
A nano perspective behind the COVID-19 pandemic

Abstract
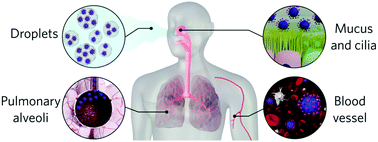
The global pandemic scenario has definitely pushed the scientific community to develop COVID-19 vaccines at unprecedented speed. Nevertheless, a worldwide vaccination campaign is still far from being achieved, making the usual precautionary measures as necessary as at the beginning of the outbreak. Many aspects of the SARS-CoV-2 infectious potential and disease severity do not solely rely on interactions at the molecular level but also on physical–chemical parameters that often involve nanoscale effects. Here the SARS-CoV-2 journey to infect a susceptible host is reviewed, focusing on the nanoscale aspects that play a role in the viral infectivity and disease progression. These nanoscale-driven interactions are essential to establish mitigation-related strategies.
- This article is part of the themed collections: Nanoscale Horizons 10th anniversary regional spotlight collection: The Americas, Coronavirus articles - free to access collection and Horizons Community Board Collection: Antimicrobial materials and surfaces
 Please wait while we load your content...
Please wait while we load your content...

