
Perturbation of gut microbiota plays an important role in micro/nanoplastics-induced gut barrier dysfunction†

Abstract
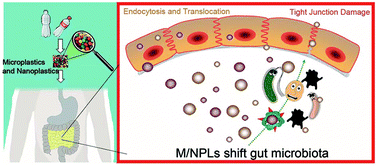
The widespread occurrence of microplastics (MPLs) and nanoplastics (NPLs), collectively abbreviated as M/NPLs, has markedly affected the ecosystem and has become a global threat to human health. Multiple investigations have shown that the chronic ingestion of M/NPLs negatively affects gut barrier function but the mechanism remains unclear. Herein, this research has investigated the toxic effects of pristine polystyrene (PS) M/NPLs, negatively charged carboxylated polystyrene M/NPLs (PS-COOH) and positively charged aminated polystyrene M/NPLs (PS-NH2) of two sizes (70 nm and 5 μm in diameter) in mice. Gavage of these PS M/NPLs for 28 days caused obvious injuries to the gut tract, leading to the decreased expression of tight junction proteins. The toxicity of the M/NPLs was ranked as PS-NH2 > PS-COOH > pristine PS. Oral administration of these M/NPLs resulted in marked gut microbiota dysbiosis. The M/NPLs-enriched genera generally contained opportunistic pathogens which are accompanied by a deteriorated intestinal barrier function, while most M/NPLs-decreased bacteria were beneficial microbes with known tight junction-promoting functions, implicating an important indirect toxic effect of gut microbiota dysbiosis in M/NPLs-induced gut barrier dysfunction. In conclusion, this research highlights the importance of gut microbiota in the toxicity of M/NPLs exposure on gut barrier function, providing novel insights into the adverse effects of M/NPLs exposure on human health.
- This article is part of the themed collections: 2021 Nanoscale HOT Article Collection and Celebrating our 2021 Prizewinners
 Please wait while we load your content...
Please wait while we load your content...

