
Colorimetric loop-mediated isothermal amplification (LAMP) for cost-effective and quantitative detection of SARS-CoV-2: the change in color in LAMP-based assays quantitatively correlates with viral copy number†
Yu Shrike
Zhang,  f
Sergio O.
Martínez-Chapa,
c
Grissel Trujillo-de
Santiago
*ac
and
Mario Moisés
Alvarez
*ab
f
Sergio O.
Martínez-Chapa,
c
Grissel Trujillo-de
Santiago
*ac
and
Mario Moisés
Alvarez
*ab
f
Sergio O.
Martínez-Chapa,
c
Grissel Trujillo-de
Santiago
*ac
and
Mario Moisés
Alvarez
*ab
Abstract
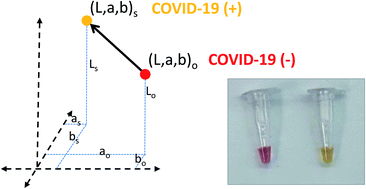
We demonstrate a loop-mediated isothermal amplification (LAMP) method to detect and amplify SARS-CoV-2 genetic sequences using a set of in-house designed initiators that target regions encoding the N protein. We were able to detect and amplify SARS-CoV-2 nucleic acids in the range of 62 to 2 × 105 DNA copies by this straightforward method. Using synthetic SARS-CoV-2 samples and RNA extracts from patients, we demonstrate that colorimetric LAMP is a quantitative method comparable in diagnostic performance to RT-qPCR (i.e., sensitivity of 92.85% and specificity of 81.25% in a set of 44 RNA extracts from patients analyzed in a hospital setting).
- This article is part of the themed collections: ENQA - 20th Brazilian Meeting on Analytical Chemistry, Coronavirus articles - free to access collection and Coronavirus collection – Analytical
 Please wait while we load your content...
Please wait while we load your content...

