
An artificial antibody for exosome capture by dull template imprinting technology†
 *a
*a
Abstract
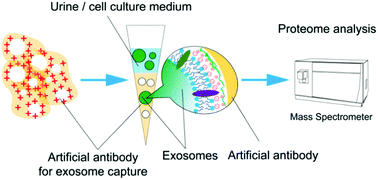
Exosomes are extracellular vesicles with unique size distribution derived from the parent cells. They are involved in intercellular communication and transport, and are also biomarkers for early diagnosis and prognosis of disease. However, the isolation and characterization of exosomes face the challenges of large sample requirements and low enrichment efficiency of traditional methods. Herein, a simple method is proposed for the preparation of an artificial antibody that has a synergistic effect by featuring nanocavities obtained by dull template imprinting and molecular recognition conferred by electrostatic interaction. With this artificial antibody, highly efficient capture and proteome analysis of exosomes from urine and cell culture media are achieved: for urine, the abundance of Top100 exosomal proteins enriched by this artificial antibody increased from 1.85% to 9.66%. For the cell culture medium, the abundance of the Top100 proteins enriched by this artificial antibody was 28.4%. Moreover, this artificial antibody has a comparable effect to the commercial precipitation-based method in capturing exosomes and has advantages in removing contaminants such as prothymosin alpha (PTMA), demonstrating the superior selectivity of the artificial antibody. Overall, this artificial antibody holds promise to capture exosomes from biofluids and is compatible with subsequent proteome analysis.
- This article is part of the themed collection: New era in advanced functional materials emerging from molecular imprinting and related techniques
 Please wait while we load your content...
Please wait while we load your content...

