
Transporting mitochondrion-targeting photosensitizers into cancer cells by low-density lipoproteins for fluorescence-feedback photodynamic therapy†‡
 a
Chunlei
Zhu
*a
a
Chunlei
Zhu
*a
Abstract
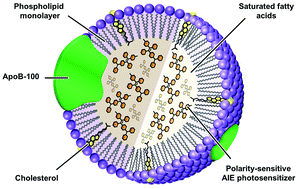
Low-density lipoproteins (LDLs) are an endogenous nanocarrier to transport lipids in vivo. Owing to their biocompatibility and biodegradability, reduced immunogenicity, and natural tumor-targeting capability, we, for the first time, report the reconstitution of native LDL particles with saturated fatty acids and a mitochondrion-targeting aggregation-induced emission (AIE) photosensitizer for fluorescence-feedback photodynamic therapy (PDT). In particular, a novel AIE photosensitizer (TPA-DPPy) with a donor–acceptor (D–A) structure and a pyridinium salt is designed and synthesized, which possesses typical AIE and twisted intramolecular charge transfer (TICT) characteristics as well as reactive oxygen species (ROS)-sensitizing capability. In view of its prominent photophysical and photochemical properties, TPA-DPPy is encapsulated into LDL particles for photodynamic killing of cancer cells that overexpress LDL receptors (LDLRs). The resultant LDL (rLDL) particles maintain a similar morphology and size distribution to native LDL particles, and are efficiently ingested by cancer cells via LDLR-mediated endocytosis, followed by the release of TPA-DPPy for mitochondrion-targeting. Upon light irradiation, the produced ROS surrounding mitochondria lead to efficient and irreversible cell apoptosis. Interestingly, this process can be fluorescently monitored in a real-time fashion, as reflected by the remarkably enhanced luminescence and blue-shifted emission, indicating the increased mechanical stress during apoptosis. Quantitative cell viability analysis suggests that TPA-DPPy exhibits an outstanding phototoxicity toward LDLR-overexpressing A549 cancer cells, with a killing efficiency of ca. 88%. The rLDL particles are a class of safe and multifunctional nanophototheranostic agents, holding great promise in high-quality PDT by providing real-time fluorescence feedback on the therapeutic outcome.
- This article is part of the themed collection: Nanoscale 2021 Emerging Investigators
 Please wait while we load your content...
Please wait while we load your content...

