
A fluorinated ionizable lipid improves the mRNA delivery efficiency of lipid nanoparticles†

Abstract
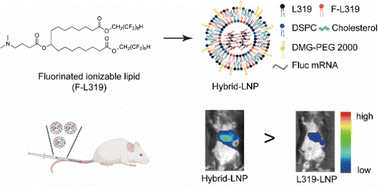
The efficacy of messenger RNA (mRNA)-based vaccines or therapies relies on delivery vehicles that can transport them into the cytosol of cells. Lipid nanoparticles (LNPs) are the most clinically advanced carrier for mRNA. The chemical structure of an ionizable lipid is critical for the delivery efficiency of the LNPs. Herein, we synthesize a new ionizable lipid containing fluorinated alkyl chains (F-L319) and evaluate its mRNA delivery efficiency compared to its hydrocarbon counterpart (L319). While LNPs formulated with F-L319 alone showed decreased mRNA encapsulation and delivery efficiencies in comparison to the L319-LNP, we found that combining the appropriate ratios of F-L319 and L319 as hybrid ionizable lipids in LNPs (hybrid-LNPs) greatly enhanced mRNA delivery efficiency both in vitro and in vivo. Upon intravenous injection, the hybrid-LNP showed targeted mRNA expression in the spleen. Mechanistic studies indicate that the enhanced mRNA delivery of the hybrid-LNP is attributed to both improved mRNA encapsulation and cellular uptake. Collectively, fluorination of ionizable lipids represents a promising strategy to improve the delivery efficiency of LNPs.
- This article is part of the themed collections: 2023 Journal of Materials Chemistry B HOT Papers, Journal of Materials Chemistry B Emerging Investigators, #MyFirstJMCB, 2024 Journal of Materials Chemistry B Lunar New Year, 2023 Journal of Materials Chemistry B Most Popular Articles and mRNA vaccines against COVID-19: Celebrating the 2023 Nobel Prize in Physiology or Medicine
 Please wait while we load your content...
Please wait while we load your content...

