
Nanoparticle personalized biomolecular corona: implications of pre-existing conditions for immunomodulation and cancer
 a
and
Ryan M.
Pearson
*abc
a
and
Ryan M.
Pearson
*abc
Abstract
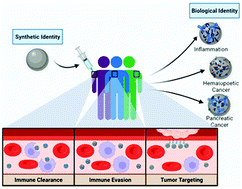
Nanoparticles (NPs) have demonstrated great promise as immunotherapies for applications ranging from cancer, autoimmunity, and infectious disease. Upon encountering biological fluids, NPs rapidly adsorb biomolecules, forming the “biomolecular corona” (BC), and the altered character of NPs due to their newly acquired biological identity can impact their in vivo fate. Recently, it has been shown that the NP–BC is person-specific, and even minute differences in the biomolecule composition can give rise to altered immune recognition, cellular interactions, pharmacokinetics, and biodistribution. Given the current rise in the development of NP-based therapeutics, it is of utmost importance to better understand how pre-existing conditions, that result in the formation of a personalized BC, can be leveraged to aid in the prediction of the therapeutic outcomes of NPs. In this minireview, we will discuss the formation of the BC, implications of the BC for NP-biological interactions, and its clinical importance in the context of immunomodulation and cancer therapeutics.
- This article is part of the themed collection: Biomaterials Science Emerging Investigator Series
 Please wait while we load your content...
Please wait while we load your content...

