
Stabilization of hairpins and bulged secondary structures of nucleic acids by single incorporation of α,β-D-CNA featuring a gauche(+) alpha torsional angle†
Béatrice Gerland,
Pierre Millard,
Christelle Dupouy,
Brice-Loïc Renard and
Jean-Marc Escudier*
Laboratoire de Synthèse et Physico-Chimie de Molécules d'Intérêt Biologique, UMR 5068 CNRS, Université Paul Sabatier, 118 Route de Narbonne, 31062 Toulouse Cedex 9, France. E-mail: escudier@chimie.ups-tlse.fr; Fax: +33-5-6155-6011; Tel: +33-5-6155-6291
First published on 12th September 2014
Abstract
A constrained dinucleotide unit featuring a gauche(+) alpha torsional angle configuration was used to stabilize DNA hairpin or bulged structures. Large five nucleotides having looped hairpin structures can be stabilized up to +5 °C. Depending on the nature of the closing base pair, the increased hairpin stability can be reached through loop moiety preorganization or stem rearrangement induced on the first two base pairs. With an alpha gauche(+) constraint either within or opposite to the bulge, the larger bulged structures had better stabilization.
Introduction
Constrained nucleotides have been introduced and developed with the aim to increase duplex formation ability mostly in the context of antisense applications or antigen therapies.1 Despite their biologically relevant functions, secondary nucleic acid structures, such as hairpins, bulges or junctions have received less attention in terms of the design of the modified nucleotides that are able to preorganize and stabilize them.2It is also well established that these disparate structures, which are predisposed to promote a significant local conformational heterogeneity in sugar/phosphate backbones, play a crucial role in fundamental biological processes where protein–nucleic acid interactions, folding, or catalytic activity are involved. As proposed by few studies, the sugar/phosphate backbone of these unusual motifs exhibits a variety of conformations, which markedly differ from the regular conformational states of the nucleic acid duplex.3 Unfortunately, experimental studies aimed at determining the structural and functional implications of such structural deformations are slightly complicated by the intrinsically transient nature of the corresponding backbone states. Stable structural analogues of these distorted backbone geometries would be very useful in elucidating the role that uncanonical conformations play in nucleic acid interactions and folding.
We have engaged in a program towards the design and synthesis of structural analogues of nucleosides in which the sugar/phosphate backbone is constrained at selected positions by a dioxaphosphorinane structure that allowed us to fix the values of two or more torsional angles (Fig. 1).4 Among the synthesized dioxaphosphorinane-constrained nucleic acid dinucleotides (D-CNAs), those featuring B-DNA canonical values for the torsional angles showed an improved duplex formation ability.5 The behaviour of these constrained analogues was easy to understand in this study according to the preorganization concept,8 and then could be selected as unnatural nucleoside analogs for antisense therapy.9 However, many of the D-CNAs exhibited conformations that strongly differ from those necessary to fit with the duplex structure (Fig. 1).10 Therefore, the preorganization concept could be extended to nucleic acid secondary structures in which the sugar/phosphate backbone is necessarily distorted to ensure functional folding.
 | ||
| Fig. 1 Left: the six backbone torsion angles (labeled α to ζ) of the nucleic acids. Middle: the α,β-D-CNA dinucleotide in which α and β are stereocontrolled by a dioxaphosphorinane ring structure. Right: superimposition of the X-ray structures of α,β-D-CNA TT featuring an alpha gauche(+) configuration (gold)6 and of unmodified TPT (black).7 | ||
Bulges and hairpin loops are some of the most important structural motifs in folded nucleic acids that can be of functional importance as ligand recognition elements or folding initiation sites or that may be created in DNA recombination or replication processes.11 These bulged motifs consist of Watson-Crick base paired stem structures and a loop or bulge sequence with unpaired or non-canonical (WC) paired nucleotides formed when one or more nucleotides cannot have regular hydrogen bonding with counterparts in the opposite strands.
Similar to the hairpin loop structure, the directionality of the sugar/phosphate backbones in the nucleic acid bulges can be sharply changed in both strands of the structure as depicted in a five nucleotide DNA bulge structure.12 The bulge induces a sharp kink in the structure, and the torsional stress within the loop of the bulge is located at one specific phosphate, which prevents base stacking at the kinking site. Moreover, on the opposite strand, the kink induced between the two stems of the structure is also ensured by the phosphate connecting the stem residues that are still involved in the Watson-Crick base pairing. Therefore, according to the preorganization concept,8 it appeared that a constrained dinucleotide featuring an alpha torsional angle deviating from the normal range of −62 ± 15° (gauche(−)) observed in B-DNA13 to a gauche(+) conformation (Fig. 1) could turn the phosphate located either in the loop moiety or within the strand opposite to the bulge. Therefore, it should be suitable for the preorganisation of a single strand and, as a consequence, should induce stabilization of the secondary structures.
We have already communicated that the dioxaphosphorinane-constrained nucleic acid dinucleotide (D-CNA) featuring a non-canonical gauche(+) alpha torsional angle value installed in the four nucleotide loop moiety of a hairpin structure was the first constrained nucleotide analogue able to stabilize the overall secondary structure (ΔTm > +3 °C).14 However, this seminal study showed that the effect was dependant on both the position of the strain within the unpaired strand and on the nature of the AT or CG loop-closing base pair.
Herein, in order to gain insight into the stem rearrangement observed only in the case of a hairpin with a CG closing base pair, we studied four thymidine-looped hairpins with stems composed of seven to two base pairs. Then, we present the effect of the application of the gauche(+) constraint to the alpha torsional angle in a larger loop made of five thymidines with either an AT or CG closing base pair. Eventually, we introduced the gauche(+) torsional stress in or opposite to bulges of various sizes in double stranded oligonucleotides. With these investigations, we aim to understand and determine the rules that govern the use of conformationally constrained nucleotides featuring non-B-DNA conformations within unpaired moieties of nucleic acid secondary structures.
Results and discussion
In this report, iHABTj and iHABTjgpx will denote hairpin structures where i is the number of stem base pairs (2 ≤ i ≤ 7), AB is the nature of the closing base pair (AT or CG), j is the number of thymidines within the loop (4 or 5) and x is the position of the alpha gauche(+) torsional angle constraint (gp) within the loop.Study of the effect of gauche(+) constrained D-CNA within the T4 loop on hairpin stem rearrangement
Our preliminary study showed that, within the T4-loop of hairpins with a CG closing base pair, an alpha gauche(+) constraint located in the middle of the unpaired moiety induced a remarkable stabilizing effect of +3.5 °C (Table 1, entry 2 vs. 1) and a similar trend was also observed when the constraint was installed close to the 3′-end of the loop with a ΔTm of +2.7 °C (Table 1, entry 3). In the former case, the effect could be attributed to a preorganization of the loop because it was known that the structure reversed the directionality of the strand in the middle of the unpaired moiety by adopting a gauche(+) constraint on the alpha torsional angle at the so-called turning phosphate.15 On the other hand, the stabilization depicted by shifting the constraint to the 3′-end of the loop was accompanied by a stem rearrangement as evidenced by the circular dichroism analysis. The displacement of the positive cotton band from 278 to 262 nm (Fig. 2, compare 7HCGT4 and 7HCGT4gp3) shows a partial change in the stem from a B- to A-type double helix. Therefore, to gain insight into the behaviour of the stem relative to the torsional stress applied to the sugar/phosphate backbone in these CG-closed loops, we synthesized T4-hairpins with a stem composed of seven to two base pairs. We applied a gauche(+) torsional stress on α either in the middle of the hairpin loop (iHCGT4gp2, 2 ≤ i ≤ 7) or close to the 3′-end (iHCGT4gp3, 2 ≤ i ≤ 7). Thermal denaturation curves and circular dichroism spectra were recorded for the unmodified and constrained hairpins, and the results and spectra are reported in Table 1 and Fig. 2, respectively. It is noteworthy that all the denatured hairpins exhibited the same circular dichroism spectrum with a maximum positive cotton band at about 278 nm (see ESI, Fig. S7–9†).| Entry | Name | Sequencea (5′–3′) | Tmb [°C] | ΔTm [°C] |
|---|---|---|---|---|
a ![[T with combining low line]](https://www.rsc.org/images/entities/b_char_0054_0332.gif) denotes a (SC5′, RP) α,β-D-CNA-modified TT dinucleotide and the italic character denotes a nucleotide within the loop.b Melting temperatures (the average of the Tm values from three experiments, ±1 °C) were measured as the maximum of the first derivative of the UV melting curve (OD260 vs. temperature, 5–80 °C, 0.5 °C min−1), which was recorded at a concentration of 5 μM in a sodium phosphate buffer (10 mM, pH 7.0) containing NaCl (100 mM) and EDTA (1 mM). denotes a (SC5′, RP) α,β-D-CNA-modified TT dinucleotide and the italic character denotes a nucleotide within the loop.b Melting temperatures (the average of the Tm values from three experiments, ±1 °C) were measured as the maximum of the first derivative of the UV melting curve (OD260 vs. temperature, 5–80 °C, 0.5 °C min−1), which was recorded at a concentration of 5 μM in a sodium phosphate buffer (10 mM, pH 7.0) containing NaCl (100 mM) and EDTA (1 mM). |
||||
| 1 | 7HCGT4 | AGGATCCTTTTGGATCCT | 74.0 | — |
| 2 | 7HCGT4gp2 | AGGATCCT![[T with combining low line]](https://www.rsc.org/images/entities/b_i_char_0054_0332.gif) TGGATCCT TGGATCCT |
77.5 | +3.5 |
| 3 | 7HCGT4gp3 | AGGATCCTTGGATCCT |
76.7 | +2.7 |
| 4 | 6HCGT4 | GGATCCTTTTGGATCC | 69.1 | — |
| 5 | 6HCGT4gp2 | GGATCCTTGGATCC |
71.2 | +2.1 |
| 6 | 6HCGT4gp3 | GGATCCTTGGATCC |
71.8 | +2.7 |
| 7 | 5HCGT4 | GATCCTTTTGGATC | 63.0 | — |
| 8 | 5HCGT4gp2 | GATCCTTGGATC |
65.0 | +2.0 |
| 9 | 5HCGT4gp3 | GATCCTTGGATC |
65.1 | +2.1 |
| 10 | 4HCGT4 | ATCCTTTTGGAT | 53.2 | — |
| 11 | 4HCGT4gp2 | ATCCTTGGAT |
56.2 | +2.9 |
| 12 | 4HCGT4gp3 | ATCCTTGGAT |
57.3 | +4.1 |
| 13 | 3HCGT4 | TCCTTTTGGA | nd | — |
| 14 | 3HCGT4gp2 | TCCTTGGA |
nd | nd |
| 15 | 3HCGT4gp3 | TCCTTGGA |
nd | nd |
| 16 | 2HCGT4 | CCTTTTGG | nd | — |
| 17 | 2HCGT4gp2 | CCTTGG |
nd | nd |
| 18 | 2HCGT4gp3 | CCTTGG |
nd | nd |
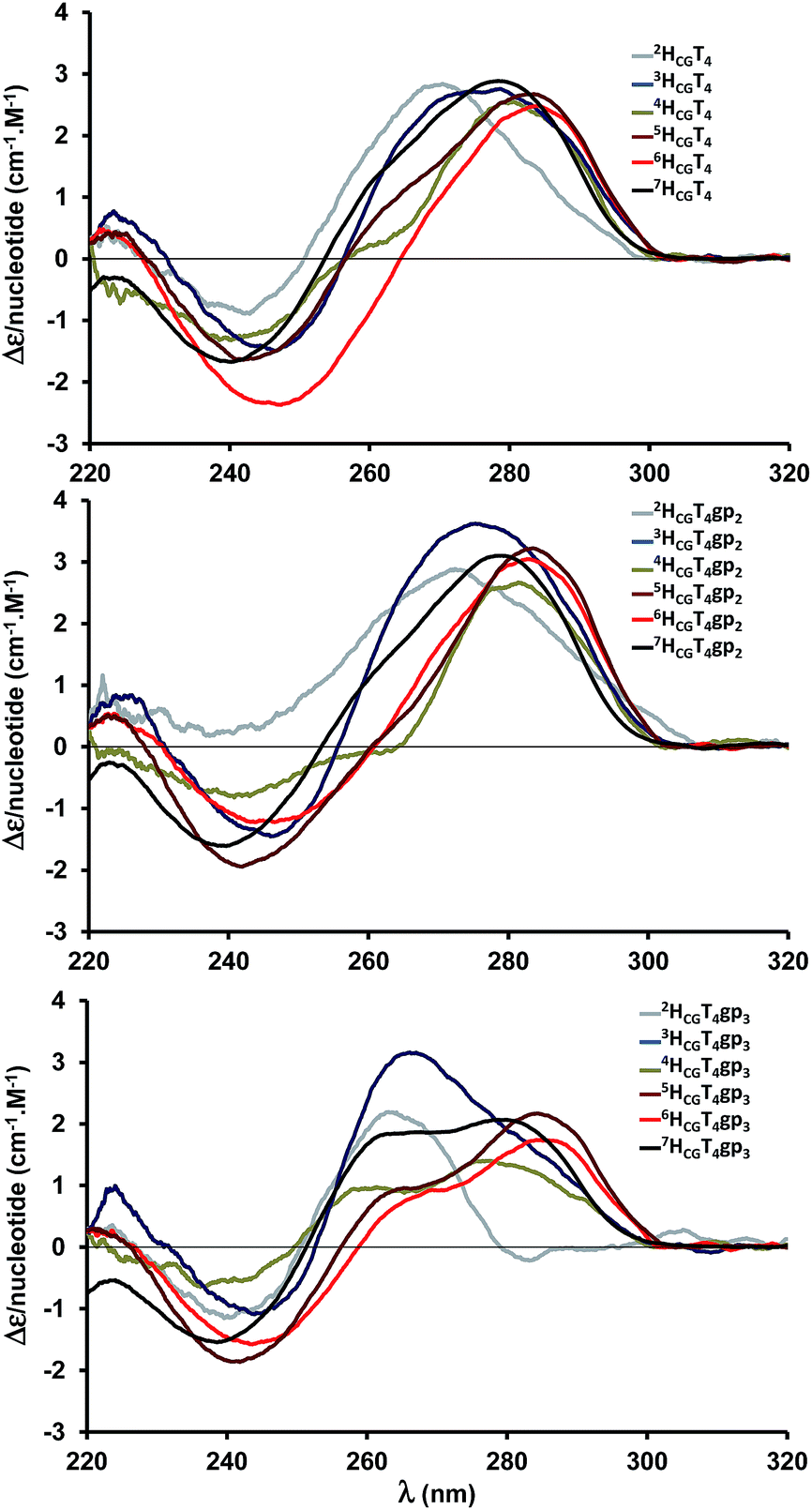 | ||
| Fig. 2 CD spectra of hairpins consisting of a loop with four thymidine residues and stems composed of seven to two base pairs in a sodium phosphate buffer (10 mM, pH 7.0), NaCl (100 mM), and EDTA (1 mM, T = 25 °C). Upper spectrum: unmodified hairpins. Middle spectrum: alpha in gauche(+) conformation at the central position of the loop. Lower spectrum: alpha in gauche(+) conformation at the 3′ end position of the loop. | ||
On shortening the stem length from seven to four base pairs, the melting temperature (Tm), determined by UV-spectrophotometry, decreased from 74.0 °C to 53.2 °C (Table 1, entries 1, 4, 7 and 10). However, it was not possible to observe a transition in the melting curves once the stem length was reduced to two or three base pairs, which disabled the determination of Tm for both the unmodified and constrained hairpins.
Interestingly, the shortened hairpins were nicely stabilized wherever the gauche(+) constraint was applied, as depicted by UV-spectrophotometry for hairpins with stems composed of six to four base pairs. Except for 7HCGT4, in which the main stabilizing effect was the result of a constraint in the middle of the loop (Table 1, entry 2 vs. 3), the predominant effect upon stabilization was established with the constraint close to the 3′-end of the loop (Table 1, iHCGT4gp2 vs. iHCGT4gp3, 6 ≤ i ≤ 4, entry 6 vs. 5, 9 vs. 8 and 12 vs. 11). The induced stem rearrangement was depicted by circular dichroism with the increased displacement of the main band from 280 to 260 nm (Fig. 2 bottom).
The maximum ΔTm value of +4.1 °C that was observed for 4HCGT4gp3 (Table 1, entry 12) led us to speculate that the gauche(+) constraint induced a more efficient stem rearrangement, and consequently a better hairpin stabilization due to the shortening of the hairpin stem.
The underlying question (hypothesis) in this work was whether the torsional stress imposed by the modified D-CNA within the loop could be responsible for the transition from the B- to A-type duplex form in a few of the base pairs of the stem, conferring a higher stability to the hairpin structure.
The UV-thermal denaturation studies were unable to determine the melting temperature for the constrained or unmodified small hairpins 3HCGT4 and 2HCGT4, but circular dichroism appeared to be able to provide information on the number of base pairs in the stem that were affected by the torsional stress imposed by the D-CNA within the loop.
The CD spectra of 3HCGT4 and 3HCGT4gp2 (Fig. 2, upper and middle spectra, blue curves) exhibited a maxima at 276 and 277 nm, respectively, whereas the maximum of the positive band was depicted at 267 nm for 3HCGT4gp3 (Fig. 2, lower spectrum, blue curve). In the latter case, the curve maximum was displaced to 267 nm with a shoulder around 280 nm in a broad band; therefore, it can be speculated that not all the three base pairs were in the A-form. Interestingly, the CD spectrum of 2HCGT4gp3 (Fig. 2, lower spectrum, grey curve) exhibited a narrow band centered at 264 nm, while 2HCGT4 and 2HCGT4gp2 had maxima at about 271 and 273.5 nm, respectively.
The observations made on the CD spectra of the short hairpins, combined with the melting measurements of the longer ones, helped us to propose that the torsional stress formed by the D-CNA modification at the 3′-end of the loop was accompanied by a conformational change of the two closing base pairs from a B- to A-duplex form, thereby effectively stabilizing the hairpin structure independently from a preorganization effect.
Thermostability of gauche(+) constrained D-CNAs within T5 hairpins
It was of interest to investigate the concept of the stabilization of the hairpin structure by a conformational restriction within the unpaired moiety in larger five-looped hairpins. We choose to study hairpins with the same stem sequences as the T4-looped hairpins previously studied with an increment of one thymidine in the loop. Therefore, two different T5-looped hairpins that differ in the nature of the loop-closing base pair AT or CG were investigated in which the alpha gauche(+) constraint has been applied once at all the possible position within the loop and denoted 6HATT5gpi (1 ≤ i ≤ 5) and 7HCGT5gpi (1 ≤ i ≤ 4). The thermal melting temperatures determined by the UV melting curves analysis of the unmodified and constrained hairpins are reported in Table 2.| Entry | Name | Sequencea (5′–3′) | Tmb [°C] | ΔTm [°C] |
|---|---|---|---|---|
| a denotes an (SC5′, RP) α,β-D-CNA-modified TT dinucleotide and the italic character denotes a nucleotide within the loop.b Melting temperatures (the average of the Tm values from three experiments, ±1 °C) were measured as the maximum of the first derivative of the UV melting curve (OD260 vs. temperature, 20–90 °C, 0.5 °C min−1), which was recorded at a concentration of 5 μM in a sodium phosphate buffer (10 mM, pH 7.0) containing NaCl (100 mM) and EDTA (1 mM). |
||||
| 1 | 6HATT5 | ATCCTATTTTTTAGGAT | 50.0 | — |
| 2 | 6HATT5gp1 | ATCCTATTTTAGGAT |
49.0 | −1.0 |
| 3 | 6HATT5gp2 | ATCCTATTTTAGGAT |
52.0 | +2.0 |
| 4 | 6HATT5gp3 | ATCCTATTTTAGGAT |
54.0 | +4.0 |
| 5 | 6HATT5gp4 | ATCCTATTTTAGGAT |
49.5 | −0.5 |
| 6 | 6HATT5gp5 | ATCCTATTTTAGGAT |
44.0 | −6.0 |
| 7 | 7HCGT5 | AGGATCCTTTTTGGATCCT | 68.0 | — |
| 8 | 7HCGT5gp1 | AGGATCCTTTGGATCCT |
68.3 | +0.3 |
| 9 | 7HCGT5gp2 | AGGATCCTTTGGATCCT |
69.8 | +1.8 |
| 10 | 7HCGT5gp3 | AGGATCCTTTGGATCCT |
73.0 | +5.0 |
| 11 | 7HCGT5gp4 | AGGATCCTTTGGATCCT |
70.6 | +2.6 |
In comparison with 6HATT4 and 7HCGT4, 6HATT5 and 7HCGT5 were less stable by 2 and 6 °C, respectively, which underlines the lower stability of the larger loop hairpin structures. Specifically, D-CNA can significantly stabilize T5-looped DNA hairpins by +4.0 and +5.0 °C (Table 2, entries 4 and 10), showing that an appropriate torsional constraint is more effective on a more flexible loop (compare Table 1, entry 2 and Table 2 entry 10).
The modulation of the overall stability of the T5-looped hairpins on replacing one phosphodiester internucleotidic linkage with the rigid dioxaphosphorinane structure was dependent on the nature of the loop closing base pair as previously observed for the T4-looped analogues.
When installed at the 5′- or 3′-end of the AT closed loop, the gauche(+) constraint slightly destabilize the hairpin (Table 2, entries 2 and 5). This constraint was not tolerated at the junction position between the stem and the loop where it induced a strong loss in thermal stability of −6 °C (Table 2, entry 6). These results showed that the T5-loop (6HATT5gpi) structures behave similarly to their T4 counterparts (6HATT4gpi)10 with two favourable positions for the constraint in the middle of the loop that provide an improved thermal stability of +2 and +4 °C (Table 2, entries 3 and 4). All the CD spectra recorded for 6HATT5gpi were identical to those of the unmodified 6HATT5 without any differences (neither in the shape nor in the value of λmax for the positive cotton band at 269 nm) (see ESI, S5†).
The results obtained for 7HCGT5gpi (1 ≤ i ≤ 4) were in line with those of jHCGT4gpi (2 ≤ j ≤ 7; 1 ≤ i ≤ 3), outlining that the torsional stress always induced a notable stabilization with a maximum of +5 °C when the modification was placed at a central position in the loop (Table 2, entry 10). From this main stabilizing position, the ΔTm decreased to +1.8 and +0.3 °C with a shift to the 5′-end of the loop (Table 2, entries 8 and 9), as previously depicted for jHCGT4gpi. The rather important stabilization measured (ΔTm = +2.6 °C) when the D-CNA was installed at the 3′-end of the loop (Table 2, entry 11) was accompanied by a stem rearrangement identified by an additional shoulder at 260 nm in the major positive cotton band at 280 nm within the dichroic spectrum of 7HCGT5gp4 (see ESI, S6†).
Therefore, these results showed that the D-CNA behaviour within hairpin structures was independent of loop and stem sizes.
Thermostability of gauche(+) constrained D-CNA within bulges
In order to further study the impact of D-CNA on partially unpaired DNA structures, we also studied bulges. In these structures, each strand is submitted to a torsional stress that increases with the size of the bulged region. However, the change of the strand orientation in the bulged moiety is rather limited when compared with monomolecular hairpins, since the directionality of the strand is not totally reversed.Two types of bulged structures have been prepared from one D-CNA modified strand 5′-GATTTGCATA![[T with combining low line]](https://www.rsc.org/images/entities/i_char_0054_0332.gif) CATGAG ( denotes an (SC5′, RP) α,β-D-CNA-modified TT dinucleotide). The modification was installed within bulges growing from one to six bases by using complementary strands of 17 to 12 bases, in which bases were removed one by one from the central region (Table 3). Similarly, to generate bulges opposite to the modification, complementary strands of 19 to 24 bases, in which bases were added one by one in the central region were used (Table 4).
CATGAG ( denotes an (SC5′, RP) α,β-D-CNA-modified TT dinucleotide). The modification was installed within bulges growing from one to six bases by using complementary strands of 17 to 12 bases, in which bases were removed one by one from the central region (Table 3). Similarly, to generate bulges opposite to the modification, complementary strands of 19 to 24 bases, in which bases were added one by one in the central region were used (Table 4).
| Entry | Target oligonucleotide 3′ → 5′ | Tm(ΔTm)a [°C] 5′-GATTTGCATATxTCATGAG | |
|---|---|---|---|
| x = PO2− | x = D-CNA g+ | ||
| a Melting temperatures (the average of the Tm values from three experiments, ±1 °C) were measured as the maximum of the first derivative of the UV melting curve (OD260 vs. temperature, 20–90 °C, 0.5 °C min−1), which was recorded at a concentration of 5 μM in a sodium phosphate buffer (10 mM, pH 7.0) containing NaCl (100 mM) and EDTA (1 mM). | |||
| 1 | CTAAACGTATAAGTACTC | 57.0 | 52.6 (−4.4) |
| 2 | CTAAACGTATAGTACTC | 49.0 | 50.0 (+1.0) |
| 3 | CTAAACGTATGTACTC | 43.0 | 45.1 (+2.1) |
| 4 | CTAAACGTAGTACTC | 39.3 | 41.2 (+1.9) |
| 5 | CTAAACGTATACTC | 36.4 | 39.0 (+2.4) |
| 6 | CTAAACGTTACTC | 26.0 | 30.0 (+4.0) |
| 7 | CTAAACGTACTC | 25.0 | 31.0 (+6.0) |
| Entry | Target oligonucleotide 3′ → 5′ | Tm(ΔTm)a [°C] 5′-GATTTGCATATxTCATGAG | |
|---|---|---|---|
| x = PO2− | x = D-CNA g+ | ||
| a Melting temperatures (the average of the Tm values from three experiments, ±1 °C) were measured as the maximum of the first derivative of the UV melting curve (OD260 vs. temperature, 20–90 °C, 0.5 °C min−1), which was recorded at a concentration of 5 μM in a sodium phosphate buffer (10 mM, pH 7.0) containing NaCl (100 mM) and EDTA (1 mM). | |||
| 1 |  |
52.0 | 46.5 (−5.5) |
| 2 |  |
47.2 | 44.2 (−3.0) |
| 3 |  |
45.0 | 43.6 (−1.4) |
| 4 |  |
43.0 | 43.3 (+0.3) |
| 5 |  |
42.0 | 44.0 (+2.0) |
| 6 |  |
39.0 | 43.2 (+4.2) |
In comparison with the fully matched duplex in which the D-CNA induced a strong thermal stability loss of −5.5 °C,16 the alpha gauche(+) torsional stress in single nucleotide bulge already stabilized it by +1 °C (Table 3, entry 1 vs. 2). Increasing the size of the bulge from two to four unpaired nucleotides resulted in moderately stabilized structures with ΔTm values around +2 °C when compared with their corresponding unmodified bulges (Table 3, entries 3–5). Larger bulged moieties of five and six nucleotides took advantage of the D-CNA addition and exhibited higher thermal stabilities than their unconstrained counterparts with ΔTm of +4 and +6 °C, respectively (Table 3, entries 6 and 7). To date, these are the only results to report an effective large stabilization by preorganized nucleotides installed in the unpaired moieties of very unstable bulged secondary DNA structures.
As it was known that the bulges induce a sharp kink and the increase in the degree of kink was roughly proportional to the increase in the number of bases in the bulge from one to six.17 The Tm measurement of the growing bulges opposite to the strand in which the alpha gauche(+) torsional stress was applied by the D-CNA modification would give insight on how such structures can be preorganized and, as a consequence, stabilized.
In these constructions, all of the different bulge loops were flanked by the same helices (11 and 7 bases pairs) that share the D-CNA at their junction (position 7–8 on the constant strand). It could be then postulated that the variations in the melting temperature between the various bulges would reflect the ability of the whole structures to accommodate the loop-induced kink. The results of thermal denaturation experiments obtained for the unmodified and D-CNA-constrained bulges are summarized in Table 4. The main observation was that all the bulges exhibited a similar thermal stability with Tm values around 44 ± 1 °C except for the single nucleotide bulge structure that had a slightly higher Tm of 46.5 °C (Table 4, entries 1 vs. 2–6). It appeared that the alpha gauche(+) torsional stress within the strand opposite to the bulge moiety was not well tolerated by bulged structures with one to three unpaired nucleotides, with ΔTm increasing with the number of unpaired nucleotides (Table 4, entries 1–3). A four nucleotides bulged structure was insensitive to the D-CNA modification with ΔTm = +0.3 °C (Table 4, entry 4). Finally, in comparison with the unmodified five and six nucleotides bulges, those featuring the alpha gauche(+) constraint were more stable by +2 and +4.2 °C, respectively (Table 4 entries 5 and 6). It was interesting to correlate the preorganization of the strand including the D-CNA that imposed a change in its directionality with the kink in the overall structure.
The larger the bulge, the more important the kink and therefore the alpha gauche(+) constraint that deviated from the canonical value by 120°. However, this was only true when at least four nucleotides were unpaired.
The stabilizing effect observed could then be attributed to the lower torsional stress applied to the helices flanking the internal loop in the overall bulged structure because of the entropic gain provided by the D-CNA.
Conclusions
Systematic studies of unusual DNA structures deviating from the double-helical structure of DNA set the groundwork for understanding the principles of nucleic acid architecture and the possible chemical modifications to favour their folding and stability. We reported here how D-CNA dinucleotides featuring the non canonical alpha gauche(+) constraint were suitable for the preorganization and stabilization of the hairpin and bulged structures of nucleic acids. We showed that the gain in thermal stability could be due to either the loop structure or the rearrangement of the closing base pairs within T4-looped hairpins. Modified T5-looped hairpins behaved similarly and were nicely stabilized with ΔTm values up to +5 °C. To the best of our knowledge, we reported the enhanced thermal stability of the bulged structures by a torsional constraint on the sugar/phosphate backbone for the first time. We showed that an alpha gauche(+) constraint was always positive when applied within the loop of 1 to 6 nucleotide bulges, whereas when located opposite to the unpaired moiety, it could be a stabilizing element for bulges reaching four unpaired nucleotides.Among their inertness towards enzymatic degradation such as snake venom phosphodiesterase as expected for phosphotriesters,18 D-CNAs within single stranded DNA oligonucleotides showed their potential as terminators in polymerase chain reactions19 and they were recently also proven to be efficient during allele selective silencing when included in antisense oligonucleotides directed against the huntingtin protein.20 In this report, we further investigated the understanding of how D-CNA, as a structural distorted nucleotide analogue, behaves within oligonucleotides that can fold into secondary structures. Because these non-canonical torsional angle conformations can be observed in protein/DNA complexes, research is in progress about the modulation of the integrase activity with D-CNA modified four-way junctions in recombination biochemical processes, and results will be published in due course. D-CNAs could be suitable elements for the modulation of these interactions and could eventually be the bases for the elaboration of decoys or stabilised synthetic functional DNAs such as aptamers or aptazymes.
Acknowledgements
The authors are grateful to the Agence Nationale de la Recherche for financial support and grants to B. Gerland and B. L. Renard (ANR project Modulation of Integrase Activity by means of Conformationaly Constrained nucleic acids, 11-BS07-012-01).Notes and references
- (a) C. J. Leumann, Bioorg. Med. Chem., 2002, 10, 841 CrossRef CAS; (b) A. J. A. Cobb, Org. Biomol. Chem., 2007, 5, 3260 RSC; (c) J. Lebreton, J.-M. Escudier, L. Arzel and C. Len, Chem. Rev., 2010, 110, 3371 CrossRef CAS PubMed; (d) C. Z. Zhou and J. Chattopadhyaya, Curr. Opin. Drug Discovery Dev., 2009, 12, 876 CAS; (e) N. M. Bell and J. Micklefield, ChemBioChem, 2009, 10, 2691 CrossRef CAS PubMed; (f) T. Yamamoto, M. Nakatani, K. Narukawa and S. Obika, Future Med. Chem., 2011, 3, 339 CrossRef CAS PubMed.
- (a) K. Seio, T. Wada, K. Sakamoto, S. Yokoyama and M. Sekine, J. Org. Chem., 1998, 63, 1429 CrossRef CAS; (b) K. Seio, T. Wada, K. Sakamoto, S. Yokoyama and M. Sekine, J. Org. Chem., 1996, 61, 1500 CrossRef CAS; (c) M. Sekine, O. Kurrasawa, K.-I. Shohda, K. Seio and T. Wada, J. Org. Chem., 2000, 65, 6515 CrossRef CAS; (d) K. Seio, T. Wada and M. Sekine, Helv. Chim. Acta, 2000, 83, 162 CrossRef CAS; (e) A. M. Sørensen and P. Nielsen, Org. Lett., 2000, 2, 4217 CrossRef PubMed; (f) A. M. Sørensen, K. E. Nielsen, B. Vogg, J. P. Jacobsen and P. Nielsen, Tetrahedron, 2001, 57, 10191 CrossRef; (g) P. Børsting and P. Nielsen, Chem. Commun., 2002, 2140 RSC; (h) P. Børsting, A. M. Sørensen and P. Nielsen, Synthesis, 2002, 797 CrossRef; (i) P. Børsting, K. E. Nielsen and P. Nielsen, Org. Biomol. Chem., 2005, 3, 2183 RSC; (j) P. Børsting, M. S. Christensen, S. I. Steffansen and P. Nielsen, Tetrahedron, 2006, 62, 1139 CrossRef PubMed.
- (a) C. M. Duarte and A. M. Pyle, J. Mol. Biol., 1998, 284, 1465 CrossRef CAS PubMed; (b) W. A. Baase, J. Jose, B. C. Ponedel, P. H. von Hippel and N. P. Jonhson, Nucleic Acids Res., 1682, 209, 37 Search PubMed; (c) S. Sharma, J. Nucleic Acids, 2011, 724215 Search PubMed; (d) F. Guo, D. N. Gopaul and G. D. van Duyne, Nature, 1997, 389(6646), 40 CrossRef CAS PubMed; (e) F. Guo, D. N. Gopaul and G. D. van Duyne, J. Biomol. Struct. Dyn., 2000, 141 CrossRef CAS PubMed.
- D.-A. Catana, B.-L. Renard, M. Maurano, C. Payrastre, N. Tarrat and J.-M. Escudier, J. Nucleic Acids, 2012, 215876 Search PubMed.
- C. Dupouy, N. Iché-Tarrat, M.-P. Durrieu, F. Rodriguez, J.-M. Escudier and A. Vigroux, Angew. Chem., Int. Ed., 2006, 45, 3623 CrossRef CAS PubMed.
- (a) I. Le Clezio, J.-M. Escudier and A. Vigroux, Org. Lett., 2003, 5, 161 CrossRef CAS PubMed; (b) M. Maturano, D.-A. Catana, P. Lavedan, N. Tarrat, N. Saffon, C. Payrastre and J.-M. Escudier, Eur. J. Org. Chem., 2012, 721 CrossRef CAS.
- N. Camerman, J. K. Fawcett and A. Camerman, J. Mol. Biol., 1976, 107, 601 CrossRef CAS.
- D. J. Cram, Angew. Chem., Int. Ed., 1988, 27, 1009 CrossRef.
- P. P. Seth and E. E. Swayze, Unnatural Nucleoside Analogs for Antisense Therapy in Natural Products in Medicinal Chemistry, ed. Stephen Hanessian, Wiley-VCH Verlag GmbH & Co. KGaA, 2014 Search PubMed.
- (a) I. Le Clezio, H. Gornitzka, J.-M. Escudier and A. Vigroux, J. Org. Chem., 2005, 70, 1620 CrossRef CAS PubMed; (b) C. Dupouy, P. Lavedan and J.-M. Escudier, Eur. J. Org. Chem., 2007, 5256 CrossRef CAS; (c) I. Le Clezio, A. Vigroux and J.-M. Escudier, Eur. J. Org. Chem., 2007, 1935 CrossRef CAS; (d) C. Dupouy, P. Lavedan and J.-M. Escudier, Tetrahedron, 2007, 63, 11235 CrossRef CAS PubMed; (e) C. Dupouy, P. Lavedan and J.-M. Escudier, Eur. J. Org. Chem., 2008, 1285 CrossRef CAS.
- (a) G. Varani, Annu. Rev. Biophys. Biomol. Struct., 1995, 24, 379 CrossRef CAS PubMed; (b) M. A. Glucksmann-Kuis, X. Dai, P. Markiewicz and L. B. Rothaman-Denes, Cell, 1996, 84, 147 CrossRef CAS; (c) S. Blouin, J. Mulhbacher, J. C. Penedo and D. A. Lafontaine, ChemBioChem, 2009, 10, 400 CrossRef CAS PubMed; (d) R. T. Batey, R. P. Rambo and J. A. Doudna, Angew. Chem., Int. Ed., 1999, 38, 2326 CrossRef; (e) E. A. Doherty and J. A. Doudna, Annu. Rev. Biophys. Biomol. Struct., 2001, 30, 457 CrossRef CAS PubMed; (f) C. A. S. A. Minetti, D. P. Remeta, R. Dickstein and K. J. Breslauer, Nucleic Acids Res., 2010, 38, 97 CrossRef CAS PubMed.
- F. A. Gollmick, M. Lorenz, U. Dornberger, J. von Langen, S. Diekmann and H. Fritzsche, Nucleic Acids Res., 2002, 30, 2669 CrossRef CAS PubMed.
- B. Schneider, S. Neidle and H. M. Berman, Biopolymers, 1997, 42, 113 CrossRef CAS.
- C. Dupouy, P. Millard, A. Boissonnet and J.-M. Escudier, Chem. Commun., 2010, 46, 5142 RSC.
- C. W. Hilbers, H. A. Heus, M. J. P. Van Dongen and S. S. Wijmenga, The hairpin elements of nucleic acid structure: DNA and RNA folding, Nucleics Acids and Molecular Biology, ed. F. Eckstein and D. M. J. Lilley, Springer-Verlag, Berlin Heidelberg, 1994, vol. 8, p. 56 Search PubMed.
- A. Boissonnet, C. Dupouy, P. Millard, M.-P. Durrieu, N. Tarrat and J.-M. Escudier, New J. Chem., 2011, 35, 1528 RSC.
- Y.-H. Wang and J. Griffith, Biochemistry, 1991, 30, 1358 CrossRef CAS.
- C. Zhou, O. Plashkevych and J. Chattopadhyaya, J. Org. Chem., 2009, 74, 3248 CrossRef CAS PubMed.
- O. Martinez, V. Ecochard, S. Mahéo, G. Gross, P. Bodin, J. Teissié, J. M. Escudier and L. Paquereau, PLoS One, 2011, 6, e25510 CAS.
- M. Østergaard, B. Gerland, J.-M. Escudier, E. E. Swayze and P. P. Seth, ACS Chem. Biol., 2014, 9(9), 1975 CrossRef PubMed.
Footnote |
| † Electronic supplementary information (ESI) available: Phosphoramidite and ODN synthesis, thermal denaturation curves and Tm vs [salt] plots. Circular dichroism spectra. See DOI: 10.1039/c4ra09639h |
| This journal is © The Royal Society of Chemistry 2014 |