
Ferroptosis/pyroptosis dual-inductive combinational anti-cancer therapy achieved by transferrin decorated nanoMOF†
 *a
*a
Abstract
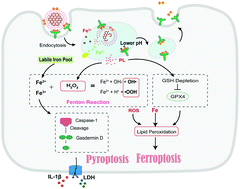
Non-apoptotic cell death such as ferroptosis and pyroptosis has shed new light on cancer treatment, whereas combinational therapy using both these mechanisms has not yet been fully explored. Herein, a dual-inductive nano-system to realize ferroptosis/pyroptosis mediated anti-cancer effects is presented. The nanodrug (Tf-LipoMof@PL) is constructed with a piperlongumine (PL) loaded metal–organic framework (MOF) coated with transferrin decorated pH sensitive lipid layer. Intracellular iron was enriched with an iron-containing MOF, whose endocytosis can be further facilitated by transferrin decorated on the lipid layer, which provides a prerequisite for the occurrence of ferroptosis and pyroptosis. Piperlongumine as the ferroptosis inducer can strengthen the ferroptotic cell death, and provide H2O2 for the dual induction system to increase ROS generation through Fenton reaction. On the basis of validation of both ferroptosis and pyroptosis, the dual-inductive nanodrug demonstrated ideal anticancer effects in the xenograft mice model, which proved that the ferroptosis/pyroptosis dual-inductive nanoplatform could be an effective and promising anticancer modality.
- This article is part of the themed collections: Nanoscale Horizons 10th anniversary regional spotlight collection: China, Nanoscale Horizons 2022 Lunar New Year Collection and Nanoscale Horizons Most Popular 2021 Articles
 Please wait while we load your content...
Please wait while we load your content...

