
Fundamentals and exploration of aggregation-induced emission molecules for amyloid protein aggregation

Abstract
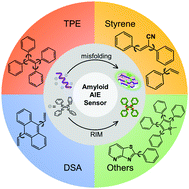
The past decade has witnessed the growing interest and advances in aggregation-induced emission (AIE) molecules as driven by their unique fluorescence/optical properties in particular sensing applications including biomolecule sensing/detection, environmental/health monitoring, cell imaging/tracking, and disease analysis/diagnosis. In sharp contrast to conventional aggregation-caused quenching (ACQ) fluorophores, AIE molecules possess intrinsic advantages for the study of disease-related protein aggregates, but such studies are still at an infant stage with much less scientific exploration. This outlook mainly aims to provide the first systematic summary of AIE-based molecules for amyloid protein aggregates associated with neurodegenerative diseases. Despite a limited number of studies on AIE–amyloid systems, we will survey recent and important developments of AIE molecules for different amyloid protein aggregates of Aβ (associated with Alzheimer's disease), insulin (associated with type 2 diabetes), (α-syn, associated with Parkinson's disease), and HEWL (associated with familial lysozyme systemic amyloidosis) with a particular focus on the working principle and structural design of four types of AIE-based molecules. Finally, we will provide our views on current challenges and future directions in this emerging area. Our goal is to inspire more researchers and investment in this emerging but less explored subject, so as to advance our fundamental understanding and practical design/usages of AIE molecules for disease-related protein aggregates.
- This article is part of the themed collection: Bioinspired Surfaces Engineering for Biomaterials
 Please wait while we load your content...
Please wait while we load your content...

