additional triggers then may activate lymphocytes to become Th2 rather than Th1 cells.121,122,128 (iii) Allergenic lipocalins may in an ongoing immune response interfere with the function of Lcn2, e.g. during infections either by providing or by sequestering ligands for Lcn2. In any of these cases, the immune function of Lcn2 may be skewed by allergens towards Th2.
Can microbiota interfere with iron homeostasis?
Microbiome depletion as a result of broad-spectrum antibiotic treatment led not only to anaemia and thrombocytosis but also to an altered immune homeostasis, resulting in diminished granulocytes and B cells116 in the bone marrow and increased CD8+ T-cells in mice. Importantly, faecal microbiota transfer partly rescued these hematopoietic changes.134 In rats inoculated with human faecal microbiota, provision of an iron-fortified diet after feeding an iron-deficient diet significantly increased the abundance of dominant bacterial groups such as Bacteroides spp. and Clostridium cluster IV members compared with rats on an iron-deficient diet only. Moreover, iron supplementation increased the gut microbial butyrate concentration 6-fold compared with iron depletion and did not affect histological colitis scores, suggesting that iron supplementation enhanced the concentration of beneficial gut microbiota metabolites and may contribute to gut health.135
Importantly, microbes able to inhabit the upper G/I tract seemed to be reduced in allergic subjects compared to non-allergic subjects.
There are only a few studies on the microbiota conducted in humans. As such, food allergic patients seem to have an increased abundance of bacteria of the order Clostridiales (Lachnospiraceae, Ruminococcaceae)136–139 and a decreased abundance of the order Bacteroidales.136,140,141
| Phylum | Family | Genus | Siderophore use | Reported abundancy in allergy |
|---|---|---|---|---|
| Code: predominantly found in colon (bold face), upper G/I (regular font). Ferritin – fr, transferrin – tr, lactoferrin – lr, hemophores – hr. | ||||
| Actinobacteria137 | Bifidobacteriaceae 136,140,141 | Bifidobacterium 140 | +/−, Fe(II) | Increased 140 /decreased 136–138,141 |
| Coriobacteriaceae 140 | Collinsella 140 | + | Increased 140/decreased137,141 | |
| Bacteriodetes137,140,142 | Bacteroidaceae | Bacteroides | +/hr | Decreased137,140,141/increased143 |
| Rikenellaceae | Rickenella | +/heme | Decreased141 | |
| Porphyromonadaceae | Porphyromonas | Fe(II)/heme | ||
| Parabacteroides 140 | ? | Decreased140,141 | ||
| Prevotellaceae | Prevotella | +, heme, tr − | Decreased140,141,143/increased144 | |
| Firmicutes | Lactobacillaceae | Lactobacillus 140,141 | −/Mn | Increased140/decreased141 |
| Streptococcaceae | Streptococcus 140 | +, fr, lr, heme, hr | Decreased138,140,141 | |
| Enterococcaceae | Enterococcus | +, heme | Decreased138/increased140,141 | |
| Staphylococcaceae | Staphylocoocus | + | Decreased138,145 | |
| Clostridiaceae/cluster I140,141 | Clostridium | +, heme | Increased136,137,141 | |
| Clostridiaceae/cluster IV 140,141 | Butyricicoccus | ? | n/a | |
| Anaerobacter 140 | ? | Increased 137,140 /decreased 139 | ||
| Lachnospiraceae 138,140,143,146,147 | Lactonifactor | ? | n/a | |
| Blautia | + | Decreased 143 /increased 138 | ||
| Roseburia | + | Increased 138,140,141 | ||
| Ruminococcaceae/cluster IV 140,143 | Ruminococcus | + | Increased 138,140,141 | |
| Faecalibacterium | + | Increased 138,140 /decreased 139,141,143 | ||
| Subdoligranulum | Fe(II) | Increased 138,140 /decreased 141,143 | ||
| Veillonellaceae | Dialister 141 | + | Decreased141 | |
| Megamonas | + | n/a | ||
| Megasphaera 141 | + | Decreased141 | ||
| Veillonella 140 | + | Decreased140,141 | ||
| Proteobacteria137 | Enterobacteriaceae139 | Escherichia | + | Decreased138/increased137,139 |
| Citrobacter | + | Increased139 | ||
| Pseudomonadaceae | Pseudomonas | + | ||
| Moraxellaceae | Acinetobacter 144 | +148 | Decreased144 | |
| Campylobacteraceae | Campylobacter 149 | + | Decreased150 | |
| Arcobacter | + | |||
| Verrucomicrobia | Akkermansiaceae140 | Akkermansia | + | Increased140/decreased139,141 |
| Kingdom Fungi: Ascomycota | Saccharomycetaceae | Candida | + | Increased141 |
The microbiota strongly manipulates the immune system. It is tempting to speculate that the composition and localization of the commensal microbiota in allergic subjects may directly impact the homeostatic iron status of the host, but more studies need to be done.
Conclusions
There is a clear epidemiological connection between a poor iron status and allergy risk, especially in females. Of note, iron-deficient conditions seem to promote a Th2-environment, which is a prerequisite for allergy. Potential contributing factors are endogenous iron levels, allergens capable of binding to iron chelators, and likely a skewed microbiota in allergic subjects.Conflicts of interest
The authors declare no conflicts of interest.Acknowledgements
This work was supported by the Austrian Science Fund FWF grant SFB F4606-B28 to EJJ.References
- United Nations Children's Fund, United Nations University, World Health Organization, Iron Deficiency Anaemia Assessment, Prevention, and Control: A guide for programme managers, WHO/NHD/01.3, 2001, p. 114.
- World Health Organization, Centers for Disease Control and Prevention Technical Consultation. Assessing the iron status of populations, WD 105, 2007, p. 108.
- M. Nairz, I. Theurl, D. Wolf and G. Weiss, Iron deficiency or anemia of inflammation?: Differential diagnosis and mechanisms of anemia of inflammation, Wien. Med. Wochenschr., 2016, 166, 411–423 CrossRef PubMed.
- G. Papanikolaou and K. Pantopoulos, Systemic iron homeostasis and erythropoiesis, IUBMB Life, 2017, 69, 399–413 CrossRef CAS PubMed.
- K. E. Drury, M. Schaeffer and J. I. Silverberg, Association Between Atopic Disease and Anemia in US Children, JAMA Pediatr., 2016, 170, 29–34 CrossRef PubMed.
- T. Haahtela, G. J. Burbach, C. Bachert, C. Bindslev-Jensen, S. Bonini, J. Bousquet, L. Bousquet-Rouanet, P. J. Bousquet, M. Bresciani, A. Bruno, G. W. Canonica, U. Darsow, P. Demoly, S. R. Durham, W. J. Fokkens, S. Giavi, M. Gjomarkaj, C. Gramiccioni, M. L. Kowalski, G. Losonczy, M. Orosz, N. G. Papadopoulos, G. Stingl, A. Todo-Bom, E. von Mutius, A. Kohli, S. Wohrl, S. Jarvenpaa, H. Kautiainen, L. Petman, O. Selroos, T. Zuberbier and L. M. Heinzerling, Clinical relevance is associated with allergen-specific wheal size in skin prick testing, Clin. Exp. Allergy, 2014, 44, 407–416 CrossRef CAS PubMed.
- J. R. Chipperfield and C. Ratledge, Salicylic acid is not a bacterial siderophore: a theoretical study, Biometals, 2000, 13, 165–168 CrossRef CAS PubMed.
- F. I. Adam, P. L. Bounds, R. Kissner and W. H. Koppenol, Redox properties and activity of iron–citrate complexes: evidence for redox cycling, Chem. Res. Toxicol., 2015, 28, 604–614 CrossRef CAS PubMed.
- R. C. Hider and X. L. Kong, Glutathione: a key component of the cytoplasmic labile iron pool, Biometals, 2011, 24, 1179–1187 CrossRef CAS PubMed.
- F. Roth-Walter, P. Starkl, T. Zuberbier, K. Hummel, K. Nobauer, E. Razzazi-Fazeli, R. Brunner, I. Pali-Scholl, J. Kinkel, F. Felix, E. Jensen-Jarolim and T. Kinaciyan, Glutathione exposes sequential IgE-epitopes in ovomucoid relevant in persistent egg allergy, Mol. Nutr. Food Res., 2013, 57, 536–544 CAS.
- A. Meister, Glutathione metabolism and its selective modification, J. Biol. Chem., 1988, 263, 17205–17208 CAS.
- G. A. Kortman, M. Raffatellu, D. W. Swinkels and H. Tjalsma, Nutritional iron turned inside out: intestinal stress from a gut microbial perspective, FEMS Microbiol. Rev., 2014, 38, 1202–1234 CrossRef CAS PubMed.
- G. Pishchany and E. P. Skaar, Taste for blood: hemoglobin as a nutrient source for pathogens, PLoS Pathog., 2012, 8, e1002535 CAS.
- K. Pantopoulos, S. K. Porwal, A. Tartakoff and L. Devireddy, Mechanisms of mammalian iron homeostasis, Biochemistry, 2012, 51, 5705–5724 CrossRef CAS PubMed.
- T. Ganz, Macrophages and systemic iron homeostasis, J. Innate Immun., 2012, 4, 446–453 CrossRef CAS PubMed.
- D. D. Funk, Plasma iron turnover in normal subjects, J. Nucl. Med., 1970, 11, 107–111 CAS.
- D. Demeyer, S. De Smet and M. Ulens, The near equivalence of haem and non-haem iron bioavailability and the need for reconsidering dietary iron recommendations, Eur. J. Clin. Nutr., 2014, 68, 750–751 CrossRef CAS PubMed.
- H. Huebers, E. Huebers, W. Forth and W. Rummel, Binding of iron to a non-ferritin protein in the mucosal cells of normal and iron-deficient rats during absorption, Life Sci., 1971, 10, 1141–1148 CrossRef CAS.
- Y. Nakai, K. Inoue, N. Abe, M. Hatakeyama, K. Y. Ohta, M. Otagiri, Y. Hayashi and H. Yuasa, Functional characterization of human proton-coupled folate transporter/heme carrier protein 1 heterologously expressed in mammalian cells as a folate transporter, J. Pharmacol. Exp. Ther., 2007, 322, 469–476 CrossRef CAS PubMed.
- G. O. Latunde-Dada, K. Takeuchi, R. J. Simpson and A. T. McKie, Haem carrier protein 1 (HCP1): expression and functional studies in cultured cells, FEBS Lett., 2006, 580, 6865–6870 CrossRef CAS PubMed.
- S. Le Blanc, M. D. Garrick and M. Arredondo, Heme carrier protein 1 transports heme and is involved in heme-Fe metabolism, Am. J. Physiol.: Cell Physiol., 2012, 302, C1780–C1785 CrossRef CAS PubMed.
- G. F. da Silva, V. P. Shinkarev, Y. A. Kamensky and G. Palmer, Spectroscopic evidence of the role of an axial ligand histidinate in the mechanism of adrenal cytochrome b561, Biochemistry, 2012, 51, 8730–8742 CrossRef CAS PubMed.
- A. T. McKie, D. Barrow, G. O. Latunde-Dada, A. Rolfs, G. Sager, E. Mudaly, M. Mudaly, C. Richardson, D. Barlow, A. Bomford, T. J. Peters, K. B. Raja, S. Shirali, M. A. Hediger, F. Farzaneh and R. J. Simpson, An iron-regulated ferric reductase associated with the absorption of dietary iron, Science, 2001, 291, 1755–1759 CrossRef CAS PubMed.
- J. S. Oakhill, S. J. Marritt, E. G. Gareta, R. Cammack and A. T. McKie, Functional characterization of human duodenal cytochrome b (Cybrd1): redox properties in relation to iron and ascorbate metabolism, Biochim. Biophys. Acta, 2008, 1777, 260–268 CrossRef CAS PubMed.
- S. Ludwiczek, F. I. Rosell, M. L. Ludwiczek and A. G. Mauk, Recombinant expression and initial characterization of the putative human enteric ferric reductase Dcytb, Biochemistry, 2008, 47, 753–761 CrossRef CAS PubMed.
- D. J. Lane, D. H. Bae, A. M. Merlot, S. Sahni and D. R. Richardson, Duodenal cytochrome b (DCYTB) in iron metabolism: an update on function and regulation, Nutrients, 2015, 7, 2274–2296 CrossRef CAS PubMed.
- A. Fini, G. Feroci, G. Fazio and P. Zuman, Interaction of iron(II) with bile salts, J. Inorg. Biochem., 1997, 68, 251–256 CrossRef CAS PubMed.
- A. J. Sanyal, M. L. Shiffmann, J. I. Hirsch and E. W. Moore, Premicellar taurocholate enhances ferrous iron uptake from all regions of rat small intestine, Gastroenterology, 1991, 101, 382–389 CrossRef CAS.
- A. J. Sanyal, J. I. Hirsch and E. W. Moore, Evidence that bile salts are important for iron absorption, Am. J. Physiol., 1994, 266, G318–G323 CAS.
- E. C. Theil, H. Chen, C. Miranda, H. Janser, B. Elsenhans, M. T. Nunez, F. Pizarro and K. Schumann, Absorption of iron from ferritin is independent of heme iron and ferrous salts in women and rat intestinal segments, J. Nutr., 2012, 142, 478–483 CrossRef CAS PubMed.
- J. M. Christensen, M. Ghannam and J. W. Ayres, Effects of divalent amino acids on iron absorption, J. Pharm. Sci., 1984, 73, 1245–1248 CrossRef CAS PubMed.
- A. Donovan, A. Brownlie, Y. Zhou, J. Shepard, S. J. Pratt, J. Moynihan, B. H. Paw, A. Drejer, B. Barut, A. Zapata, T. C. Law, C. Brugnara, S. E. Lux, G. S. Pinkus, J. L. Pinkus, P. D. Kingsley, J. Palis, M. D. Fleming, N. C. Andrews and L. I. Zon, Positional cloning of zebrafish ferroportin1 identifies a conserved vertebrate iron exporter, Nature, 2000, 403, 776–781 CrossRef CAS PubMed.
- S. Clemens, Zn and Fe biofortification: the right chemical environment for human bioavailability, Plant Sci., 2014, 225, 52–57 CrossRef CAS PubMed.
- M. Dell'mour, W. Schenkeveld, E. Oburger, L. Fischer, S. Kraemer, M. Puschenreiter, M. Lammerhofer, G. Koellensperger and S. Hann, Analysis of iron-phytosiderophore complexes in soil related samples: LC-ESI-MS/MS versus CE-MS, Electrophoresis, 2012, 33, 726–733 CrossRef PubMed.
- J. R. Hunt and Z. K. Roughead, Nonheme-iron absorption, fecal ferritin excretion, and blood indexes of iron status in women consuming controlled lactoovovegetarian diets for 8 wk, Am. J. Clin. Nutr., 1999, 69, 944–952 CAS.
- J. Suliburska, P. Bogdanski, M. Szulinska, M. Stepien, D. Pupek-Musialik and A. Jablecka, Effects of green tea supplementation on elements, total antioxidants, lipids, and glucose values in the serum of obese patients, Biol. Trace Elem. Res., 2012, 149, 315–322 CrossRef CAS PubMed.
- N. R. Perron and J. L. Brumaghim, A review of the antioxidant mechanisms of polyphenol compounds related to iron binding, Cell Biochem. Biophys., 2009, 53, 75–100 CrossRef CAS PubMed.
- A. Imessaoudene, H. Merzouk, F. Berroukeche, N. Mokhtari, B. Bensenane, S. Cherrak, S. A. Merzouk and M. Elhabiri, Beneficial effects of quercetin-iron complexes on serum and tissue lipids and redox status in obese rats, J. Nutr. Biochem., 2016, 29, 107–115 CrossRef CAS PubMed.
- P. Schwarz, J. A. Kubler, P. Strnad, K. Muller, T. F. Barth, A. Gerloff, P. Feick, C. Peyssonnaux, S. Vaulont, G. Adler and H. Kulaksiz, Hepcidin is localised in gastric parietal cells, regulates acid secretion and is induced by Helicobacter pylori infection, Gut, 2012, 61, 193–201 CrossRef CAS PubMed.
- T. E. Galesloot, S. H. Vermeulen, A. J. Geurts-Moespot, S. M. Klaver, J. J. Kroot, D. van Tienoven, J. F. Wetzels, L. A. Kiemeney, F. C. Sweep, M. den Heijer and D. W. Swinkels, Serum hepcidin: reference ranges and biochemical correlates in the general population, Blood, 2011, 117, e218–e225 CrossRef CAS PubMed.
- G. Ramey, J. C. Deschemin, B. Durel, F. Canonne-Hergaux, G. Nicolas and S. Vaulont, Hepcidin targets ferroportin for degradation in hepatocytes, Haematologica, 2010, 95, 501–504 CrossRef CAS PubMed.
- T. Nakanishi, Y. Hasuike, M. Nanami, M. Yahiro and T. Kuragano, Novel iron-containing phosphate binders and anemia treatment in CKD: oral iron intake revisited, Nephrol., Dial., Transplant., 2016, 31, 1588–1594 CrossRef PubMed.
- M. Munoz, J. A. Garcia-Erce and A. F. Remacha, Disorders of iron metabolism. Part 1: molecular basis of iron homoeostasis, J. Clin. Pathol., 2011, 64, 281–286 CrossRef CAS PubMed.
- A. Rolfs, I. Kvietikova, M. Gassmann and R. H. Wenger, Oxygen-regulated transferrin expression is mediated by hypoxia-inducible factor-1, J. Biol. Chem., 1997, 272, 20055–20062 CrossRef CAS PubMed.
- A. M. Silva and R. C. Hider, Influence of non-enzymatic post-translation modifications on the ability of human serum albumin to bind iron. Implications for non-transferrin-bound iron speciation, Biochim. Biophys. Acta, 2009, 1794, 1449–1458 CrossRef CAS PubMed.
- Y. Ma, H. de Groot, Z. Liu, R. C. Hider and F. Petrat, Chelation and determination of labile iron in primary hepatocytes by pyridinone fluorescent probes, Biochem. J., 2006, 395, 49–55 CrossRef CAS PubMed.
- L. de Swart, J. C. Hendriks, L. N. van der Vorm, Z. I. Cabantchik, P. J. Evans, E. A. Hod, G. M. Brittenham, Y. Furman, B. Wojczyk, M. C. Janssen, J. B. Porter, V. E. Mattijssen, B. J. Biemond, M. A. MacKenzie, R. Origa, R. Galanello, R. C. Hider and D. W. Swinkels, Second international round robin for the quantification of serum non-transferrin-bound iron and labile plasma iron in patients with iron-overload disorders, Haematologica, 2016, 101, 38–45 CrossRef CAS PubMed.
- J. L. Beard, Iron biology in immune function, muscle metabolism and neuronal functioning, J. Nutr., 2001, 131, 568S–579S CAS , discussion 580S.
- E. S. Wintergerst, S. Maggini and D. H. Hornig, Contribution of selected vitamins and trace elements to immune function, Ann. Nutr. Metab., 2007, 51, 301–323 CrossRef CAS PubMed.
- D. Z. de Back, E. B. Kostova, M. van Kraaij, T. K. van den Berg and R. van Bruggen, Of macrophages and red blood cells; a complex love story, Front. Physiol., 2014, 5, 9 Search PubMed.
- S. Recalcati, M. Locati, A. Marini, P. Santambrogio, F. Zaninotto, M. De Pizzol, L. Zammataro, D. Girelli and G. Cairo, Differential regulation of iron homeostasis during human macrophage polarized activation, Eur. J. Immunol., 2010, 40, 824–835 CrossRef CAS PubMed.
- J. P. Pinto, J. Arezes, V. Dias, S. Oliveira, I. Vieira, M. Costa, M. Vos, A. Carlsson, Y. Rikers, M. Rangel and G. Porto, Physiological implications of NTBI uptake by T lymphocytes, Front. Pharmacol., 2014, 5, 24 Search PubMed.
- H. H. Jabara, S. E. Boyden, J. Chou, N. Ramesh, M. J. Massaad, H. Benson, W. Bainter, D. Fraulino, F. Rahimov, C. Sieff, Z. J. Liu, S. H. Alshemmari, B. K. Al-Ramadi, H. Al-Dhekri, R. Arnaout, M. Abu-Shukair, A. Vatsayan, E. Silver, S. Ahuja, E. G. Davies, M. Sola-Visner, T. K. Ohsumi, N. C. Andrews, L. D. Notarangelo, M. D. Fleming, W. Al-Herz, L. M. Kunkel and R. S. Geha, A missense mutation in TFRC, encoding transferrin receptor 1, causes combined immunodeficiency, Nat. Genet., 2016, 48, 74–78 CrossRef CAS PubMed.
- K. C. Bergmann, J. Heinrich and H. Niemann, Current status of allergy prevalence in Germany: position paper of the Environmental Medicine Commission of the Robert Koch Institute, Allergol. J. Int., 2016, 25, 6–10 CrossRef PubMed.
- K. D. Jackson, L. D. Howie and L. J. Akinbami, Trends in allergic conditions among children: United States, 1997–2011, NCHS Data Brief, 2013, 1–8 Search PubMed.
- I. Pali-Scholl, J. Namazy and E. Jensen-Jarolim, Allergic diseases and asthma in pregnancy, a secondary publication, World Allergy Organ. J., 2017, 10, 10 CrossRef PubMed.
- R. Weigert, N. C. Dosch, M. E. Bacsik-Campbell, T. W. Guilbert, C. L. Coe and P. J. Kling, Maternal pregnancy weight gain and cord blood iron status are associated with eosinophilia in infancy, J. Perinatol., 2015, 35, 621–626 CrossRef CAS PubMed.
- M. K. Georgieff, R. L. Schmidt, M. M. Mills, W. J. Radmer and J. A. Widness, Fetal iron and cytochrome c status after intrauterine hypoxemia and erythropoietin administration, Am. J. Physiol., 1992, 262, R485–R491 CAS.
- B. I. Nwaru, H. Hayes, L. Gambling, L. C. Craig, K. Allan, N. Prabhu, S. W. Turner, G. McNeill, M. Erkkola, A. Seaton, H. J. McArdle and G. Devereux, An exploratory study of the associations between maternal iron status in pregnancy and childhood wheeze and atopy, Br. J. Nutr., 2014, 112, 2018–2027 CrossRef CAS PubMed.
- S. O. Shaheen, R. B. Newson, A. J. Henderson, P. M. Emmett, A. Sherriff, M. Cooke and A. S. Team, Umbilical cord trace elements and minerals and risk of early childhood wheezing and eczema, Eur. Respir. J., 2004, 24, 292–297 CrossRef CAS PubMed.
- T. J. David, F. E. Wells, T. C. Sharpe, A. C. Gibbs and J. Devlin, Serum levels of trace metals in children with atopic eczema, Br. J. Dermatol., 1990, 122, 485–489 CrossRef CAS PubMed.
- S. Y. Oh, J. Chung, M. K. Kim, S. O. Kwon and B. H. Cho, Antioxidant nutrient intakes and corresponding biomarkers associated with the risk of atopic dermatitis in young children, Eur. J. Clin. Nutr., 2010, 64, 245–252 CrossRef CAS PubMed.
- C. T. McEvoy, D. Schilling, N. Clay, K. Jackson, M. D. Go, P. Spitale, C. Bunten, M. Leiva, D. Gonzales, J. Hollister-Smith, M. Durand, B. Frei, A. S. Buist, D. Peters, C. D. Morris and E. R. Spindel, Vitamin C supplementation for pregnant smoking women and pulmonary function in their newborn infants: a randomized clinical trial, JAMA, 2014, 311, 2074–2082 CrossRef PubMed.
- H. Maazi, S. Shirinbak, N. Bloksma, M. C. Nawijn and A. J. van Oosterhout, Iron administration reduces airway hyperreactivity and eosinophilia in a mouse model of allergic asthma, Clin. Exp. Immunol., 2011, 166, 80–86 CrossRef CAS PubMed.
- M. Gharagozloo, M. Karimi and Z. Amirghofran, Double-faced cell-mediated immunity in beta-thalassemia major: stimulated phenotype versus suppressed activity, Ann. Hematol., 2009, 88, 21–27 CrossRef CAS PubMed.
- G. Pardalos, F. Kanakoudi-Tsakalidis, M. Malaka-Zafiriu, H. Tsantali, M. Athanasiou-Metaxa, G. Kallinikos and G. Papaevangelou, Iron-related disturbances of cell-mediated immunity in multitransfused children with thalassemia major, Clin. Exp. Immunol., 1987, 68, 138–145 CAS.
- E. Jensen-Jarolim and E. Untersmayr, Gender-medicine aspects in allergology, Allergy, 2008, 63, 610–615 CrossRef CAS PubMed.
- U. Langen, R. Schmitz and H. Steppuhn, Prevalence of allergic diseases in Germany: results of the German Health Interview and Examination Survey for Adults (DEGS1), Bundesgesundheitsblatt Gesundheitsforschung Gesundheitsschutz, 2013, 56, 698–706 CrossRef CAS PubMed.
- C. Fonseca, F. Marques, A. Robalo Nunes, A. Belo, D. Brilhante and J. Cortez, Prevalence of anaemia and iron deficiency in Portugal: the EMPIRE study, Intern. Med. J., 2016, 46, 470–478 CrossRef CAS PubMed.
- G. Alvarez-Uria, P. K. Naik, M. Midde, P. S. Yalla and R. Pakam, Prevalence and severity of anaemia stratified by age and gender in rural India, Anemia, 2014, 2014, 176182 CrossRef PubMed.
- E. Jensen-Jarolim, M. C. Turner and S. N. Karagiannis, AllergoOncology: IgE- and IgG4-mediated immune mechanisms linking allergy with cancer and their translational implications, J. Allergy Clin. Immunol., 2017, 140, 982–984 CrossRef PubMed.
- E. Jensen-Jarolim, G. Achatz, M. C. Turner, S. Karagiannis, F. Legrand, M. Capron, M. L. Penichet, J. A. Rodriguez, A. G. Siccardi, L. Vangelista, A. B. Riemer and H. Gould, AllergoOncology: the role of IgE-mediated allergy in cancer, Allergy, 2008, 63, 1255–1266 CrossRef CAS PubMed.
- A. C. Chua, M. W. Knuiman, D. Trinder, M. L. Divitini and J. K. Olynyk, Higher concentrations of serum iron and transferrin saturation but not serum ferritin are associated with cancer outcomes, Am. J. Clin. Nutr., 2016, 104, 736–742 CrossRef CAS PubMed.
- H. Cao, C. Wang, R. Chai, Q. Dong and S. Tu, Iron intake, serum iron indices and risk of colorectal adenomas: a meta-analysis of observational studies, Eur. J. Cancer Care, 2017, 26, e12486 CrossRef PubMed.
- C. P. Wen, J. H. Lee, Y. P. Tai, C. Wen, S. B. Wu, M. K. Tsai, D. P. Hsieh, H. C. Chiang, C. A. Hsiung, C. Y. Hsu and X. Wu, High serum iron is associated with increased cancer risk, Cancer Res., 2014, 74, 6589–6597 CrossRef CAS PubMed.
- A. G. Mainous, 3rd, J. M. Gill and C. J. Everett, Transferrin saturation, dietary iron intake, and risk of cancer, Ann. Fam. Med., 2005, 3, 131–137 CrossRef PubMed.
- A. G. Mainous, 3rd, B. J. Wells, R. J. Koopman, C. J. Everett and J. M. Gill, Iron, lipids, and risk of cancer in the Framingham Offspring cohort, Am. J. Epidemiol., 2005, 161, 1115–1122 CrossRef PubMed.
- B. J. Wells, A. G. Mainous, 3rd, C. J. Everett and J. M. Gill, Iron, cholesterol, and the risk of cancer in an 18-year cohort, Asian Pac. J. Cancer Prev., 2005, 6, 505–509 Search PubMed.
- A. Sica and A. Mantovani, Macrophage plasticity and polarization: in vivo veritas, J. Clin. Invest., 2012, 122, 787–795 CAS.
- J. A. Thorson, K. M. Smith, F. Gomez, P. W. Naumann and J. D. Kemp, Role of iron in T cell activation: TH1 clones differ from TH2 clones in their sensitivity to inhibition of DNA synthesis caused by IgG Mabs against the transferrin receptor and the iron chelator deferoxamine, Cell. Immunol., 1991, 134, 126–137 CrossRef CAS PubMed.
- S. Leung, A. Holbrook, B. King, H. T. Lu, V. Evans, N. Miyamoto, C. Mallari, S. Harvey, D. Davey, E. Ho, W. W. Li, J. Parkinson, R. Horuk, S. Jaroch, M. Berger, W. Skuballa, C. West, R. Pulk, G. Phillips, J. Bryant, B. Subramanyam, C. Schaefer, H. Salamon, E. Lyons, D. Schilling, H. Seidel, J. Kraetzschmar, M. Snider and D. Perez, Differential inhibition of inducible T cell cytokine secretion by potent iron chelators, J. Biomol. Screening, 2005, 10, 157–167 CrossRef CAS PubMed.
- G. Corna, L. Campana, E. Pignatti, A. Castiglioni, E. Tagliafico, L. Bosurgi, A. Campanella, S. Brunelli, A. A. Manfredi, P. Apostoli, L. Silvestri, C. Camaschella and P. Rovere-Querini, Polarization dictates iron handling by inflammatory and alternatively activated macrophages, Haematologica, 2010, 95, 1814–1822 CrossRef CAS PubMed.
- J. W. Larrick and P. Cresswell, Transferrin receptors on human B and T lymphoblastoid cell lines, Biochim. Biophys. Acta, 1979, 583, 483–490 CrossRef CAS.
- P. A. Seligman, J. Kovar, R. B. Schleicher and E. W. Gelfand, Transferrin-independent iron uptake supports B lymphocyte growth, Blood, 1991, 78, 1526–1531 CAS.
- G. Li, E. J. Pone, D. C. Tran, P. J. Patel, L. Dao, Z. Xu and P. Casali, Iron inhibits activation-induced cytidine deaminase enzymatic activity and modulates immunoglobulin class switch DNA recombination, J. Biol. Chem., 2012, 287, 21520–21529 CrossRef CAS PubMed.
- Y. S. Jang, G. Y. Seo, J. M. Lee, H. Y. Seo, H. J. Han, S. J. Kim, B. R. Jin, H. J. Kim, S. R. Park, K. J. Rhee, W. S. Kim and P. H. Kim, Lactoferrin causes IgA and IgG2b isotype switching through betaglycan binding and activation of canonical TGF-beta signaling, Mucosal Immunol., 2015, 8, 906–917 CrossRef CAS PubMed.
- K. Nakashima, T. Takeuchi and T. Shirakawa, Differentiation, distribution, and chemical state of intracellular trace elements in LAD2 mast cell line, Biol. Trace Elem. Res., 2005, 108, 105–114 CrossRef CAS PubMed.
- M. Shalit, A. Tedeschi, A. Miadonna and F. Levi-Schaffer, Desferal (desferrioxamine)–a novel activator of connective tissue-type mast cells, J. Allergy Clin. Immunol., 1991, 88, 854–860 CrossRef CAS PubMed.
- S. Higa, T. Hirano, M. Kotani, M. Matsumoto, A. Fujita, M. Suemura, I. Kawase and T. Tanaka, Fisetin, a flavonol, inhibits TH2-type cytokine production by activated human basophils, J. Allergy Clin. Immunol., 2003, 111, 1299–1306 CrossRef CAS PubMed.
- A. Singh, A. Demont, L. Actis-Goretta, S. Holvoet, A. Leveques, M. Lepage, S. Nutten and A. Mercenier, Identification of epicatechin as one of the key bioactive constituents of polyphenol-enriched extracts that demonstrate an anti-allergic effect in a murine model of food allergy, Br. J. Nutr., 2014, 112, 358–368 CrossRef CAS PubMed.
- D. F. Finn and J. J. Walsh, Twenty-first century mast cell stabilizers, Br. J. Pharmacol., 2013, 170, 23–37 CrossRef CAS PubMed.
- S. K. Kritas, A. Saggini, G. Varvara, G. Murmura, A. Caraffa, P. Antinolfi, E. Toniato, A. Pantalone, G. Neri, S. Frydas, M. Rosati, M. Tei, A. Speziali, R. Saggini, F. Pandolfi, G. Cerulli, T. C. Theoharides and P. Conti, Luteolin inhibits mast cell-mediated allergic inflammation, J. Biol. Regul. Homeostatic Agents, 2013, 27, 955–959 CAS.
- Y. Shi, J. Dai, H. Liu, R. R. Li, P. L. Sun, Q. Du, L. L. Pang, Z. Chen and K. S. Yin, Naringenin inhibits allergen-induced airway inflammation and airway responsiveness and inhibits NF-kappaB activity in a murine model of asthma, Can. J. Physiol. Pharmacol., 2009, 87, 729–735 CrossRef CAS PubMed.
- M. J. Bae, H. S. Shin, H. J. See, S. Y. Jung, D. A. Kwon and D. H. Shon, Baicalein induces CD4(+)Foxp3(+) T cells and enhances intestinal barrier function in a mouse model of food allergy, Sci. Rep., 2016, 6, 32225 CrossRef CAS PubMed.
- H. Kang, C. H. Lee, J. R. Kim, J. Y. Kwon, M. J. Son, J. E. Kim and K. W. Lee, Theobroma cacao extract attenuates the development of Dermatophagoides farinae-induced atopic dermatitis-like symptoms in NC/Nga mice, Food Chem., 2017, 216, 19–26 CrossRef CAS PubMed.
- J. K. Choi and S. H. Kim, Rutin suppresses atopic dermatitis and allergic contact dermatitis, Exp. Biol. Med., 2013, 238, 410–417 CrossRef PubMed.
- W. Zhou and X. Nie, Afzelin attenuates asthma phenotypes by downregulation of GATA3 in a murine model of asthma, Mol. Med. Rep., 2015, 12, 71–76 CrossRef CAS PubMed.
- D. Y. Zhou, S. R. Fang, C. F. Zou, Q. Zhang and W. Gu, Proanthocyanidin from grape seed extract inhibits airway inflammation and remodeling in a murine model of chronic asthma, Nat. Prod. Commun., 2015, 10, 257–262 Search PubMed.
- R. R. Li, L. L. Pang, Q. Du, Y. Shi, W. J. Dai and K. S. Yin, Apigenin inhibits allergen-induced airway inflammation and switches immune response in a murine model of asthma, Immunopharmacol. Immunotoxicol., 2010, 32, 364–370 CrossRef CAS PubMed.
- S. Masuda, M. Maeda-Yamamoto, S. Usui and T. Fujisawa, ‘Benifuuki’ green tea containing O-methylated catechin reduces symptoms of Japanese cedar pollinosis: a randomized, double-blind, placebo-controlled trial, Allergol. Int., 2014, 63, 211–217 CrossRef CAS PubMed.
- A. Patrizi, B. Raone, I. Neri, C. Gurioli, M. Carbonara, N. Cassano and G. A. Vena, Randomized, controlled, double-blind clinical study evaluating the safety and efficacy of MD2011001 cream in mild-to-moderate atopic dermatitis of the face and neck in children, adolescents and adults, J. Dermatol. Treat., 2016, 27, 346–350 CrossRef CAS PubMed.
- T. Virtanen, Lipocalin allergens, Allergy, 2001, 56(suppl 67), 48–51 CrossRef PubMed.
- H. Breiteneder and E. N. Mills, Molecular properties of food allergens, J. Allergy Clin. Immunol., 2005, 115, 14–23 CrossRef CAS PubMed , quiz 24.
- G. A. Stewart and P. J. Thompson, The biochemistry of common aeroallergens, Clin. Exp. Allergy, 1996, 26, 1020–1044 CrossRef CAS PubMed.
- F. Roth-Walter, M. C. Berin, P. Arnaboldi, C. R. Escalante, S. Dahan, J. Rauch, E. Jensen-Jarolim and L. Mayer, Pasteurization of milk proteins promotes allergic sensitization by enhancing uptake through Peyer's patches, Allergy, 2008, 63, 882–890 CrossRef CAS PubMed.
- D. R. Flower, A. C. North and T. K. Attwood, Structure and sequence relationships in the lipocalins and related proteins, Protein Sci., 1993, 2, 753–761 CrossRef CAS PubMed.
- D. R. Flower, A. C. North and C. E. Sansom, The lipocalin protein family: structural and sequence overview, Biochim. Biophys. Acta, 2000, 1482, 9–24 CrossRef CAS.
- C. Jost and A. Pluckthun, Engineered proteins with desired specificity: DARPins, other alternative scaffolds and bispecific IgGs, Curr. Opin. Struct. Biol., 2014, 27, 102–112 CrossRef CAS PubMed.
- T. Schafer, J. Merkl, E. Klemm, H. E. Wichmann and J. Ring, We and our pets: allergic together?, Acta Vet. Hung., 2008, 56, 153–161 CrossRef PubMed.
- M. A. Holmes, W. Paulsene, X. Jide, C. Ratledge and R. K. Strong, Siderocalin (Lcn 2) also binds carboxymycobactins, potentially defending against mycobacterial infections through iron sequestration, Structure, 2005, 13, 29–41 CrossRef CAS PubMed.
- J. J. Rodvold, N. R. Mahadevan and M. Zanetti, Lipocalin 2 in cancer: when good immunity goes bad, Cancer Lett., 2012, 316, 132–138 CrossRef CAS PubMed.
- K. M. Schmidt-Ott, K. Mori, J. Y. Li, A. Kalandadze, D. J. Cohen, P. Devarajan and J. Barasch, Dual action of neutrophil gelatinase-associated lipocalin, J. Am. Soc. Nephrol., 2007, 18, 407–413 CrossRef CAS PubMed.
- V. Catalan, J. Gomez-Ambrosi, A. Rodriguez, B. Ramirez, C. Silva, F. Rotellar, M. J. Gil, J. A. Cienfuegos, J. Salvador and G. Fruhbeck, Increased adipose tissue expression of lipocalin-2 in obesity is related to inflammation and matrix metalloproteinase-2 and metalloproteinase-9 activities in humans, J. Mol. Med., 2009, 87, 803–813 CrossRef CAS PubMed.
- T. H. Flo, K. D. Smith, S. Sato, D. J. Rodriguez, M. A. Holmes, R. K. Strong, S. Akira and A. Aderem, Lipocalin 2 mediates an innate immune response to bacterial infection by sequestrating iron, Nature, 2004, 432, 917–921 CrossRef CAS PubMed.
- L. R. Devireddy, D. O. Hart, D. H. Goetz and M. R. Green, A mammalian siderophore synthesized by an enzyme with a bacterial homolog involved in enterobactin production, Cell, 2010, 141, 1006–1017 CrossRef CAS PubMed.
- R. C. Hider and X. Kong, Chemistry and biology of siderophores, Nat. Prod. Rep., 2010, 27, 637–657 RSC.
- S. M. Barry and G. L. Challis, Recent advances in siderophore biosynthesis, Curr. Opin. Chem. Biol., 2009, 13, 205–215 CrossRef CAS PubMed.
- M. Guo, C. Perez, Y. Wei, E. Rapoza, G. Su, F. Bou-Abdallah and N. D. Chasteen, Iron-binding properties of plant phenolics and cranberry's bio-effects, Dalton Trans., 2007, 4951–4961, 10.1039/b705136k.
- P. Mladenka, K. Macakova, T. Filipsky, L. Zatloukalova, L. Jahodar, P. Bovicelli, I. P. Silvestri, R. Hrdina and L. Saso, In vitro analysis of iron chelating activity of flavonoids, J. Inorg. Biochem., 2011, 105, 693–701 CrossRef CAS PubMed.
- S. G. Parkar, D. E. Stevenson and M. A. Skinner, The potential influence of fruit polyphenols on colonic microflora and human gut health, Int. J. Food Microbiol., 2008, 124, 295–298 CrossRef CAS PubMed.
- F. Roth-Walter, C. Gomez-Casado, L. F. Pacios, N. Mothes-Luksch, G. A. Roth, J. Singer, A. Diaz-Perales and E. Jensen-Jarolim, Bet v 1 from Birch Pollen is a Lipocalin-like Protein acting as Allergen only when devoid of Iron by promoting Th2 lymphocytes, J. Biol. Chem., 2014, 289, 17416–17421 CrossRef CAS PubMed.
- F. Roth-Walter, L. F. Pacios, C. Gomez-Casado, G. Hofstetter, G. A. Roth, J. Singer, A. Diaz-Perales and E. Jensen-Jarolim, The major cow milk allergen Bos d 5 manipulates T-helper cells depending on its load with siderophore-bound iron, PLoS One, 2014, 9, e104803 Search PubMed.
- C. Seutter von Loetzen, T. Hoffmann, M. J. Hartl, K. Schweimer, W. Schwab, P. Rosch and O. Hartl-Spiegelhauer, Secret of the major birch pollen allergen Bet v 1: identification of the physiological ligand, Biochem. J., 2014, 457, 379–390 CrossRef CAS PubMed.
- J. Vesic, I. Stambolic, D. Apostolovic, M. Milcic and D. Stanic-Vucinic, and T. Cirkovic Velickovic, Complexes of green tea polyphenol, epigalocatechin-3-gallate, and 2S albumins of peanut, Food Chem., 2015, 185, 309–317 CrossRef CAS PubMed.
- E. L. van Boxtel, L. A. van den Broek, S. J. Koppelman, J. P. Vincken and H. Gruppen, Peanut allergen Ara h 1 interacts with proanthocyanidins into higher molecular weight complexes, J. Agric. Food Chem., 2007, 55, 8772–8778 CrossRef CAS PubMed.
- A. Casanal, U. Zander, F. Dupeux, V. Valpuesta and J. A. Marquez, Purification, crystallization and preliminary X-ray analysis of the strawberry allergens Fra a 1E and Fra a 3 in the presence of catechin, Acta Crystallogr., Sect. F: Struct. Biol. Cryst. Commun., 2013, 69, 510–514 CrossRef CAS PubMed.
- G. M. Escandar and L. F. Sala, Complexing behavior of rutin and quercetin, Can. J. Chem., 1991, 69, 1994–2001 CrossRef CAS.
- E. Jensen-Jarolim, L. F. Pacios, R. Bianchini, G. Hofstetter and F. Roth-Walter, Structural similarities of human and mammalian lipocalins, and their function in innate immunity and allergy, Allergy, 2016, 71, 286–294 CrossRef CAS PubMed.
- D. P. Strachan, Hay fever, hygiene, and household size, BMJ, 1989, 299, 1259–1260 CrossRef CAS PubMed.
- E. von Mutius and D. Vercelli, Farm living: effects on childhood asthma and allergy, Nat. Rev. Immunol., 2010, 10, 861–868 CrossRef CAS PubMed.
- M. Rajilic-Stojanovic and W. M. de Vos, The first 1000 cultured species of the human gastrointestinal microbiota, FEMS Microbiol. Rev., 2014, 38, 996–1047 CrossRef CAS PubMed.
- J. Qin, R. Li, J. Raes, M. Arumugam, K. S. Burgdorf, C. Manichanh, T. Nielsen, N. Pons, F. Levenez, T. Yamada, D. R. Mende, J. Li, J. Xu, S. Li, D. Li, J. Cao, B. Wang, H. Liang, H. Zheng, Y. Xie, J. Tap, P. Lepage, M. Bertalan, J. M. Batto, T. Hansen, D. Le Paslier, A. Linneberg, H. B. Nielsen, E. Pelletier, P. Renault, T. Sicheritz-Ponten, K. Turner, H. Zhu, C. Yu, S. Li, M. Jian, Y. Zhou, Y. Li, X. Zhang, S. Li, N. Qin, H. Yang, J. Wang, S. Brunak, J. Dore, F. Guarner, K. Kristiansen, O. Pedersen, J. Parkhill, J. Weissenbach, H. I. T. C. Meta, P. Bork, S. D. Ehrlich and J. Wang, A human gut microbial gene catalogue established by metagenomic sequencing, Nature, 2010, 464, 59–65 CrossRef CAS PubMed.
- R. Balfour Sartor, Bacteria in Crohn's disease: mechanisms of inflammation and therapeutic implications, J. Clin. Gastroenterol., 2007, 41(suppl 1), S37–S43 CrossRef PubMed.
- K. S. Josefsdottir, M. T. Baldridge, C. S. Kadmon and K. Y. King, Antibiotics impair murine hematopoiesis by depleting the intestinal microbiota, Blood, 2017, 129, 729–739 CrossRef CAS PubMed.
- A. Dostal, C. Lacroix, V. T. Pham, M. B. Zimmermann, C. Del'homme, A. Bernalier-Donadille and C. Chassard, Iron supplementation promotes gut microbiota metabolic activity but not colitis markers in human gut microbiota-associated rats, Br. J. Nutr., 2014, 111, 2135–2145 CrossRef CAS PubMed.
- M. Kalliomaki, P. Kirjavainen, E. Eerola, P. Kero, S. Salminen and E. Isolauri, Distinct patterns of neonatal gut microflora in infants in whom atopy was and was not developing, J. Allergy Clin. Immunol., 2001, 107, 129–134 CrossRef CAS PubMed.
- Z. Ling, Z. Li, X. Liu, Y. Cheng, Y. Luo, X. Tong, L. Yuan, Y. Wang, J. Sun, L. Li and C. Xiang, Altered fecal microbiota composition associated with food allergy in infants, Appl. Environ. Microbiol., 2014, 80, 2546–2554 CrossRef PubMed.
- R. Berni Canani, N. Sangwan, A. T. Stefka, R. Nocerino, L. Paparo, R. Aitoro, A. Calignano, A. A. Khan, J. A. Gilbert and C. R. Nagler, Lactobacillus rhamnosus GG-supplemented formula expands butyrate-producing bacterial strains in food allergic infants, ISME J., 2016, 10, 742–750 CrossRef CAS PubMed.
- M. Candela, S. Rampelli, S. Turroni, M. Severgnini, C. Consolandi, G. De Bellis, R. Masetti, G. Ricci, A. Pession and P. Brigidi, Unbalance of intestinal microbiota in atopic children, BMC Microbiol., 2012, 12, 95 CrossRef CAS PubMed.
- C. C. Chen, K. J. Chen, M. S. Kong, H. J. Chang and J. L. Huang, Alterations in the gut microbiotas of children with food sensitization in early life, Pediatr. Allergy Immunol., 2016, 27, 254–262 CrossRef PubMed.
- K. E. Fujimura, A. R. Sitarik, S. Havstad, D. L. Lin, S. Levan, D. Fadrosh, A. R. Panzer, B. LaMere, E. Rackaityte, N. W. Lukacs, G. Wegienka, H. A. Boushey, D. R. Ownby, E. M. Zoratti, A. M. Levin, C. C. Johnson and S. V. Lynch, Neonatal gut microbiota associates with childhood multisensitized atopy and T cell differentiation, Nat. Med., 2016, 22, 1187–1191 CrossRef CAS PubMed.
- M. Rajilic-Stojanovic, H. G. Heilig, S. Tims, E. G. Zoetendal and W. M. de Vos, Long-term monitoring of the human intestinal microbiota composition, Environ. Microbiol., 2013, 15, 1146–1159 CrossRef CAS PubMed.
- X. Hua, J. J. Goedert, A. Pu, G. Yu and J. Shi, Allergy associations with the adult fecal microbiota: analysis of the American Gut Project, EBioMedicine, 2016, 3, 172–179 CrossRef PubMed.
- N. Fyhrquist, L. Ruokolainen, A. Suomalainen, S. Lehtimaki, V. Veckman, J. Vendelin, P. Karisola, M. Lehto, T. Savinko, H. Jarva, T. U. Kosunen, J. Corander, P. Auvinen, L. Paulin, L. von Hertzen, T. Laatikainen, M. Makela, T. Haahtela, D. Greco, I. Hanski and H. Alenius, Acinetobacter species in the skin microbiota protect against allergic sensitization and inflammation, J. Allergy Clin. Immunol., 2014, 134, 1301–1309 CrossRef CAS PubMed e1311.
- E. A. Kennedy, J. Connolly, J. O. Hourihane, P. G. Fallon, W. H. McLean, D. Murray, J. H. Jo, J. A. Segre, H. H. Kong and A. D. Irvine, Skin microbiome before development of atopic dermatitis: Early colonization with commensal staphylococci at 2 months is associated with a lower risk of atopic dermatitis at 1 year, J. Allergy Clin. Immunol., 2017, 139, 166–172 CrossRef PubMed.
- J. Jalanka-Tuovinen, A. Salonen, J. Nikkila, O. Immonen, R. Kekkonen, L. Lahti, A. Palva and W. M. de Vos, Intestinal microbiota in healthy adults: temporal analysis reveals individual and common core and relation to intestinal symptoms, PLoS One, 2011, 6, e23035 CAS.
- J. Jalanka-Tuovinen, J. Salojarvi, A. Salonen, O. Immonen, K. Garsed, F. M. Kelly, A. Zaitoun, A. Palva, R. C. Spiller and W. M. de Vos, Faecal microbiota composition and host-microbe cross-talk following gastroenteritis and in postinfectious irritable bowel syndrome, Gut, 2014, 63, 1737–1745 CrossRef PubMed.
- J. A. Shapiro and T. A. Wencewicz, Acinetobactin Isomerization Enables Adaptive Iron Acquisition in Acinetobacter baumannii through pH-Triggered Siderophore Swapping, ACS Infect. Dis., 2016, 2, 157–168 CrossRef CAS PubMed.
- B. H. Baig, I. K. Wachsmuth and G. K. Morris, Utilization of exogenous siderophores by Campylobacter species, J. Clin. Microbiol., 1986, 23, 431–433 CAS.
- D. Jovanovic, N. Ilic, B. Miljkovic-Selimovic, D. Djokic, T. Relic, Z. Tambur, R. Doder and G. Kostic, Campylobacter jejuni infection and IgE sensitization in up to 2-year-old infants, Vojnosanit. Pregl., 2015, 72, 140–147 CrossRef PubMed.
| This journal is © The Royal Society of Chemistry 2017 |